Impact of Different Exercise Modalities on the Human Gut Microbiome

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subject Population and General Design
2.2. Procedures
2.2.1. EXMP-CRE—Cardiorespiratory Fitness
2.2.2. EXMP-RTE—Resistance Training
2.3. EXMP-CRE Measures
2.3.1. Maximal Aerobic Capacity
2.3.2. Ventilatory Threshold
2.3.3. Anthropometric Measurements
2.4. EXMP-RTE Measures
2.4.1. Submaximal Muscular Strength
2.4.2. Anthropometric Measurements
2.5. Statistical Analysis of Fitness and Anthropometric Measures
2.6. DNA Extraction and Microbiome Sequencing
2.6.1. EXMP-CRE
2.6.2. EXMP-RTE
2.7. Microbiome Bioinformatics
2.8. Statistical Analysis of Microbiome Measures
3. Results
3.1. Exercise Interventions
3.1.1. EXMP-CRE—Cardiorespiratory Fitness
3.1.2. EXMP-RTE–Resistance Training
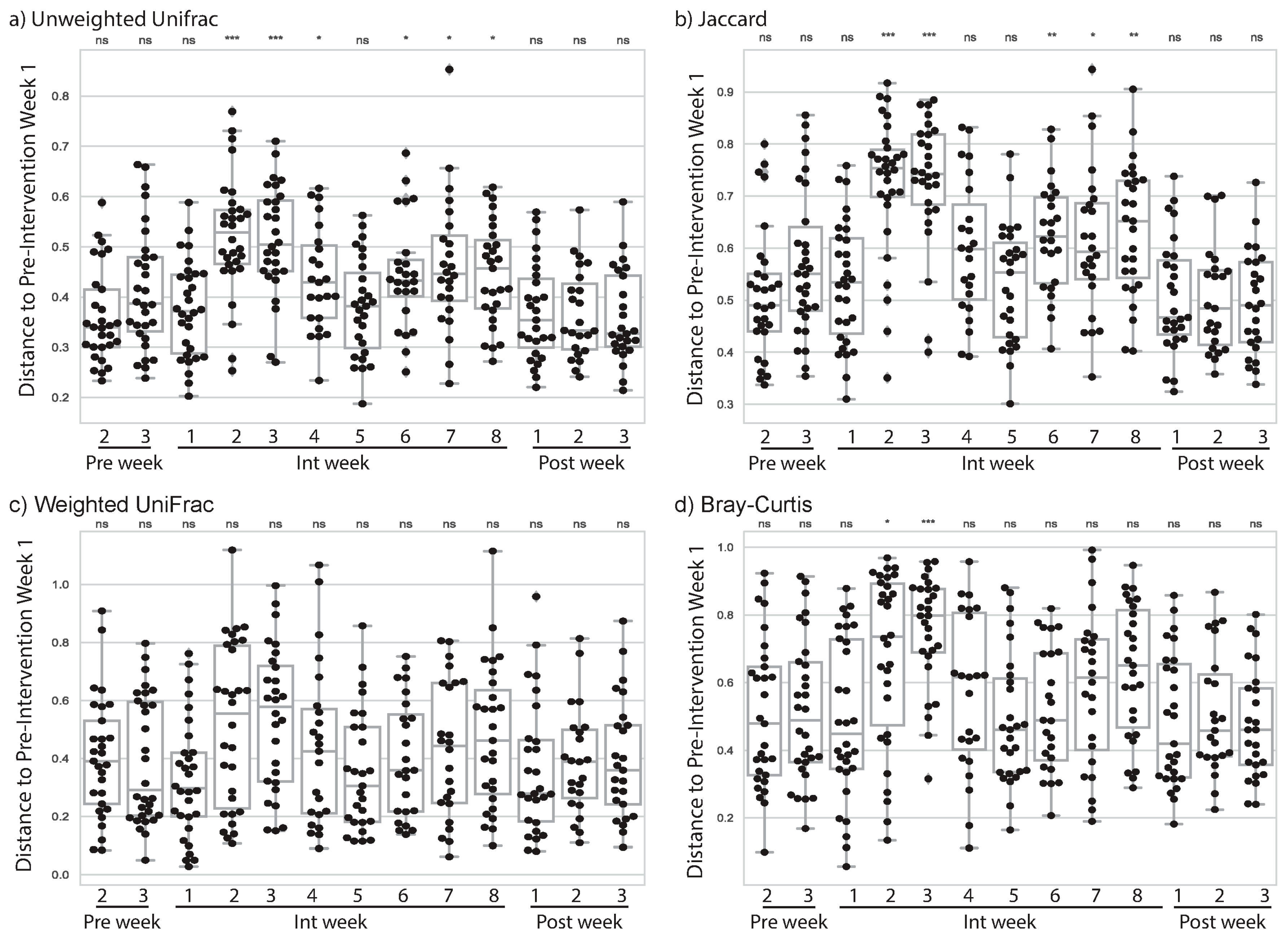
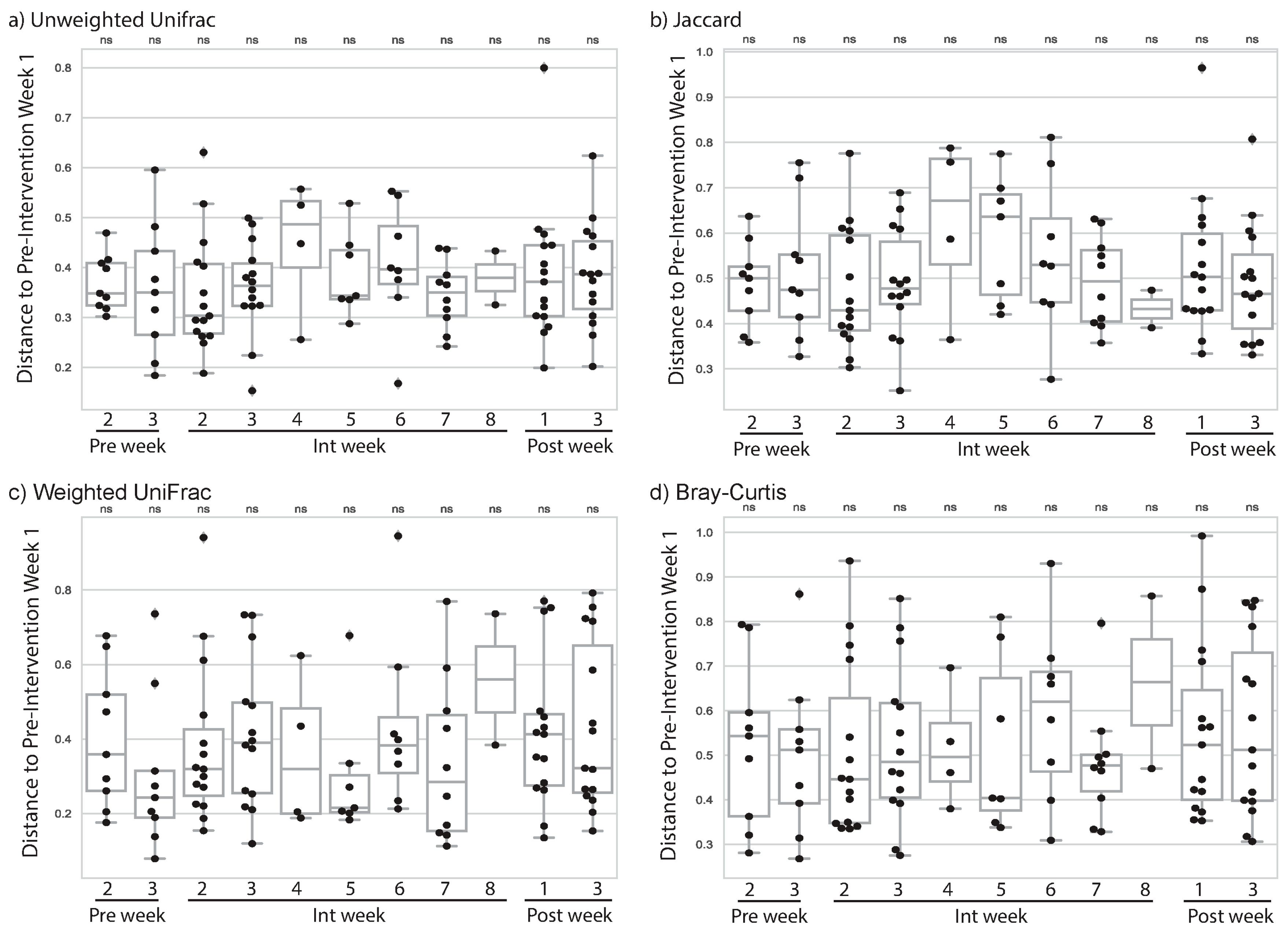
3.2. Cardiorespiratory Exercise Was Associated with Changes to Subjects’ Gut Microbiome
3.3. Resistance Training Was Not Associated with Changes to Subjects’ Gut Microbiome
3.4. Starting Microbiome State May Predict Magnitude of Change in CRE
3.5. Starting Microbiome State Predicts Exercise Gains during Resistance Training
3.6. Magnitude of Change in Microbiome Composition Is Not Correlated with Magnitude of Exercise Gains
4. Discussion
4.1. Change in Microbiome Associated with Cardiorespiratory Exercise
4.2. Cardiorespiratory Fitness Adaptations
4.3. Resistance Training Associated Change in Microbiome
4.4. Resistance Training Fitness Adaptations
4.5. Initial Microbiome Status and Subsequent Exercise Adaptations
4.6. Next Steps in Understanding the Role of Exercise in Shaping the Microbiome
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [Green Version]
- Łaniewski, P.; Ilhan, Z.E.; Herbst-Kralovetz, M.M. The microbiome and gynaecological cancer development, prevention and therapy. Nat. Rev. Urol. 2020, 17, 232–250. [Google Scholar] [CrossRef]
- Younis, N.; Zarif, R.; Mahfouz, R. Inflammatory bowel disease: Between genetics and microbiota. Mol. Biol. Rep. 2020, 47, 3053–3063. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The influence of the gut microbiome on cancer, immunity, and cancer immunotherapy. Cancer Cell 2018, 33, 570–588. [Google Scholar] [CrossRef] [Green Version]
- Pabst, O. Correlation, consequence, and functionality in microbiome-immune interplay. Immunol. Rev. 2017, 279, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Saha, S.; Khanna, S. Therapies to modulate gut microbiota: Past, present and future. World J. Gastroenterol. 2020, 26, 777–788. [Google Scholar] [CrossRef]
- Garber, C.E.; Blissmer, B.; Deschenes, M.R.; Franklin, B.A.; Lamonte, M.J.; Lee, I.M.; Nieman, D.C.; Swain, D.P. Quantity and quality of exercise for developing and maintaining cardiorespiratory, musculoskeletal, and neuromotor fitness in apparently healthy adults: Guidance for prescribing exercise. Med. Sci. Sports Exerc. 2011, 43, 1334–1359. [Google Scholar] [CrossRef]
- Mailing, L.J.; Allen, J.M.; Buford, T.W.; Fields, C.J.; Woods, J.A. Exercise and the gut microbiome: A review of the evidence, potential mechanisms, and implications for human health. Exerc. Sport Sci. Rev. 2019, 47, 75–85. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Davy, B.M.; Hulver, M.W.; Neilson, A.P.; Bennett, B.J.; Davy, K.P. Does exercise alter gut microbial composition? A systematic review. Med. Sci. Sports Exerc. 2019, 51, 160–167. [Google Scholar] [CrossRef]
- Pasini, E.; Corsetti, G.; Assanelli, D.; Testa, C.; Romano, C.; Dioguardi, F.S.; Aquilani, R. Effects of chronic exercise on gut microbiota and intestinal barrier in human with type 2 diabetes. Minerva Med. 2019, 110, 3–11. [Google Scholar] [CrossRef]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holscher, H.D.; Woods, J.A. Exercise alters gut microbiota composition and function in lean and obese humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef]
- Taniguchi, H.; Tanisawa, K.; Sun, X.; Kubo, T.; Hoshino, Y.; Hosokawa, M.; Takeyama, H.; Higuchi, M. Effects of short-term endurance exercise on gut microbiota in elderly men. Physiol. Rep. 2018, 6, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Munukka, E.; Ahtiainen, J.P.; Puigbó, P.; Jalkanen, S.; Pahkala, K.; Keskitalo, A.; Kujala, U.M.; Pietilä, S.; Hollmén, M.; Elo, L.; et al. Six-week endurance exercise alters gut metagenome that is not reflected in systemic metabolism in over-weight women. Front. Microbiol. 2018, 9, 2323. [Google Scholar] [CrossRef] [PubMed]
- Cronin, O.; Barton, W.; Skuse, P.; Penney, N.C.; Garcia-Perez, I.; Murphy, E.F.; Woods, T.; Nugent, H.; Fanning, A.; Melgar, S.; et al. A prospective metagenomic and metabolomic analysis of the impact of exercise and/or whey protein supplementation on the gut microbiome of sedentary adults. mSystems 2018, 3, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Y.; Ni, Y.; Cheung, C.K.; Lam, K.S.; Wang, Y.; Xia, Z.; Ye, D.; Guo, J.; Tse, M.A.; et al. Clinical and translational report gut microbiome fermentation determines the efficacy of exercise for diabetes prevention. Cell Metab. 2020, 31, 77–91.e5. [Google Scholar] [CrossRef]
- Álvarez, C.; Ramírez-Campillo, R.; Ramírez-Vélez, R.; Izquierdo, M. Prevalence of non-responders for glucose control markers after 10 weeks of high-intensity interval training in adult women with higher and lower insulin resistance. Front. Physiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Motiani, K.K.; Collado, M.C.; Eskelinen, J.J.; Virtanen, K.A.; Löyttyniemi, E.; Salminen, S.; Nuutila, P.; Kalliokoski, K.K.; Hannukainen, J.C. Exercise training modulates gut microbiota profile and improves endotoxemia. Med. Sci. Sports Exerc. 2020, 52, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Riebe, D.; Ehrman, J.K.; Liguori, G.; Magal, M. (Eds.) ACSM’s Guidelines for Exercise Testing and Prescription, 10th ed.; Wolters Kluwer: Philadelphia, PA, USA, 2018. [Google Scholar]
- Haff, G.G.; Triplett, N.T. (Eds.) Essentials of Strength Training and Conditioning, 4th ed.; Human Kinetics: Champaign, IL, USA, 2016. [Google Scholar]
- Bruce, R.A.; Kusumi, F.; Hosmer, D. Maximal oxygen intake and nomographic assessment of functional aerobic impairment in cardiovascular disease. Am. Heart J. 1973, 85, 546–562. [Google Scholar] [CrossRef]
- Balady, G.J.; Arena, R.; Sietsema, K.; Myers, J.; Coke, L.; Fletcher, G.F.; Forman, D.; Franklin, B.; Guazzi, M.; Gulati, M.; et al. Clinician’s guide to cardiopulmonary exercise testing in adults: A scientific statement from the American heart association. Circulation 2010, 122, 191–225. [Google Scholar] [CrossRef] [Green Version]
- LeSuer, D.A.; McCormick, J.H.; Mayhew, J.L.; Wasserstein, R.L.; Arnold, M.D. The accuracy of prediction equations for estimating 1-RM performance in the bench press, squat, and deadlift. J. Strength Cond. Res. 1997, 11, 211–213. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A communal catalogue reveals earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Janssen, S.; Mcdonald, D.; Gonzalez, A.; Navas-molina, J.A.; Jiang, L.; Xu, Z. Phylogenetic placement of exact amplicon sequences. mSystems 2018, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Estaki, M.; Pither, J.; Baumeister, P.; Little, J.P.; Gill, S.K.; Ghosh, S.; Ahmadi-Vand, Z.; Marsden, K.R.; Gibson, D.L. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 2016, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gibson, A.L.; Wagner, D.R.; Heyward, V.H. Advanced Fitness Assessment and Exercise Prescription, 8th ed.; Human Kinetics: Champaign, IL, USA, 2018. [Google Scholar]
- Hoffman, J. Norms for Fitness, Performance, and Health; Human Kinetics: Champaign, IL, USA, 2006. [Google Scholar]
- Poyet, M.; Groussin, M.; Gibbons, S.M.; Avila-Pacheco, J.; Jiang, X.; Kearney, S.M.; Perrotta, A.R.; Berdy, B.; Zhao, S.; Lieberman, T.D.; et al. A library of human gut bacterial isolates paired with longitudinal multiomics data enables mechanistic microbiome research. Nat. Med. 2019, 25, 1442–1452. [Google Scholar] [CrossRef]
- Bermon, S.; Petriz, B.; Kajeniene, A.; Prestes, J.; Castell, L.; Franco, O.L. The microbiota: An exercise immunology perspective. Exerc. Immunol. Rev. 2015, 21, 70–79. [Google Scholar]
- Heuberger, J.A.A.C.; Gal, P.; Stuurman, F.E.; De Muinck Keizer, W.A.S.; Miranda, Y.M.; Cohen, A.F. Repeatability and predictive value of lactate threshold concepts in endurance sports. PLoS ONE 2018, 13, e0206846. [Google Scholar] [CrossRef]
- Radovanović, S.; Kocić, S.; Gajović, G.; Radević, S.; Milosavljević, M.; Nićiforović, J. The impact of body weight on aerobic capacity. Med. Glas 2014, 11, 204–209. [Google Scholar]
- Pribis, P.; Burtnack, C.A.; Mckenzie, S.O.; Thayer, J. Trends in body fat, body mass index and physical fitness among male and female college students. Nutrients 2010, 2, 1075–1085. [Google Scholar] [CrossRef] [Green Version]
- Ratamess, N. ACSM’s Foundations of Strength Training and Conditioning; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012. [Google Scholar]
- Kraemer, W.J.; Adams, K.; Cafarelli, E.; Dudley, G.A.; Dooly, C.; Feigenbaum, M.S.; Fleck, S.J.; Franklin, B.; Fry, A.C.; Hoffman, J.R.; et al. Progression models in resistance training for healthy adults. Med. Sci. Sports Exerc. 2002, 34, 364–380. [Google Scholar]
- Chen, Y.M.; Wei, L.; Chiu, Y.S.; Hsu, Y.J.; Tsai, T.Y.; Wang, M.F.; Huang, C.C. Lactobacillus plantarum TWK10 supplementation improves exercise performance and increases muscle mass in mice. Nutrients 2016, 8, 205. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe. 2018, 23, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut Microbiota 2018, 67, 625–633. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Pre-Intervention Phase (Study Weeks 1–3; Pre Weeks 1–3) | Intervention Phase (Study Weeks 4–11; Int Weeks 1–8) | Post-Intervention Phase (Study Weeks 12–14; Post Weeks 1–3) |
|---|---|---|
|
|
|
| EXMP-CRE | ||||||
| Total Sample (N = 28) | Female Subjects (n = 21) | Male Subjects (n = 7) | ||||
| Pre | Post | Pre | Post | Pre | Post | |
| Age (year) | 20.54 (1.93) | 20.71 (1.88) | 20.00 (2.16) | |||
| Weight (kg) | 67.83 (10.70) | 68.14 (10.59) | 66.22 (10.84) | 66.57 (10.96) | 72.63 (9.35) | 72.86 (8.34) |
| BMI (kg·m−2) | 24.41 (4.20) | 24.55 (4.41) | 24.54 (4.58) | 24.72 (4.90) | 24.04 (3.02) | 24.06 (2.66) |
| FFM (kg) | 48.71 (7.66) | 48.85 (7.65) | 45.28 (4.11) | 45.43 (4.23) | 59.01 (6.52) | 59.11 (6.31) |
| %BF | 27.57 (9.13) | 27.68 (9.10) | 30.62 (7.77) | 30.71 (7.86) | 18.43 (6.63) | 18.57 (6.16) |
| EXMP-RTE | ||||||
| Total Sample (N = 28) | Female Subjects (n = 17) | Male Subjects (n = 11) | ||||
| Pre | Post | Pre | Post | Pre | Post | |
| Age (year) | 21.28 (3.85) | 20.41 (3.34) | 22.64 (4.34) | |||
| Weight (kg) | 67.72 (15.03) | 68.32 (14.67) | 61.58 (12.84) | 62.08 (12.02) | 77.20 (13.55) | 77.97 (13.44) |
| BMI (kg·m−2) | 23.77 (4.15) | 23.97 (3.93) | 23.24 (4.36) | 23.43 (4.11) | 24.59 (3.87) | 24.81 (3.67) |
| FFM (kg) | 49.58 (11.63) | 50.47 (12.32) * | 41.96 (5.15) | 42.55 (5.12) | 62.54 (6.89) | 63.95 (8.48) |
| %BF | 27.08 (8.10) | 26.54 (8.35) | 30.69 (6.49) | 30.46 (6.00) | 20.94 (6.92) | 19.86 (7.67) |
| EXMP-CRE | |||||||||
| Total Sample (N = 28) | Female Subjects (n = 21) | Male Subjects (n = 7) | |||||||
| Pre | Post | Δ | Pre | Post | Δ | Pre | Post | Δ | |
| VO2max (mL·kg−1 ·min−1) | 35.55 (6.48) | 35.57 (5.88) | 0.03 (2.91) | 33.59 (5.64) | 33.64 (5.08) | 0.06 (2.74) | 41.43 (5.38) | 41.36 (4.23) | −0.07 (3.62) |
| RER | 1.24 (0.12) | 1.31 (0.09) | 0.07 (0.13) ** | 1.21 (0.11) | 1.31 (0.09) | 0.09 (0.13) ** | 1.33 (0.11) | 1.32 (0.11) | −0.01 (0.08) |
| Treadmill Test Time (s) | 579.89 (114.78) | 631.11 (98.49) | 51.21 (76.17) ** | 550.48 (114.55) | 595.10 (75.10) | 44.62 (83.73) * | 668.14 (58.54) | 739.14 (81.84) | 71.00 (46.17) ** |
| EXMP-RTE | |||||||||
| Total Sample (N = 28) | Female Subjects (n = 17) | Male Subjects (n = 11) | |||||||
| Pre | Post | Δ | Pre | Post | Δ | Pre | Post | Δ | |
| 3RM Squat (kg) | 71.28 (30.48) | 96.47 (34.86) | 25.19 (14.11) *** | 56.83 (16.46) | 77.78 (16.51) | 20.95 (8.58) *** | 93.60 (34.22) | 125.36 (36.52) | 31.75 (18.48) *** |
| Pred. 1RM Squat (kg) | 75.54 (32.30) | 102.24 (36.95) | 26.70 (14.95) *** | 60.23 (17.45) | 82.43 (17.49) | 22.20 (9.09) *** | 99.20 (36.26) | 132.86 (38.70) | 33.64 (19.59) *** |
| 3RM Bench Press (kg) | 43.25 (22.20) | 50.95 (22.88) | 7.69 (4.11) *** | 28.55 (6.32) | 35.62 (6.20) | 7.07 (3.75) *** | 65.98 (18.19) | 74.64 (18.30) | 8.66 (4.63) *** |
| Pred. 1RM Bench Press (kg) | 45.84 (23.52) | 54.00 (24.24) | 8.16 (4.36) *** | 30.26 (6.69) | 37.75 (6.57) | 7.49 (3.98) *** | 69.92 (19.27) | 79.10 (19.39) | 9.18 (4.90) *** |
| 3RM Bent-Over Row (kg) | 56.78 (23.36) | 74.19 (25.78) | 17.41 (5.92) *** | 41.89 (7.61) | 57.10 (8.49) | 15.21 (4.73) *** | 79.79 (20.53) | 100.62 (20.34) | 20.82 (6.15) *** |
| Pred. 1RM Bent-Over Row (kg) | 60.18 (24.76) | 78.63 (27.32) | 18.46 (6.28) *** | 44.40 (8.07) | 60.52 (9.00) | 16.12 (5.01) *** | 84.56 (21.76) | 106.63 (21.56) | 22.07 (6.52) *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bycura, D.; Santos, A.C.; Shiffer, A.; Kyman, S.; Winfree, K.; Sutliffe, J.; Pearson, T.; Sonderegger, D.; Cope, E.; Caporaso, J.G. Impact of Different Exercise Modalities on the Human Gut Microbiome. Sports 2021, 9, 14. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9020014
Bycura D, Santos AC, Shiffer A, Kyman S, Winfree K, Sutliffe J, Pearson T, Sonderegger D, Cope E, Caporaso JG. Impact of Different Exercise Modalities on the Human Gut Microbiome. Sports. 2021; 9(2):14. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9020014
Chicago/Turabian StyleBycura, Dierdra, Anthony C. Santos, Arron Shiffer, Shari Kyman, Kyle Winfree, Jay Sutliffe, Talima Pearson, Derek Sonderegger, Emily Cope, and J. Gregory Caporaso. 2021. "Impact of Different Exercise Modalities on the Human Gut Microbiome" Sports 9, no. 2: 14. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9020014