Monitoring Exercise-Induced Muscle Fatigue and Adaptations: Making Sense of Popular or Emerging Indices and Biomarkers

 ,
,  , and
, and 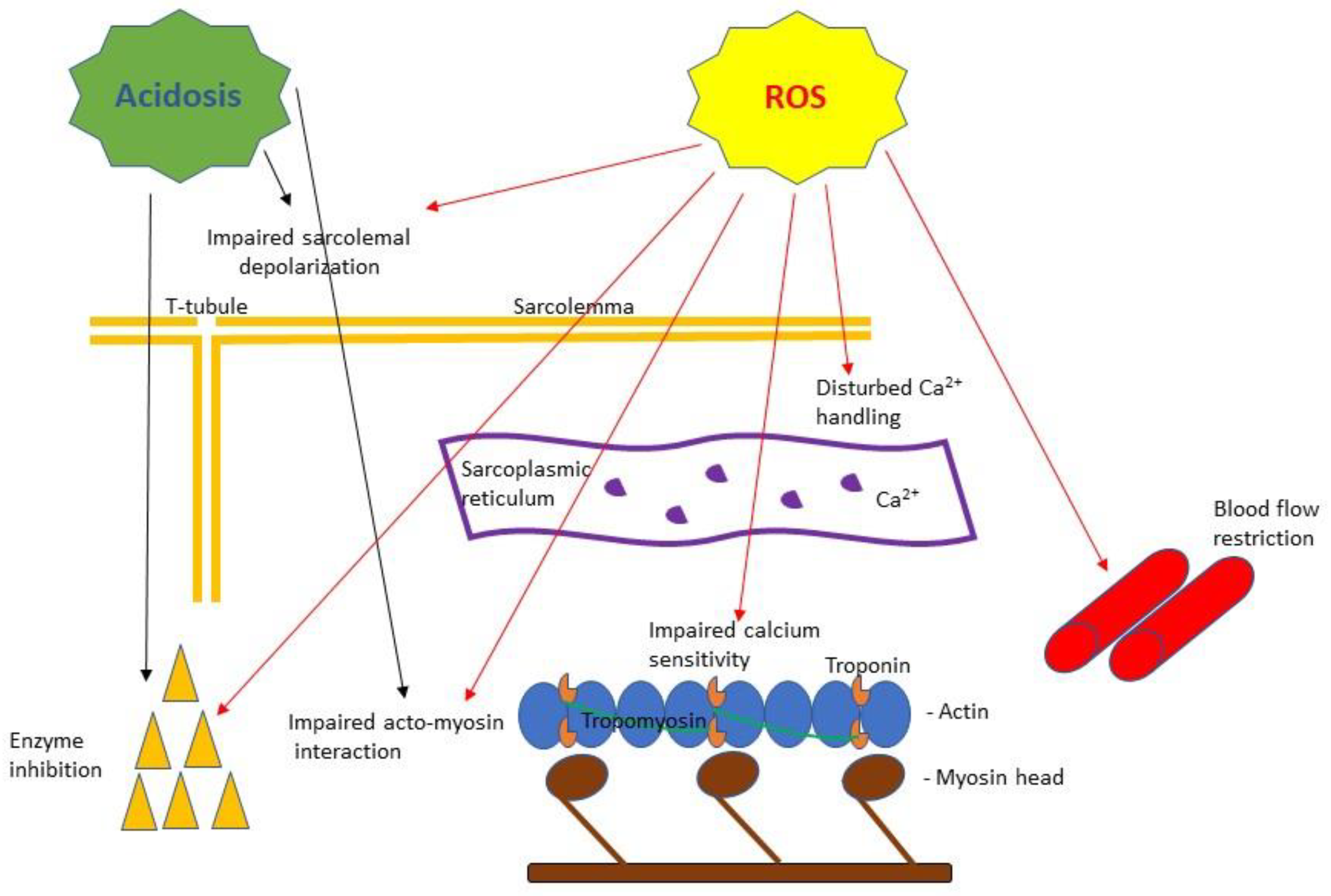{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Skeletal Muscle Fatigue
4. Lactate as a Fatigue Agent and as a Signaling Molecule
5. Exercise-Induced Oxidative Stress and Inflammation
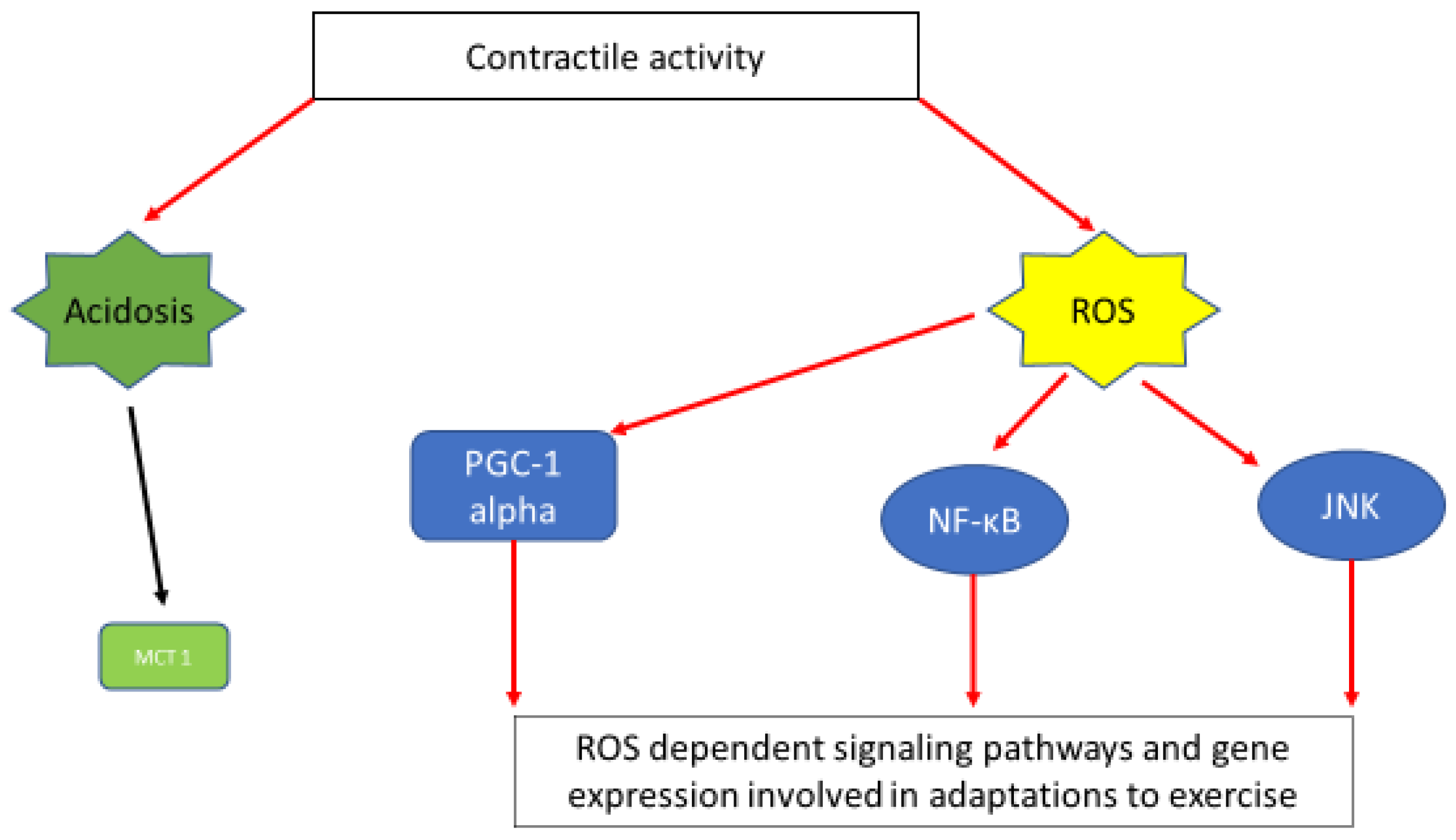
6. Reactive Oxygen Species (ROS) as Fatigue Agents and Signaling Molecules
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Frontera, W.R.; Ochala, J. Skeletal muscle: A brief review of structure and function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Westerblad, H.; Bruton, J.D.; Katz, A. Skeletal muscle: Energy metabolism, fiber types, fatigue and adaptability. Exp. Cell Res. 2010, 316, 3093–3099. [Google Scholar] [CrossRef] [PubMed]
- Bangsbo, J. Performance in sports—With specific emphasis on the effect of intensified training. Scand. J. Med. Sci. Sports 2015, 25, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Mahajan, P.B. Can genotype determine the sports phenotype? A paradigm shift in sports medicine. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Nader, G.A. Concurrent strength and endurance training: From molecules to man. Med. Sci. Sports Exerc. 2006, 38, 1965–1970. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [PubMed]
- Cooke, R. Modulation of the actomyosin interaction during fatigue of skeletal muscle. Muscle Nerve 2007, 36, 756–777. [Google Scholar] [CrossRef] [PubMed]
- Chase, P.B.; Kushmerick, M.J. Effects of pH on contraction of rabbit fast and slow skeletal muscle fibers. Biophys. J. 1988, 53, 935–946. [Google Scholar] [CrossRef] [Green Version]
- Nemes, R.; Koltai, E.; Taylor, A.W.; Suzuki, K. Reactive Oxygen and Nitrogen Species Regulate Key Metabolic, Anabolic, and Catabolic Pathways in Skeletal Muscle. Antioxidants 2018, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Karatzaferi, C.; Franks-Skiba, K.; Cooke, R. Inhibition of shortening velocity of skinned skeletal muscle fibers in conditions that mimic fatigue. Am. J. Physiol. Regul. Integrat. Comp. Physiol. 2008, 294, R948–R955. [Google Scholar] [CrossRef] [PubMed]
- Mitrou, G.I.; Poulianiti, K.P.; Koutedakis, Y.; Jamurtas, A.Z.; Maridaki, M.D.; Stefanidis, I.; Sakkas, G.K.; Karatzaferi, C. Functional responses of uremic single skeletal muscle fibers to redox imbalances. Hippokratia 2017, 21, 3. [Google Scholar] [PubMed]
- Allen, D.G.; Clugston, E.; Petersen, Y.; Roder, I.V.; Chapman, B.; Rudolf, R. Interactions between intracellular calcium and phosphate in intact mouse muscle during fatigue. J. Appl. Physiol. 2011, 111, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzger, J.M.; Moss, R.L. Greater hydrogen ion-induced depression of tension and velocity in skinned single fibres of rat fast than slow muscles. J. Physiol. 1987, 393, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Finsterer, J.; Mahjoub, S.Z. Fatigue in healthy and diseased individuals. Am. J. Hosp. Palliat. Care 2014, 31, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, R.T.; Atkinson, G.; Drust, B.; Gregson, W. Monitoring Fatigue Status in Elite Team-Sport Athletes: Implications for Practice. Int. J. Sports Physiol. Perform. 2017, 12, S227–S234. [Google Scholar] [CrossRef] [PubMed]
- Finsterer, J.; Drory, V.E. Wet, volatile, and dry biomarkers of exercise-induced muscle fatigue. BMC Musculoskelet. Disord. 2016, 17, 40. [Google Scholar] [CrossRef] [PubMed]
- Kent-Braun, J.A.; Fitts, R.H.; Christie, A. Skeletal muscle fatigue. Compr. Physiol. 2012, 2, 997–1044. [Google Scholar] [CrossRef] [PubMed]
- Finsterer, J. Biomarkers of peripheral muscle fatigue during exercise. BMC Musculoskelet. Disord. 2012, 13, 218. [Google Scholar] [CrossRef] [PubMed]
- Enoka, R.M.; Duchateau, J. Muscle fatigue: What, why and how it influences muscle function. J. Physiol. 2008, 586, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Noakes, T.D. Fatigue is a Brain-Derived Emotion that Regulates the Exercise Behavior to Ensure the Protection of Whole Body Homeostasis. Front. Physiol. 2012, 3, 82. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, G.K.; Karatzaferi, C. Hemodialysis fatigue: Just “simple” fatigue or a syndrome on its own right? Front. Physiol. 2012, 3, 306. [Google Scholar] [CrossRef] [PubMed]
- Debold, E.P.; Walcott, S.; Woodward, M.; Turner, M.A. Direct observation of phosphate inhibiting the force-generating capacity of a miniensemble of Myosin molecules. Biophys. J. 2013, 105, 2374–2384. [Google Scholar] [CrossRef] [PubMed]
- Karatzaferi, C.; Adamek, N.; Geeves, M.A. Modulators of actin-myosin dissociation: Basis for muscle type functional differences during fatigue. Am. J. Physiol. Cell Physiol. 2017, 313, C644–C654. [Google Scholar] [CrossRef] [PubMed]
- Debold, E.P. Potential molecular mechanisms underlying muscle fatigue mediated by reactive oxygen and nitrogen species. Front. Physiol. 2015, 6, 239. [Google Scholar] [CrossRef] [PubMed]
- Pickar, J.G.; Atrakchi, A.; Gray, S.D.; Carlsen, R.C. Apparent upregulation of Na+,K+ pump sites in SHR skeletal muscle with reduced transport capacity. Clin. Exp. Hypertens. Part A Theory Pract. 1991, 13, 645–652. [Google Scholar] [CrossRef]
- Nogueira, L.; Shiah, A.A.; Gandra, P.G.; Hogan, M.C. Ca2+-pumping impairment during repetitive fatiguing contractions in single myofibers: Role of cross-bridge cycling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R118–R125. [Google Scholar] [CrossRef] [PubMed]
- Leppik, J.A.; Aughey, R.J.; Medved, I.; Fairweather, I.; Carey, M.F.; McKenna, M.J. Prolonged exercise to fatigue in humans impairs skeletal muscle Na+-K+-ATPase activity, sarcoplasmic reticulum Ca2+ release, and Ca2+ uptake. J. Appl. Physiol. 2004, 97, 1414–1423. [Google Scholar] [CrossRef] [PubMed]
- Kannus, P. Isokinetic evaluation of muscular performance: Implications for muscle testing and rehabilitation. Int. J. Sports Med. 1994, 15, S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Vandewalle, H.; Peres, G.; Monod, H. Standard anaerobic exercise tests. Sports Med. 1987, 4, 268–289. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Villanueva, A.; Hamer, P.; Bishop, D. Fatigue in repeated-sprint exercise is related to muscle power factors and reduced neuromuscular activity. Eur. J. Appl. Physiol. 2008, 103, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karatzaferi, C.; Giakas, G.; Ball, D. Fatigue profile: A numerical method to examine fatigue in cycle ergometry. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 508–510. [Google Scholar] [CrossRef] [PubMed]
- Terzis, G.; Spengos, K.; Manta, P.; Sarris, N.; Georgiadis, G. Fiber type composition and capillary density in relation to submaximal number of repetitions in resistance exercise. J. Strength Cond. Res. Natl. Strength Cond. Assoc. 2008, 22, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, J.L.; Brechue, W.F.; Smith, A.E.; Kemmler, W.; Lauber, D.; Koch, A.J. Impact of testing strategy on expression of upper-body work capacity and one-repetition maximum prediction after resistance training in college-aged men and women. J. Strength Cond. Res. Natl. Strength Cond. Assoc. 2011, 25, 2796–2807. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.L.; Richard, R.; Binsse, V.M.; Koralsztein, J.P.; Haouzi, P. The VO2 slow component for severe exercise depends on type of exercise and is not correlated with time to fatigue. J. Appl. Physiol. 1998, 85, 2118–2124. [Google Scholar] [CrossRef] [PubMed]
- Slawinski, J.S.; Billat, V.L. Changes in internal mechanical cost during overground running to exhaustion. Med. Sci. Sports Exerc. 2005, 37, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Tsintzas, O.K.; Williams, C.; Boobis, L.; Greenhaff, P. Carbohydrate ingestion and single muscle fiber glycogen metabolism during prolonged running in men. J. Appl. Physiol. 1996, 81, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Black, M.I.; Jones, A.M.; Blackwell, J.R.; Bailey, S.J.; Wylie, L.J.; McDonagh, S.T.; Thompson, C.; Kelly, J.; Sumners, P.; Mileva, K.N.; et al. Muscle metabolic and neuromuscular determinants of fatigue during cycling in different exercise intensity domains. J. Appl. Physiol. 2017, 122, 446–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhatalo, A.; Black, M.I.; DiMenna, F.J.; Blackwell, J.R.; Schmidt, J.F.; Thompson, C.; Wylie, L.J.; Mohr, M.; Bangsbo, J.; Krustrup, P.; et al. The mechanistic bases of the power-time relationship: Muscle metabolic responses and relationships to muscle fibre type. J. Physiol. 2016, 594, 4407–4423. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, R.A.; Fluck, D.; Bonne, T.C.; Burgi, S.; Christensen, P.M.; Toigo, M.; Lundby, C. Improvements in exercise performance with high-intensity interval training coincide with an increase in skeletal muscle mitochondrial content and function. J. Appl. Physiol. 2013, 115, 785–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorentzon, R.; Johansson, C.; Sjostrom, M.; Fagerlund, M.; Fugl-Meyer, A.R. Fatigue during dynamic muscle contractions in male sprinters and marathon runners: Relationships between performance, electromyographic activity, muscle cross-sectional area and morphology. Acta Physiol. Scand. 1988, 132, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Bosquet, L.; Gouadec, K.; Berryman, N.; Duclos, C.; Gremeaux, V.; Croisier, J.L. Physiological Interpretation of the Slope during an Isokinetic Fatigue Test. Int. J. Sports Med. 2015, 36, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Bosquet, L.; Gouadec, K.; Berryman, N.; Duclos, C.; Gremeaux, V.; Croisier, J.L. The Total Work Measured During a High Intensity Isokinetic Fatigue Test Is Associated With Anaerobic Work Capacity. J. Sports Sci. Med. 2016, 15, 126–130. [Google Scholar] [PubMed]
- Bogdanis, G.C. Effects of physical activity and inactivity on muscle fatigue. Front. Physiol. 2012, 3, 142. [Google Scholar] [CrossRef] [PubMed]
- Bogdanis, G.C.; Nevill, M.E.; Boobis, L.H.; Lakomy, H.K. Contribution of phosphocreatine and aerobic metabolism to energy supply during repeated sprint exercise. J. Appl. Physiol. 1996, 80, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Bogdanis, G.C.; Nevill, M.E.; Boobis, L.H.; Lakomy, H.K.; Nevill, A.M. Recovery of power output and muscle metabolites following 30 s of maximal sprint cycling in man. J. Physiol. 1995, 482, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Karatzaferi, C.; de Haan, A.; van Mechelen, W.; Sargeant, A.J. Metabolism changes in single human fibres during brief maximal exercise. Exp. Physiol. 2001, 86, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Karatzaferi, C.; de Haan, A.; Ferguson, R.A.; van Mechelen, W.; Sargeant, A.J. Phosphocreatine and ATP content in human single muscle fibres before and after maximum dynamic exercise. Pflugers Arch. Eur. J. Physiol. 2001, 442, 467–474. [Google Scholar] [CrossRef]
- Wyon, M.A.; Koutedakis, Y. Muscular fatigue: Considerations for dance. J. Dance Med. Sci. 2013, 17, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.S.; Wary, C.; Wray, D.W.; Hoff, J.; Rossiter, H.B.; Layec, G.; Carlier, P.G. MRS Evidence of Adequate O(2) Supply in Human Skeletal Muscle at the Onset of Exercise. Med. Sci. Sports Exerc. 2015, 47, 2299–2307. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.M.; Nordsborg, N.B.; Nybo, L.; Mortensen, S.P.; Sander, M.; Secher, N.H.; Bangsbo, J. Thigh oxygen uptake at the onset of intense exercise is not affected by a reduction in oxygen delivery caused by hypoxia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R843–R849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, H.B. Arterial desaturation during exercise in man: Implication for O2 uptake and work capacity. Scand. J. Med. Sci. Sports 2003, 13, 339–358. [Google Scholar] [CrossRef] [PubMed]
- Cairns, S.P. Lactic acid and exercise performance: Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Messonnier, L.; Kristensen, M.; Juel, C.; Denis, C. Importance of pH regulation and lactate/H+ transport capacity for work production during supramaximal exercise in humans. J. Appl. Physiol. 2007, 102, 1936–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohmoto, O.; Spitzer, K.W.; Movsesian, M.A.; Barry, W.H. Effects of intracellular acidosis on [Ca2+]i transients, transsarcolemmal Ca2+ fluxes, and contraction in ventricular myocytes. Circ. Res. 1990, 66, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Woodward, M.; Debold, E.P. Acidosis and Phosphate Directly Reduce Myosin’s Force-Generating Capacity Through Distinct Molecular Mechanisms. Front. Physiol. 2018, 9, 862. [Google Scholar] [CrossRef] [PubMed]
- Parolin, M.L.; Chesley, A.; Matsos, M.P.; Spriet, L.L.; Jones, N.L.; Heigenhauser, G.J. Regulation of skeletal muscle glycogen phosphorylase and PDH during maximal intermittent exercise. Am. J. Physiol. 1999, 277, E890–E900. [Google Scholar] [CrossRef] [PubMed]
- Bogdanis, G.C.; Nevill, M.E.; Lakomy, H.K. Effects of previous dynamic arm exercise on power output during repeated maximal sprint cycling. J. Sports Sci. 1994, 12, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Posterino, G.S.; Fryer, M.W. Effects of high myoplasmic L-lactate concentration on E-C coupling in mammalian skeletal muscle. J. Appl. Physiol. 2000, 89, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Posterino, G.S.; Dutka, T.L.; Lamb, G.D. L(+)-lactate does not affect twitch and tetanic responses in mechanically skinned mammalian muscle fibres. Pflugers Arch. Eur. J. Physiol. 2001, 442, 197–203. [Google Scholar] [CrossRef]
- Kristensen, M.; Albertsen, J.; Rentsch, M.; Juel, C. Lactate and force production in skeletal muscle. J. Physiol. 2005, 562, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGinley, C.; Bishop, D.J. Rest interval duration does not influence adaptations in acid/base transport proteins following 10 wk of sprint-interval training in active women. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R702–R717. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Ye, X.; Xie, M.; Ye, J. Induction of triglyceride accumulation and mitochondrial maintenance in muscle cells by lactate. Sci. Rep. 2016, 6, 33732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikooie, R.; Samaneh, S. Exercise-induced lactate accumulation regulates intramuscular triglyceride metabolism via transforming growth factor-beta1 mediated pathways. Mol. Cell. Endocrinol. 2016, 419, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Juel, C. Regulation of pH in human skeletal muscle: Adaptations to physical activity. Acta Physiol. 2008, 193, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Driss, T.; Vandewalle, H. The measurement of maximal (anaerobic) power output on a cycle ergometer: A critical review. BioMed Res. Int. 2013, 2013, 589361. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.M.; Rajasekaran, S.; Thomsen, T.W.; Peterson, A.R. Lactate: Friend or Foe. PM&R 2016, 8, S8–S15. [Google Scholar] [CrossRef]
- Gupta, S.; Goswami, A.; Mukhopadhyay, S. Heart rate and blood lactate in 400 m flat and 400 m hurdle running: A comparative study. Indian J. Physiol. Pharmacol. 1999, 43, 361–366. [Google Scholar] [PubMed]
- Sporis, G.; Jukic, I.; Ostojic, S.M.; Milanovic, D. Fitness profiling in soccer: Physical and physiologic characteristics of elite players. J. Strength Cond. Res. Natl. Strength Cond. Assoc. 2009, 23, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.C.; D’Ottavio, S.; Vera, J.G.; Castagna, C. Aerobic fitness in futsal players of different competitive level. J. Strength Cond. Res. Natl. Strength Cond. Assoc. 2009, 23, 2163–2166. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Fuchs, P.; Fusco, A.; Fuchs, P.; Bell, W.J.; Duvillard, S.P. Physical Performance in Elite Male and Female Team Handball Players. Int. J. Sports Physiol. Perform. 2018. [Google Scholar] [CrossRef] [PubMed]
- Beneke, R.; Leithauser, R.M.; Ochentel, O. Blood lactate diagnostics in exercise testing and training. Int. J. Sports Physiol. Perform. 2011, 6, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Neil Spurway, A.M.J. Lactate Testing. In SPOST AND EXERCISE PHYSIOLOGY TESTING GUIDELINES The British Association of Sport and Exercise Sciences Guide; Routledge: Oxon, UK, 2006; Volume II, pp. 112–119. [Google Scholar]
- Jamnick, N.A.; Botella, J.; Pyne, D.B. Manipulating graded exercise test variables affects the validity of the lactate threshold and V−O2 peak. PLoS ONE 2018, 13, e0199794. [Google Scholar] [CrossRef] [PubMed]
- Lima-Silva, A.E.; Pires, F.O.; Bertuzzi, R.; Silva-Cavalcante, M.D.; Oliveira, R.S.; Kiss, M.A.; Bishop, D. Effects of a low- or a high-carbohydrate diet on performance, energy system contribution, and metabolic responses during supramaximal exercise. Appl. Physiol. Nutr. Metab. 2013, 38, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Maughan, R.J.; Greenhaff, P.L.; Leiper, J.B.; Ball, D.; Lambert, C.P.; Gleeson, M. Diet composition and the performance of high-intensity exercise. J. Sports Sci. 1997, 15, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Nelson, W.B.; Hudson, M.B. Exercise-induced oxidative stress in humans: Cause and consequences. Free Radic. Biol. Med. 2011, 51, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Nikolaidis, M.G.; Kyparos, A.; Spanou, C.; Paschalis, V.; Theodorou, A.A.; Vrabas, I.S. Redox biology of exercise: An integrative and comparative consideration of some overlooked issues. J. Exp. Biol. 2012, 215, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Close, G.L.; Ashton, T.; McArdle, A.; Maclaren, D.P. The emerging role of free radicals in delayed onset muscle soreness and contraction-induced muscle injury. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 142, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Ji, L.L.; Kavazis, A.N.; Jackson, M.J. Reactive oxygen species: Impact on skeletal muscle. Compr. Physiol. 2011, 1, 941–969. [Google Scholar] [CrossRef] [PubMed]
- Close, G.L.; Ashton, T.; Cable, T.; Doran, D.; MacLaren, D.P. Eccentric exercise, isokinetic muscle torque and delayed onset muscle soreness: The role of reactive oxygen species. Eur. J. Appl. Physiol. 2004, 91, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Gokbel, H.; Oz, M.; Okudan, N.; Belviranli, M.; Esen, H. Effects of exercise preconditioning on intestinal ischemia-reperfusion injury. Bratisl. Lek. Listy 2014, 115, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Fadel, P.J.; Farias Iii, M.; Gallagher, K.M.; Wang, Z.; Thomas, G.D. Oxidative stress and enhanced sympathetic vasoconstriction in contracting muscles of nitrate-tolerant rats and humans. J. Physiol. 2012, 590, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.B. Reactive Oxygen Species as Agents of Fatigue. Med. Sci. Sports Exerc. 2016, 48, 2239–2246. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M.J.; Medved, I.; Goodman, C.A.; Brown, M.J.; Bjorksten, A.R.; Murphy, K.T.; Petersen, A.C.; Sostaric, S.; Gong, X. N-acetylcysteine attenuates the decline in muscle Na+,K+-pump activity and delays fatigue during prolonged exercise in humans. J. Physiol. 2006, 576, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Bakalova, R.A.; Rangelova, D.S.; Stoyanovsky, D.A.; Koynova, G.M.; Wolinsky, I. Oxidative stress leads to inhibition of calcium transport by sarcoplasmic reticulum in skeletal muscle. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1989, 190, 365–368. [Google Scholar] [CrossRef]
- Andrade, F.H.; Reid, M.B.; Westerblad, H. Contractile response of skeletal muscle to low peroxide concentrations: Myofibrillar calcium sensitivity as a likely target for redox-modulation. FASEB J. 2001, 15, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Moopanar, T.R.; Allen, D.G. Reactive oxygen species reduce myofibrillar Ca2+ sensitivity in fatiguing mouse skeletal muscle at 37 degrees C. J. Physiol. 2005, 564, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.C.; Moen, R.J.; Smith, E.A.; Titus, M.A.; Thomas, D.D. Structural and functional impact of site-directed methionine oxidation in myosin. Biochemistry 2011, 50, 10318–10327. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Naito, Y.; Yoshikawa, T. Potential role of oxidative protein modification in energy metabolism in exercise. Sub-Cell. Biochem. 2014, 77, 175–187. [Google Scholar] [CrossRef]
- Jendzjowsky, N.G.; Delorey, D.S. Acute superoxide scavenging reduces sympathetic vasoconstrictor responsiveness in short-term exercise-trained rats. J. Appl. Physiol. 2013, 114, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Coombes, J.S.; Powers, S.K.; Rowell, B.; Hamilton, K.L.; Dodd, S.L.; Shanely, R.A.; Sen, C.K.; Packer, L. Effects of vitamin E and alpha-lipoic acid on skeletal muscle contractile properties. J. Appl. Physiol. 2001, 90, 1424–1430. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.J.; Bruton, J.D.; Lanner, J.T.; Westerblad, H. Antioxidant treatments do not improve force recovery after fatiguing stimulation of mouse skeletal muscle fibres. J. Physiol. 2015, 593, 457–472. [Google Scholar] [CrossRef] [PubMed]
- McClung, J.M.; Deruisseau, K.C.; Whidden, M.A.; Van Remmen, H.; Richardson, A.; Song, W.; Vrabas, I.S.; Powers, S.K. Overexpression of antioxidant enzymes in diaphragm muscle does not alter contraction-induced fatigue or recovery. Exp. Physiol. 2010, 95, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cabrera, M.C.; Borras, C.; Pallardo, F.V.; Sastre, J.; Ji, L.L.; Vina, J. Decreasing xanthine oxidase-mediated oxidative stress prevents useful cellular adaptations to exercise in rats. J. Physiol. 2005, 567, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, A.C.; McKenna, M.J.; Medved, I.; Murphy, K.T.; Brown, M.J.; Della Gatta, P.; Cameron-Smith, D. Infusion with the antioxidant N-acetylcysteine attenuates early adaptive responses to exercise in human skeletal muscle. Acta Physiol. 2012, 204, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; O’Moore, K.M.; Dickman, J.R.; Ji, L.L. Exercise activation of muscle peroxisome proliferator-activated receptor-gamma coactivator-1alpha signaling is redox sensitive. Free Radic. Biol. Med. 2009, 47, 1394–1400. [Google Scholar] [CrossRef] [PubMed]
- Margaritelis, N.V.; Theodorou, A.A.; Paschalis, V.; Veskoukis, A.S.; Dipla, K.; Zafeiridis, A.; Panayiotou, G.; Vrabas, I.S.; Kyparos, A.; Nikolaidis, M.G. Adaptations to endurance training depend on exercise-induced oxidative stress: Exploiting redox interindividual variability. Acta Physiol. 2018, 222. [Google Scholar] [CrossRef] [PubMed]
- Mohr, M.; Draganidis, D.; Chatzinikolaou, A.; Barbero-Alvarez, J.C.; Castagna, C.; Douroudos, I.; Avloniti, A.; Margeli, A.; Papassotiriou, I.; Flouris, A.D.; et al. Muscle damage, inflammatory, immune and performance responses to three football games in 1 week in competitive male players. Eur. J. Appl. Physiol. 2016, 116, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Cobley, J.N.; Close, G.L.; Bailey, D.M.; Davison, G.W. Exercise redox biochemistry: Conceptual, methodological and technical recommendations. Redox Biol. 2017, 12, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Tanskanen, M.; Atalay, M.; Uusitalo, A. Altered oxidative stress in overtrained athletes. J. Sports Sci. 2010, 28, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jammes, Y.; Steinberg, J.G.; Delliaux, S. Chronic fatigue syndrome: Acute infection and history of physical activity affect resting levels and response to exercise of plasma oxidant/antioxidant status and heat shock proteins. J. Intern. Med. 2012, 272, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Knight, J.S.M.; Webster, S.; Purdue, J. Managing fatigue and early infection in football: A novel approach. In Proceedings of the Football Medicine Strategies: Return to Play London, London, UK, 9–10 April 2016; pp. 227–229. [Google Scholar]
- Becatti, M.; Mannucci, A.; Barygina, V.; Mascherini, G.; Emmi, G.; Silvestri, E.; Wright, D.; Taddei, N.; Galanti, G.; Fiorillo, C. Redox status alterations during the competitive season in elite soccer players: Focus on peripheral leukocyte-derived ROS. Intern. Emerg. Med. 2017, 12, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Carrera-Quintanar, L.; Funes, L.; Vicente-Salar, N.; Blasco-Lafarga, C.; Pons, A.; Micol, V.; Roche, E. Effect of polyphenol supplements on redox status of blood cells: A randomized controlled exercise training trial. Eur. J. Nutr. 2015, 54, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Total antioxidant capacity: Appraisal of a concept. J. Nutr. 2007, 137, 1493–1495. [Google Scholar] [CrossRef] [PubMed]
- Vezzoli, A.; Pugliese, L.; Marzorati, M.; Serpiello, F.R.; La Torre, A.; Porcelli, S. Time-course changes of oxidative stress response to high-intensity discontinuous training versus moderate-intensity continuous training in masters runners. PLoS ONE 2014, 9, e87506. [Google Scholar] [CrossRef] [PubMed]
- Spanidis, Y.; Stagos, D.; Orfanou, M.; Goutzourelas, N.; Bar-Or, D.; Spandidos, D.; Kouretas, D. Variations in Oxidative Stress Levels in 3 Days Follow-up in Ultramarathon Mountain Race Athletes. J. Strength Cond. Res. Natl. Strength Cond. Assoc. 2017, 31, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Jamurtas, A.Z.; Fatouros, I.G.; Deli, C.K.; Georgakouli, K.; Poulios, A.; Draganidis, D.; Papanikolaou, K.; Tsimeas, P.; Chatzinikolaou, A.; Avloniti, A.; et al. The Effects of Acute Low-Volume HIIT and Aerobic Exercise on Leukocyte Count and Redox Status. J. Sports Sci. Med. 2018, 17, 501–508. [Google Scholar] [PubMed]
- Zalavras, A.; Fatouros, I.G.; Deli, C.K.; Draganidis, D.; Theodorou, A.A.; Soulas, D.; Koutsioras, Y.; Koutedakis, Y.; Jamurtas, A.Z. Age-related responses in circulating markers of redox status in healthy adolescents and adults during the course of a training macrocycle. Oxid. Med. Cell. Longev. 2015, 2015, 283921. [Google Scholar] [CrossRef] [PubMed]
- Youssef, H.; Groussard, C.; Machefer, G.; Minella, O.; Couillard, A.; Knight, J.; Gratas-Delamarche, A. Comparison of total antioxidant capacity of salivary, capillary and venous samplings: Interest of the salivary total antioxidant capacity on triathletes during training season. J. Sports Med. Phys. Fit. 2008, 48, 522–529. [Google Scholar]
- Nikolaidis, M.G.; Kyparos, A.; Dipla, K.; Zafeiridis, A.; Sambanis, M.; Grivas, G.V.; Paschalis, V.; Theodorou, A.A.; Papadopoulos, S.; Spanou, C.; et al. Exercise as a model to study redox homeostasis in blood: The effect of protocol and sampling point. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2012, 17, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Goldfarb, A.H.; Rescino, M.H.; Hegde, S.; Patrick, S.; Apperson, K. Eccentric exercise effect on blood oxidative-stress markers and delayed onset of muscle soreness. Med. Sci. Sports Exerc. 2002, 34, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Kwak, Y.S. Impact of aerobic and anaerobic exercise training on oxidative stress and antioxidant defense in athletes. J. Exerc. Rehabil. 2016, 12, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Veskoukis, A.S.; Nikolaidis, M.G.; Kyparos, A.; Kouretas, D. Blood reflects tissue oxidative stress depending on biomarker and tissue studied. Free Radic. Biol. Med. 2009, 47, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
- Nystrom, T. Role of oxidative carbonylation in protein quality control and senescence. EMBO J. 2005, 24, 1311–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokota, T.; Kinugawa, S.; Yamato, M.; Hirabayashi, K.; Suga, T.; Takada, S.; Harada, K.; Morita, N.; Oyama-Manabe, N.; Kikuchi, Y.; et al. Systemic oxidative stress is associated with lower aerobic capacity and impaired skeletal muscle energy metabolism in patients with metabolic syndrome. Diabetes Care 2013, 36, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.M.; Bagatini, M.D.; Roth, M.A.; Martins, C.C.; Rezer, J.F.; Mello, F.F.; Lopes, L.F.; Morsch, V.M.; Schetinger, M.R. Acute effects of resistance exercise and intermittent intense aerobic exercise on blood cell count and oxidative stress in trained middle-aged women. Braz. J. Med. Biol. Res. 2012, 45, 1172–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubrilo, D.; Djordjevic, D.; Zivkovic, V.; Djuric, D.; Blagojevic, D.; Spasic, M.; Jakovljevic, V. Oxidative stress and nitrite dynamics under maximal load in elite athletes: Relation to sport type. Mol. Cell. Biochem. 2011, 355, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Peart, D.J.; Kirk, R.J.; Hillman, A.R.; Madden, L.A.; Siegler, J.C.; Vince, R.V. The physiological stress response to high-intensity sprint exercise following the ingestion of sodium bicarbonate. Eur. J. Appl. Physiol. 2013, 113, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Margaritelis, N.V.; Cobley, J.N.; Paschalis, V.; Veskoukis, A.S.; Theodorou, A.A.; Kyparos, A.; Nikolaidis, M.G. Principles for integrating reactive species into in vivo biological processes: Examples from exercise physiology. Cell. Signal. 2016, 28, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Nikolaidis, M.G.; Jamurtas, A.Z.; Paschalis, V.; Fatouros, I.G.; Koutedakis, Y.; Kouretas, D. The effect of muscle-damaging exercise on blood and skeletal muscle oxidative stress: Magnitude and time-course considerations. Sports Med. 2008, 38, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Paschalis, V.; Nikolaidis, M.G.; Fatouros, I.G.; Giakas, G.; Koutedakis, Y.; Karatzaferi, C.; Kouretas, D.; Jamurtas, A.Z. Uniform and prolonged changes in blood oxidative stress after muscle-damaging exercise. In Vivo 2007, 21, 877–883. [Google Scholar] [PubMed]
- Rodriguez, D.A.; Kalko, S.; Puig-Vilanova, E.; Perez-Olabarria, M.; Falciani, F.; Gea, J.; Cascante, M.; Barreiro, E.; Roca, J. Muscle and blood redox status after exercise training in severe COPD patients. Free Radic. Biol. Med. 2012, 52, 88–94. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Theofilidis, G.; Bogdanis, G.C.; Koutedakis, Y.; Karatzaferi, C. Monitoring Exercise-Induced Muscle Fatigue and Adaptations: Making Sense of Popular or Emerging Indices and Biomarkers. Sports 2018, 6, 153. https://0-doi-org.brum.beds.ac.uk/10.3390/sports6040153
Theofilidis G, Bogdanis GC, Koutedakis Y, Karatzaferi C. Monitoring Exercise-Induced Muscle Fatigue and Adaptations: Making Sense of Popular or Emerging Indices and Biomarkers. Sports. 2018; 6(4):153. https://0-doi-org.brum.beds.ac.uk/10.3390/sports6040153
Chicago/Turabian StyleTheofilidis, George, Gregory C. Bogdanis, Yiannis Koutedakis, and Christina Karatzaferi. 2018. "Monitoring Exercise-Induced Muscle Fatigue and Adaptations: Making Sense of Popular or Emerging Indices and Biomarkers" Sports 6, no. 4: 153. https://0-doi-org.brum.beds.ac.uk/10.3390/sports6040153