
Weak Association between Vastus Lateralis Muscle Fiber Composition and Fascicle Length in Young Untrained Females


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
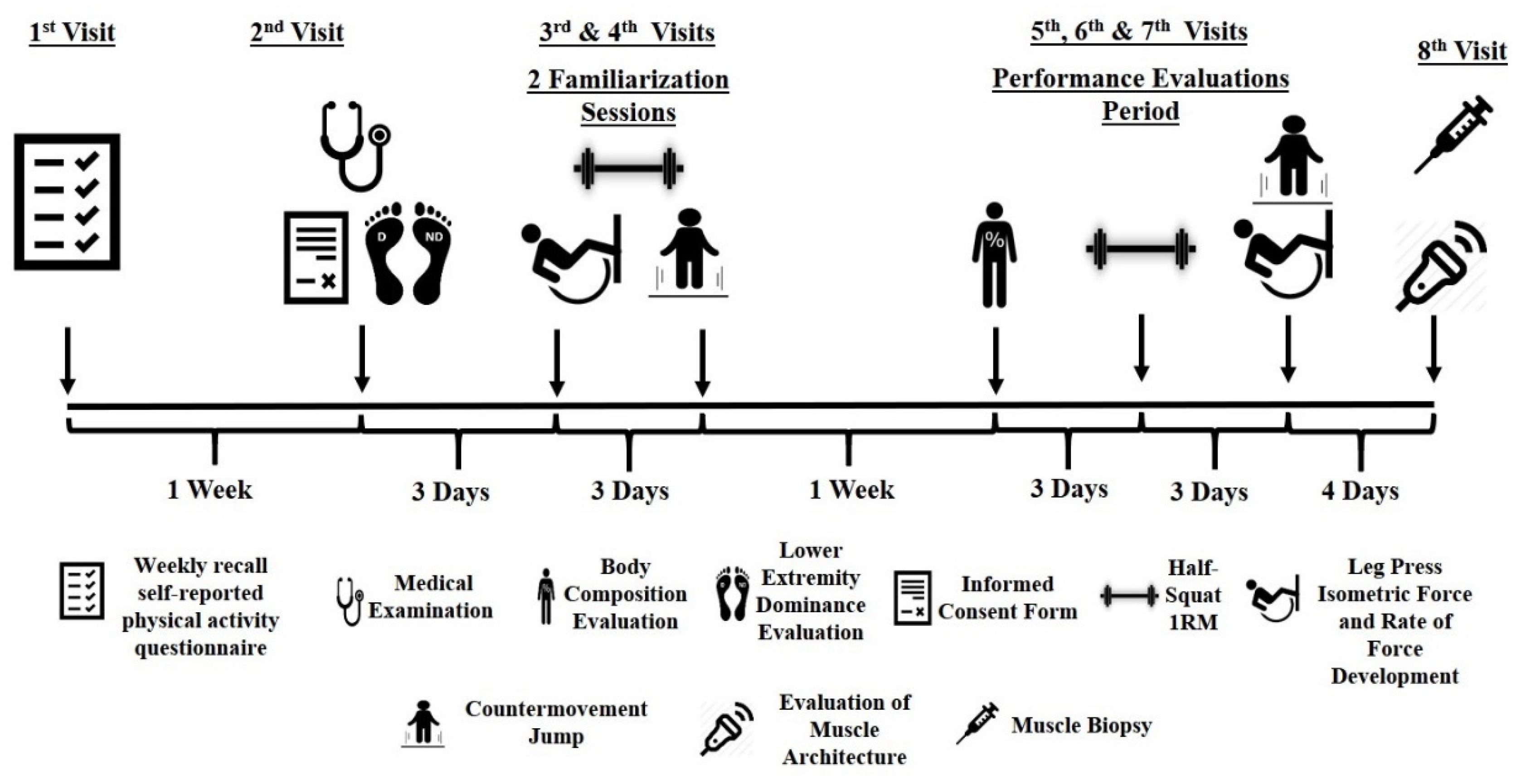
2.1. Experimental Approach
2.2. Participants
2.3. Procedures
2.3.1. Evaluation of Body Composition
2.3.2. Evaluation of Muscle Strength and Power
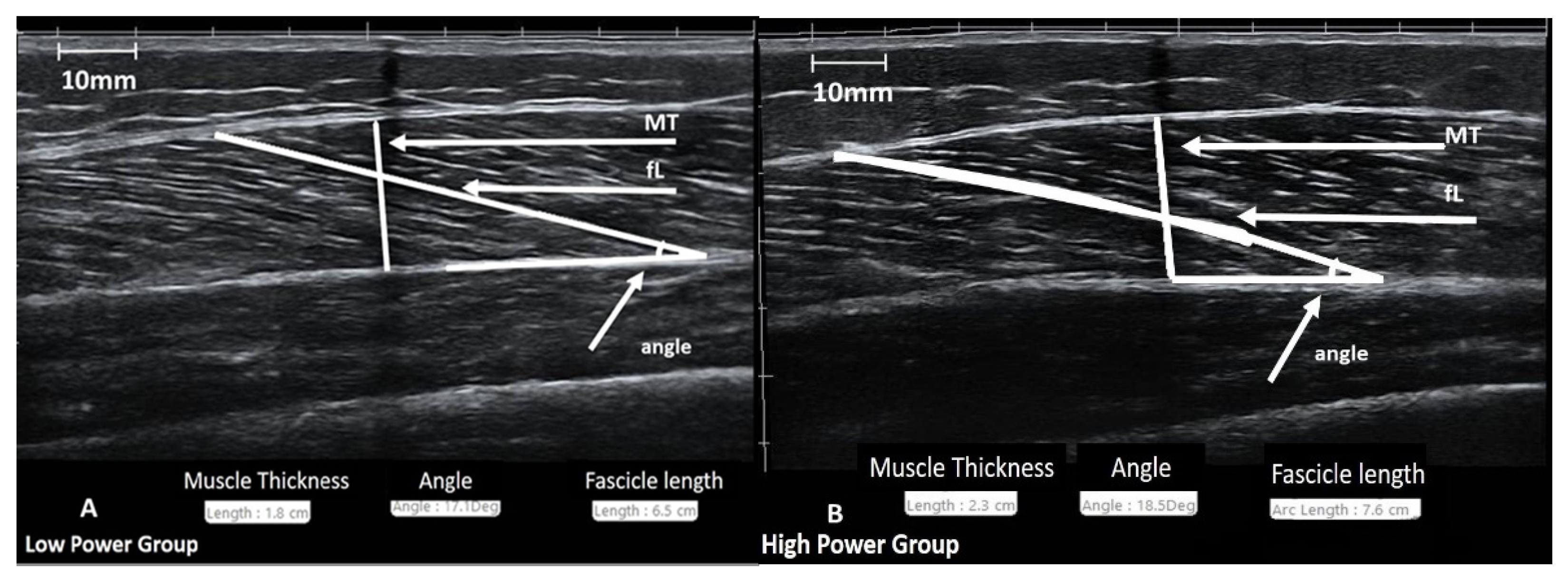
2.3.3. Evaluation of Muscle Architecture
2.3.4. Muscle Biopsies and Histochemistry
2.4. Statistical Analyses
3. Results
3.1. Comparison between Groups
3.2. Correlations between Biological and Performance Parameters
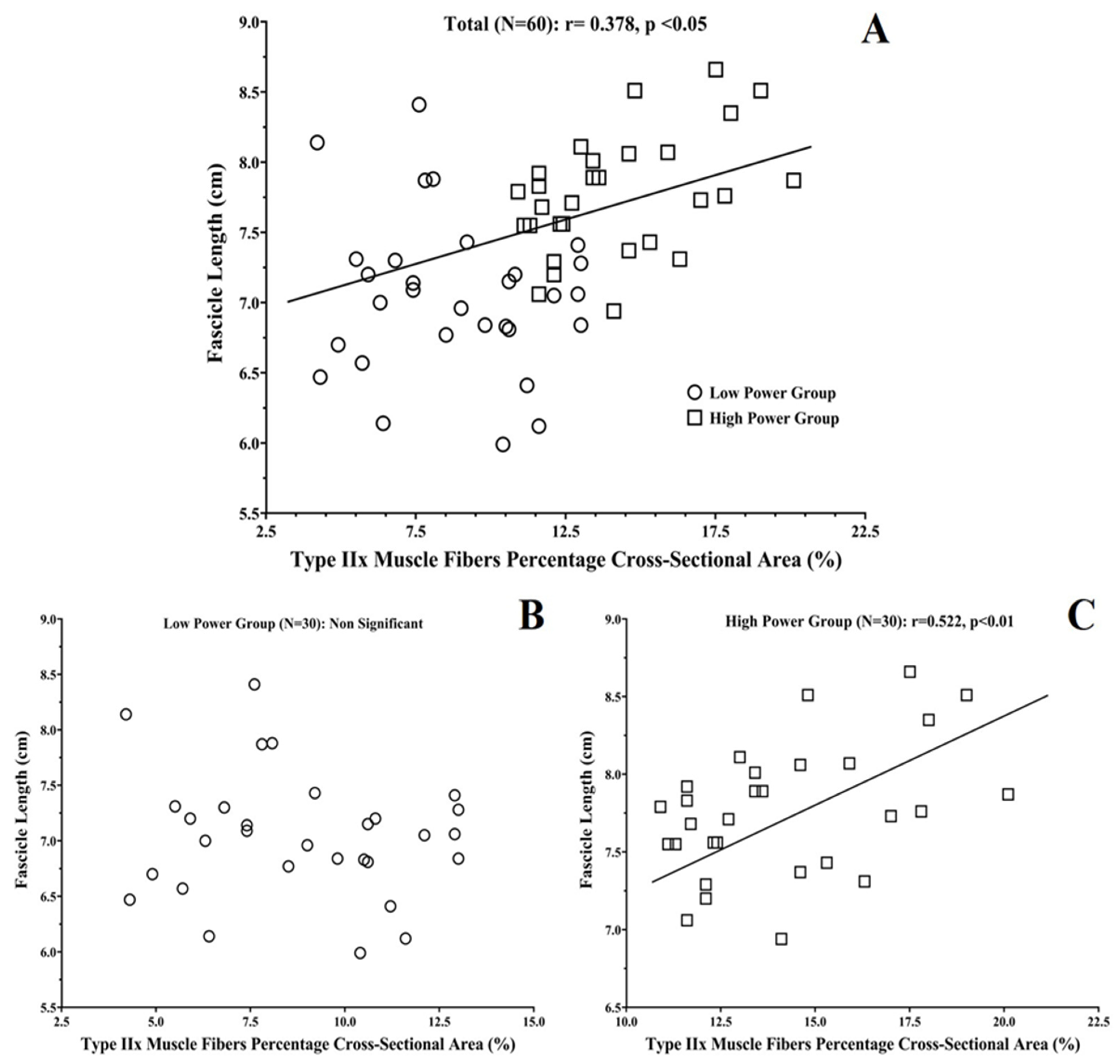
3.3. Correlations between Vastus Lateralis Architecture and Muscle Fiber Compostition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mero, A.; Luhtanen, P.; Viitasalo, J.T. Relationships between the maximal running velocity, muscle fiber characteristics, force production and force relaxation of sprinters. Scand. J. Sports Sci. 1981, 3, 16–22. [Google Scholar]
- Methenitis, S.; Karandreas, N.; Spengos, K.; Zaras, N.; Stasinaki, A.N.; Terzis, G. Muscle fiber conduction velocity, muscle fiber composition, and power performance. Med. Sci. Sports. Exerc. 2016, 48, 1761–1771. [Google Scholar] [CrossRef] [PubMed]
- Bottinelli, R.; Canepari, M.; Pellegrino, M.A.; Reggiani, C. Force-velocity properties of human skeletal muscle fibres: Myosin heavy chain isoform and temperature dependence. J. Physiol. 1996, 495 Pt 2, 573–586. [Google Scholar] [CrossRef]
- Stafilidis, S.; Arampatzis, A. Muscle–tendon unit mechanical and morphological properties and sprint performance. J Sports Sci. 2007, 25, 1035–1046. [Google Scholar] [CrossRef]
- Abe, T.; Loenneke, J.P.; Thiebaud, R.S. Morphological and functional relationships with ultrasound measured muscle thickness of the lower extremity: A brief review. Ultrasound 2015, 23, 166–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazevich, A.J.; Gill, N.D.; Bronks, R.; Newton, R.U. Training-specific muscle architecture adaptation after 5-wk training in athletes. Med. Sci. Sports Exerc. 2003, 35, 2013–2022. [Google Scholar] [CrossRef] [PubMed]
- Blazevich, A.; Sharp, C. Understanding muscle architectural adaptation: Macro-and micro-level research. Cells Tissues Organs. 2006, 181, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ema, R.; Akagi, R.; Wakahara, T.; Kawakami, Y. Training-induced changes in architecture of human skeletal muscles: Current evidence and unresolved issues. J Phys. Fit Sport. Med. 2016, 5, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Stasinaki, A.N.; Zaras, N.; Methenitis, S.; Tsitkanou, S.; Krase, A.; Kavvoura, A.; Terzis, G. Triceps Brachii Muscle Strength and Architectural Adaptations with Resistance Training Exercises at Short or Long Fascicle Length. J. Funct. Morphol. Kinesiol. 2018, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Stasinaki, A.N.; Zaras, Ν.; Methenitis, S.; Bogdanis, G.; Terzis, G. Rate of force development and muscle architecture after fast and slow velocity eccentric training. Sports 2019, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmins, R.G.; Shield, A.J.; Williams, M.D.; Lorenzen, C.; Opar, D.A. Architectural adaptations of muscle to training and injury: A narrative review outlining the contributions by fascicle length, pennation angle and muscle thickness. Br. J. Sports Med. 2016, 50, 1467–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaras, N.; Stasinaki, A.N.; Spiliopoulou, P.; Arnaoutis, G.; Hadjicharalambous, M.; Terzis, G. Rate of Force Development, Muscle Architecture, and Performance in Elite Weightlifters. Int. J. Sports Physiol. Perform. 2020, 16, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Zaras, N.; Stasinaki, A.; Spiliopoulou, P.; Hadjicharalambous, M.; Terzis, G. Lean Body Mass, Muscle Architecture, and Performance in Well-Trained Female Weightlifters. Sports 2020, 8, 67. [Google Scholar] [CrossRef]
- Zaras, N.; Stasinaki, A.N.; Methenitis, S.; Karampatsos, G.; Fatouros, I.; Hadjicharalambous, M.; Terzis, G. Track and field throwing performance prediction: Training intervention, muscle architecture adaptations and field tests explosiveness ability. J. Phys. Educ. Sport. 2019, 19, 436–443. [Google Scholar] [CrossRef]
- Narici, M.; Franchi, M.; Maganaris, C. Muscle structural assembly and functional consequences. J. Exp. Biol. 2016, 219 Pt 2, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Methenitis, S.; Zaras, N.; Spengos, K.; Stasinaki, A.N.; Karampatsos, G.; Georgiadis, G.; Terzis, G. Role of muscle morphology in jumping, sprinting, and throwing performance in participants with different power training duration experience. J. Strength. Cond. Res. 2016, 30, 807–817. [Google Scholar] [CrossRef]
- Zaras, N.D.; Stasinaki, A.N.; Methenitis, S.K.; Krase, A.A.; Karampatsos, G.P.; Georgiadis, G.V.; Spengos, K.M.; Terzis, G.D. Rate of force development, muscle architecture, and performance in young competitive track and field throwers. J. Strength. Cond. Res. 2016, 30, 81–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coratella, G.; Longo, S.; Rampichini, S.; Limonta, E.; Shokohyar, S.; Bisconti, V.A.; Ce, E.; Espocito, F. Quadriceps and gastrocnemii anatomical cross-sectional area and vastus lateralis fascicle length predict peak-power and time-to-peak-power. Res. Q. Exerc. Sport. 2020, 91, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Fukashiro, S.; Harada, Y.; Kawamoto, K. Relationship between sprint performance and muscle fascicle length in female sprinters. J. Physiol. Anthropol. Appl. Human. Sci. 2001, 20, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brechue, W.F.; Abe, T. The role of FFM accumulation and skeletal muscle architecture in powerlifting performance. Eur. J. Appl. Physiol. 2002, 86, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Lieber, R.; Fridén, J. Clinical significance of skeletal muscle architecture. Clin. Orthop. Relat. Res. 2001, 383, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Kumagai, K.; Brechue, W. Fascicle length of leg muscles is greater in sprinters than distance runners. Med. Sci. Sports Exerc. 2000, 32, 1125–1129. [Google Scholar] [CrossRef] [Green Version]
- Alegre, L.M.; Aznar, D.; Delgado, T.; Jimenez, F.; Aguado, X. Architectural characteristics of vastus lateralis muscle and jump performance in young men. J. Hum. Mov. Stud. 2005, 48, 109–123. [Google Scholar]
- Mangine, T.G.; Fukuda, H.D.; LaMonica, B.M.; Gonzalez, M.A.; Wells, J.D.; Townsend, R.J.; Jajtner, R.A.; Fragala, S.M.; Stout, R.J.; Hoffman, R.J. Influence of gender and muscle architecture asymmetry on jump and sprint performance. J. Sports. Sci. Med. 2014, 13, 904–911. [Google Scholar] [PubMed]
- Costill, D.L.; Daniels, J.; Evans, W.; Fink, W.; Krahenbuhl, G.; Saltin, B. Skeletal muscle enzymes and fiber composition in male and female track athletes. J. Appl. Physiol. 1976, 40, 149–154. [Google Scholar] [CrossRef]
- Methenitis, S.; Stasinaki, A.N.; Zaras, N.; Spengos, K.; Karandreas, N.; Terzis, G. Intramuscular fibre conduction velocity and muscle fascicle length in human vastus lateralis. Appl. Physiol. Nutr. Metab. 2019, 44, 133–138. [Google Scholar] [CrossRef]
- Burkholder, T.; Fingado, B.; Baron, S.; Lieber, R. Relationship between muscle fiber types and sizes and muscle architectural properties in the mouse hindlimb. J. Morphol. 1994, 221, 177–190. [Google Scholar] [CrossRef]
- Methenitis, S.; Mpampoulis, T.; Spiliopoulou, P.; Papadimas, G.; Papadopoulos, C.; Chalari, E.; Evangelidou, E.; Stasinaki, A.N.; Nomikos, T.; Terzis, G. Muscle fiber composition, jumping performance and rate of force development adaptations induced by different power training volumes in females. Appl. Physiol. Nutr. Metab. 2020, 45, 996–1006. [Google Scholar] [CrossRef]
- Sharifnezhad, A.; Marzilger, R.; Arampatzis, A. Effects of load magnitude, muscle length and velocity during eccentric chronic loading on the longitudinal growth of the vastus lateralis muscle. J. Exp. Biol. 2014, 217, 2726–2733. [Google Scholar] [CrossRef] [Green Version]
- Stasinaki, A.N.; Gloumis, G.; Spengos, K.; Blazevich, J.A.; Zaras, N.; Georgiadis, G.; Karampatsos, G.; Terzis, G. Muscle strength, power, and morphologic adaptations after 6 weeks of compound vs. complex training in healthy men. J. Strength. Cond. Res. 2015, 29, 2559–2569. [Google Scholar] [CrossRef] [PubMed]
- Kavvoura, A.; Zaras, N.; Stasinaki, A.N.; Arnaoutis, G.; Methenitis, S.; Terzis, G. The importance of lean body mass for the rate of force development in taekwondo athletes and track and field throwers. J. Funct. Morphol. Kinesiol. 2018, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Terzis, G.; Spengos, K.; Kavouras, S.; Manta, P.; Georgiadis, G. Muscle fibre type composition and body composition in hammer throwers. J. Sports Sci. Med. 2010, 9, 104–109. [Google Scholar] [PubMed]
- Terzis, G.; Spengos, K.; Methenitis, S.; Aagaard, P.; Karandreas, N.; Bogdanis, G. Early phase interference between low-intensity running and power training in moderately trained females. Eur. J. Appl. Physiol. 2016, 116, 1063–1073. [Google Scholar] [CrossRef]
- Zaras, N.; Stasinaki, A.N.; Spiliopoulou, P.; Mpampoulis, T.; Hadjicharalambous, M.; Terzis, G. Effect of inter-repetition rest vs. traditional strength training on lower body strength, rate of force development, and muscle architecture. Appl. Sci. 2021, 11, 45. [Google Scholar] [CrossRef]
- Maffiuletti, N.A.; Aagaard, P.; Blazevich, A.J.; Folland, J.; Tillin, N.; Duchateau, J. Rate of force development: Physiological and methodological considerations. Eur. J. Appl. Physiol. 2016, 116, 1091–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Rosell, D.; Pareja-Blanco, F.; Aagaard, P.; González-Badillo, J. Physiological and methodological aspects of rate of force development assessment in human skeletal muscle. Clin. Physiol. Funct. Imaging 2017, 38, 743–762. [Google Scholar] [CrossRef] [PubMed]
- Spiliopoulou, P.; Zaras, N.; Methenitis, S.; Papadimas, G.; Papadopoulos, C.; Bogdanis, G.; Terzis, G. Effect of concurrent power training and high-intensity interval cycling on muscle morphology and performance. J. Strength. Cond. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zacharia, E.; Spiliopoulou, P.; Methenitis, S.; Stasinaki, A.N.; Zaras, N.; Papadopoulos, C.; Papadimas, C.; Karampatsos, G.; Bogdanis, G.C.; Terzis, G. Changes in muscle power and muscle morphology with different volumes of fast eccentric half-squats. Sports 2019, 7, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsitkanou, S.; Spengos, K.; Stasinaki, A.N.; Zaras, N.; Bogdanis, G.; Papadimas, G.; Terzis, G. Effects of high-intensity interval cycling performed after resistance training on muscle strength and hypertrophy. Scand. J. Med. Sci. Sports 2017, 27, 1317–1327. [Google Scholar] [CrossRef]
- Ounjian, M.; Roy, R.R.; Eldred, E.; Garfinkel, A.; Payne, J.R.; Armstrong, A.; Toga, W.; Edgerton, V.R. Physiological and developmental implications of motor unit anatomy. J. Neurobiol. 1991, 22, 547–559. [Google Scholar] [CrossRef]
- Lexell, J.; Downham, D.; Sjöström, M. Morphological detection of neurogenic muscle disorders: How can statistical methods aid diagnosis? Acta Neuropathol. 1987, 75, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.L.; Andersen, J.L.; Zebis, M.K.; Aagaard, P. Early and late rate of force development: Differential adaptive responses to resistance training? Scand. J. Med. Sci. Sports. 2010, 20, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Fry, A.C.; Schilling, B.K.; Staron, R.S.; Hagerman, F.C.; Hikida, R.S.; Thrush, J.T. Muscle fiber characteristics and performance correlates of male Olympic-style weightlifters. J. Strength. Cond. Res. 2003, 17, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Hvid, L.; Aagaard, P.; Justensen, L.; Bayer, M.L.; Andersen, J.L.; Ørtenblad, N.; Kjaer, M.; Suetta, C. Effects of aging on muscle mechanical function and muscle fiber morphology during short-term immobilization and subsequent. J. Appl. Physiol. 2010, 109, 1628–1634. [Google Scholar] [CrossRef] [Green Version]
- Billeter, R.; Jostarndt-Fögen, K.; Günthör, W.; Hoppeler, H. Fiber type characteristics and myosin light chain expression in a world champion shot putter. Int. J. Sports Med. 2003, 24, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Zaras, N.; Spengos, K.; Methenitis, S.; Papadopoulos, C.; Karampatsos, G.; Georgiadis, G.; Stasinaki, A.; Manta, P.; Terzis, G. Effects of strength vs. ballistic-power training on throwing performance. J. Sports Sci. Med. 2013, 12, 130–137. [Google Scholar]
- Widrick, J.; Trappe, S.; Costill, D.; Fitts, R.H. Force-velocity and force-power properties of single muscle fibers from elite master runners and sedentary men. Am. J. Physiol. 1996, 271, 676–683. [Google Scholar] [CrossRef]
- Brocca, L.; Longa, E.; Cannavino, J.; Seynnes, O.; de Vito, G.; McPhee, J.; Narici, M.; Pellegrino, M.A.; Bottinelli, R. Human skeletal muscle fibre contractile properties and proteomic profile: Adaptations to 3 weeks of unilateral lower limb suspension and active recovery. J. Physiol. 2015, 593, 5361–5385. [Google Scholar] [CrossRef] [Green Version]
- Frontera, W.; Suh, D.; Krivickas, L.; Hughes, V.; Goldstein, R.; Roubenoff, R. Skeletal muscle fiber quality in older men and women. Am. J. Physiol. Cell Physiol. 2000, 279, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Bedrin, N.; Ades, P.; Palmer, P.; Toth, M. Molecular determinants of force production in human skeletal muscle fibers: Effects of myosin isoform expression and cross-sectional area. Am. J. Physiol. Cell. Physiol. 2015, 308, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottinelli, R.; Pellegrino, M.; Canepari, M.; Rossi, R.; Reggiani, C. Specific contributions of various muscle fibre types to human muscle performance: An in vitro study. J. Electromyogr. Kinesiol. 1999, 9, 87–95. [Google Scholar] [CrossRef]
- Canepari, M.; Pellegrino, M.; D’Antona, G.; Bottinelli, R. Skeletal muscle fibre diversity and the underlying mechanisms. Acta Physiol. 2010, 199, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Harridge, S.; Bottinelli, R.; Canepari, M.; Pellegrino, M.A.; Reggiani, C.; Esbjörnsson, M.; Saltin, B. Whole-muscle and single-fibre contractile properties and myosin heavy chain isoforms in humans. Pflüg. Arch. 1996, 432, 913–920. [Google Scholar] [CrossRef]
- Bodine, S.C.; Roy, R.R.; Meadows, D.A.; Zernicke, R.F.; Sacks, R.D.; Fournier, M.; Edgerton, V.R. Architectural, histochemical, and contractile characteristics of a unique biarticular muscle: The cat semitendinosus. J. Neurophysiol. 1982, 48, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieber, L.R.; Friden, J. Functional and clinical significance of skeletal muscle architecture. Muscle Nerve 2000, 23, 1647–1666. [Google Scholar] [CrossRef]
- Cormie, P.; McGuigan, M.R.; Newton, R.U. Developing maximal neuromuscular power: Part 1--biological basis of maximal power production. Sports Med. 2011, 41, 17–38. [Google Scholar] [CrossRef]
- Methenitis, S.; Spengos, K.; Zaras, N.; Stasinaki, A.N.; Papadimas, G.; Karampatsos, G.; Arnaoutis, G.; Terzis, G. Fiber type composition and rate of force development in endurance and resistance trained individuals. J. Strength. Cond. Res. 2019, 33, 2388–2397. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, A.D.; Negro, F.; Holobar, A.; Casolo, A.; Folland, J.; Felici, F.; Farina, D. You are as fast as your motor neurons: Speed of recruitment and maximal discharge of motor neurons determine the maximal rate of force development in humans. J. Physiol. 2019, 597, 2445–2456. [Google Scholar] [CrossRef] [Green Version]
- Dideriksen, J.; Del Vecchio, A.; Farina, D. Neural and muscular determinants of maximal rate of force development. J. Neurophysiol. 2020, 123, 149–157. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| All Participants (N = 60) | Comparison between the Groups | ||||
|---|---|---|---|---|---|
| High Power (N = 30) | Low Power (N = 30) | Partial η2/p Values | |||
| Lean Mass (kg) | 41.7 ± 4.2 | 43.4 ± 4.2 | 39.9 ± 3.5 | 0.173/0.009 * | |
| Legs Lean Mass (kg) | 14.5 ± 1.6 | 15.3 ± 1.4 | 13.6 ± 1.5 | 0.239/0.002 * | |
| Vastus Lateralis Architecture | |||||
| Muscle Thickness (cm) | 2.1 ± 0.3 | 2.2 ± 03 | 1.8 ± 0.2 | 0.157/0.038 * | |
| Pennation angle (°) | 17.7 ± 3.7 | 18.4 ± 3 | 17.1 ± 3.2 | 0.029/0.192 | |
| Fascicle length (cm) | 7.1 ± 0.7 | 7.3 ± 0.7 | 6.5 ± 0.8 | 0.181/0.001 * | |
| Vastus Lateralis Fiber Type Composition | |||||
| Type I Percentage (%) | 48.65 ± 6.4 | 46.1 ± 7.8 | 51.2 ± 4.6 | 0.126/0.009 * | |
| Type IIa Percentage (%) | 38.5 ± 6.7 | 37.6 ± 8.0 | 39.3 ± 5.2 | 0.016/0.337 | |
| Type IIx Percentage (%) | 12.9 ± 5.1 | 16.3 ± 5.8 | 9.5 ± 3.4 | 0.127/0.005 * | |
| Type I Cross-Sectional Area (μm2) | 3349 ± 444 | 3622 ± 471 | 3173 ± 425 | 0.123/0.010 * | |
| Type IIa Cross-Sectional Area (μm2) | 3411 ± 462 | 3803 ± 420 | 3219 ± 505 | 0.119/0.010 * | |
| Type IIx Cross-Sectional Area (μm2) | 2627 ± 431 | 2997 ± 231 | 2162 ± 229 | 0.124/0.009 * | |
| Type I Percentage Cross-Sectional Area (%) | 47.9 ± 7.5 | 43.6 ± 8.9 | 53.1 ± 5.9 | 0.128/0.009 * | |
| Type IIa Percentage Cross-Sectional Area (%) | 40.7 ± 7.2 | 42.6 ± 8.2 | 38.7 ± 6.0 | 0.092/0.043 * | |
| Type IIx Percentage Cross-Sectional Area (%) | 11.4 ± 4.4 | 14.2 ± 4.8 | 8.2 ± 3.9 | 0.136/0.002 * | |
| Counter Movement Jump | |||||
| Power (W) | 1948 ± 388 | 2236 ± 315 | 1640 ± 145 | 0.598/0.000 * | |
| Height (cm) | 25.6 ± 4.2 | 27.1 ± 4.8 | 24.1 ± 2.7 | 0.134/0.005 * | |
| Maximum Isometric Strength and Rate of Force Development | |||||
| Maximum Isometric Force (N) | 2035 ± 607 | 2215 ± 697 | 1849 ± 435 | 0.092/0.019 * | |
| Rate of Force Development (N·s−1) | 20 ms | 6799 ± 2644 | 8030 ± 2450 | 5526 ± 2227 | 0.227/<0.001 * |
| 80 ms | 8777 ± 3228 | 10347 ± 2800 | 7153 ± 2844 | 0.248/<0.001 * | |
| 100 ms | 8740 ± 3028 | 10192 ± 2593 | 7238 ± 2727 | 0.241/<0.001 * | |
| 150 ms | 8025 ± 2382 | 9070 ± 2145 | 6944 ± 2147 | 0.202/<0.001 * | |
| 200 ms | 7018 ± 1952 | 7779 ± 1730 | 6231 ± 1878 | 0.159/0.002 * | |
| 250 ms | 6080 ± 1660 | 6663 ± 1491 | 5478 ± 1632 | 0.129/0.005 * | |
| Maximum Strength | |||||
| Half Squat 1-RM Strength (kg) | 112.7 ± 19.7 | 117.6 ± 20.5 | 107.8 ± 17.7 | 0.082/0.043 * | |
| Legs Lean Body Mass | Vastus Lateralis Architecture | Vastus Lateralis Fiber Type Composition | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Percentage | Cross-Sectional Area | Percentage Cross-Sectional Area | |||||||||||
| Muscle Thickness | Pennation Angle | Fascicle Length | IIa | IIx | I | IIa | IIx | I | IIa | IIx | |||
| All Participants (N = 60) | |||||||||||||
| CMJP | 0.595 | 0.411 | 0.279 | 0.341 | |||||||||
| MIF | 0.389 | 0.330 | 0.355 | −0.403 | 0.410 | 0.384 | −0.499 | 0.434 | |||||
| RFD | 150 ms | 0.456 | 0.282 | ||||||||||
| 200 ms | 0.432 | 0.274 | 0.306 | ||||||||||
| 250 ms | 0.447 | 0.303 | 0.316 | 0.372 | |||||||||
| 1RM | 0.426 | 0.267 | 0.300 | 0.407 | −0.487 | 0.511 | 0.509 | −0.500 | 0.512 | ||||
| High-Power Group (N = 30) | |||||||||||||
| CMJP | 0.645 | 0.477 | 0.320 | 0.400 | 0.497 | 0.466 | 0.500 | 0.421 | |||||
| MIF | 0.500 | 0.480 | 0.499 | −0.575 | 0.574 | 0.538 | −0.672 | 0.624 | 0.703 | ||||
| RFD | 150 ms | 0.514 | 0.347 | 0.324 | |||||||||
| 200 ms | 0.537 | 0.333 | 0.324 | 0.375 | |||||||||
| 250 ms | 0.567 | 0.378 | 0.360 | 0.400 | |||||||||
| 1RM | 0.612 | 0.389 | 0.500 | 0.577 | 0.451 | −0.598 | 0.630 | 0.599 | −0.601 | 0.621 | 0.579 | ||
| Low-Power Group (N = 30) | |||||||||||||
| CMJP | 0.444 | 0.366 | |||||||||||
| MIF | 0.312 | 0.290 | |||||||||||
| RFD | 250 ms | 0.399 | |||||||||||
| 1RM | 0.326 | ||||||||||||
| Performance Parameter (R2/p) | Lower Extremities Lean Body Mass | Fascicle Length | Type IIa Percentage Cross-Sectional Area | Type IIx Percentage Cross-Sectional Area | |
|---|---|---|---|---|---|
| Countermovement Jump Max Power (0.720/<0.001) | B | 0.621 | 0.481 | 0.384 | 0.548 |
| p | <0.001 | 0.002 | 0.029 | 0.004 | |
| Max Isometric Force (0.692/<0.001) | B | 0.589 | 0.424 | 0.625 | 0.384 |
| p | <0.001 | <0.001 | <0.001 | 0.013 | |
| Half Squat Maximum Strength (0.632/<0.001) | B | 0.712 | 0.399 | 0.521 | 0.412 |
| p | <0.001 | 0.011 | <0.001 | <0.001 |
| Vastuls Lateralis Fiber Type Composition | ||||||
|---|---|---|---|---|---|---|
| Cross Sectional Area | Percentage Cross-Sectional Area | |||||
| Vastuls Lateralis Architecture | I | IIa | IIx | I | IIa | IIx |
| All Participants (N = 60) | ||||||
| Muscle Thickness | 0.312 | 0.294 | −0.400 | 0.368 | 0.325 | |
| Pennation Angle | 0.300 | 0.314 | 0.322 | 0.300 | ||
| Fascicle Length | −0.333 | 0.321 | 0.378 | |||
| High Power Group (N = 30) | ||||||
| Muscle Thickness | 0.425 | 0.471 | −0.500 | 0.523 | 0.500 | |
| Pennation Angle | 0.392 | 0.400 | 0.491 | 0.420 | ||
| Fascicle Length | −0.414 | 0.499 | 0.522 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mpampoulis, T.; Methenitis, S.; Papadopoulos, C.; Papadimas, G.; Spiliopoulou, P.; Stasinaki, A.-N.; Bogdanis, G.C.; Karampatsos, G.; Terzis, G. Weak Association between Vastus Lateralis Muscle Fiber Composition and Fascicle Length in Young Untrained Females. Sports 2021, 9, 56. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9050056
Mpampoulis T, Methenitis S, Papadopoulos C, Papadimas G, Spiliopoulou P, Stasinaki A-N, Bogdanis GC, Karampatsos G, Terzis G. Weak Association between Vastus Lateralis Muscle Fiber Composition and Fascicle Length in Young Untrained Females. Sports. 2021; 9(5):56. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9050056
Chicago/Turabian StyleMpampoulis, Thomas, Spyridon Methenitis, Constantinos Papadopoulos, Giorgos Papadimas, Polyxeni Spiliopoulou, Angeliki-Nikoletta Stasinaki, Gregory C. Bogdanis, Giorgos Karampatsos, and Gerasimos Terzis. 2021. "Weak Association between Vastus Lateralis Muscle Fiber Composition and Fascicle Length in Young Untrained Females" Sports 9, no. 5: 56. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9050056