Accurate Diagnosis of Small Ruminant Lentivirus Infection Is Needed for Selection of Resistant Sheep through TMEM154 E35K Genotyping

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
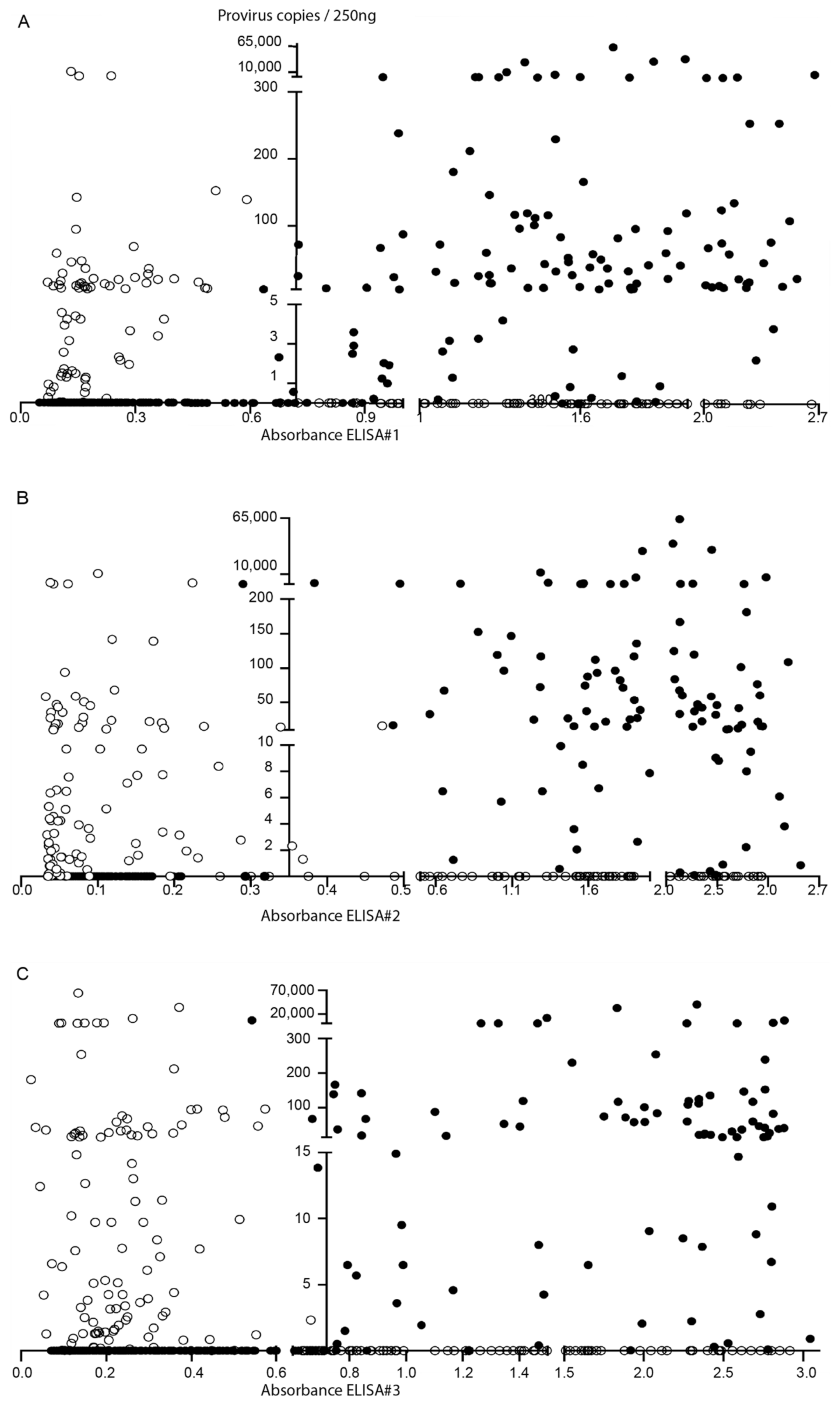
2.1. Serodiagnosis
2.2. Molecular Diagnosis
2.3. TMEM154 Genotyping
2.4. TMEM154 E35K Association with SRLV Infection Status
3. Discussion
4. Materials and Methods
4.1. Animals and Samples
4.2. Serological Survey
4.3. DNA Extraction and Quantification
4.4. SRLV Molecular Diagnosis
4.5. TMEM154 Genotyping
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Minguijón, E.; Reina, R.; Pérez, M.; Polledo, L.; Villoria, M.; Ramírez, H.; Leginagoikoa, I.; Badiola, J.J.; García-Marín, J.F.; de Andrés, D.; et al. Small ruminant lentivirus infections and diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef]
- Preziuso, S.; Renzoni, G.; Allen, T.E.; Taccini, E.; Rossi, G.; Demartini, J.C.; Braca, G. Colostral transmission of maedi visna virus: Sites of viral entry in lambs born from experimentally infected ewes. Vet. Microbiol. 2004, 104, 157–164. [Google Scholar] [CrossRef]
- Blacklaws, B.A.; Berriatua, E.; Torsteinsdottir, S.; Watt, N.J.; De Andres, D.; Klein, D.; Harkiss, G.D. Transmission of small ruminant lentiviruses. Vet. Microbiol. 2004, 101, 199–208. [Google Scholar] [CrossRef]
- Pérez, M.; Biescas, E.; de Andrés, X.; Leginagoikoa, I.; Salazar, E.; Berriatua, E.; Reina, R.; Bolea, R.; de Andrés, D.; Juste, R.A.; et al. Visna/maedi virus serology in sheep: Survey, risk factors and implementation of a successful control programme in Aragón (Spain). Vet. J. 2010, 186, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Herrmann-Hoesing, L.M. Diagnostic assays used to control small ruminant lentiviruses. J. Vet. Diagn. Investig. 2010, 22, 843–855. [Google Scholar] [CrossRef] [Green Version]
- Kalogianni, A.I.; Bossis, I.; Ekateriniadou, L.V.; Gelasakis, A.I. Etiology, Epizootiology and Control of Maedi-Visna in Dairy Sheep: A Review. Animals 2020, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Echeverría, I.; De Miguel, R.; De Pablo-Maiso, L.; Glaria, I.; Benito, A.A.; De Blas, I.; De Andrés, D.; Luján, L.; Reina, R. Multi-Platform Detection of Small Ruminant Lentivirus Antibodies and Provirus as Biomarkers of Production Losses. Front. Vet. Sci. 2020, 7, 182. [Google Scholar] [CrossRef]
- Lacerenza, D.; Giammarioli, M.; Grego, E.; Marini, C.; Profiti, M.; Rutili, D.; Rosati, S. Antibody response in sheep experimentally infected with different small ruminant lentivirus genotypes. Vet. Immunol. Immunopathol. 2006, 112, 264–271. [Google Scholar] [CrossRef]
- Cardinaux, L.; Zahno, M.L.; Deubelbeiss, M.; Zanoni, R.; Vogt, H.R.; Bertoni, G. Virological and phylogenetic characterization of attenuated small ruminant lentivirus isolates eluding efficient serological detection. Vet. Microbiol. 2013, 162, 572–581. [Google Scholar] [CrossRef]
- Ritchie, C.; Hosie, B. Concern over maedi visna breakdowns. Vet. Rec. 2014, 175, 50–51. [Google Scholar] [CrossRef]
- Ritchie, C.; Hosie, B. Health schemes: Increase in maedi-visna breakdowns. Vet. Rec. 2010, 167, 389. [Google Scholar]
- Kaba, J.; Czopowicz, M.; Ganter, M.; Nowicki, M.; Witkowski, L.; Nowicka, D.; Szaluś-Jordanow, O. Risk factors associated with seropositivity to small ruminant lentiviruses in goat herds. Res. Vet. Sci. 2013, 94, 225–227. [Google Scholar] [CrossRef]
- Brinkhof, J.M.A.; Houwers, D.J.; Moll, L.; Dercksen, D.; van Maanen, C. Diagnostic performance of ELISA and PCR in identifying SRLV-infected sheep and goats using serum, plasma and milk samples and in early detection of infection in dairy flocks through bulk milk testing. Vet. Microbiol. 2010, 142, 193–198. [Google Scholar] [CrossRef]
- Marinho, R.C.; Martins, G.R.; Souza, K.C.; Sousa, A.L.M.; Silva, S.T.C.; Nobre, J.A.; Teixeira, M.F.S. Duplex nested-PCR for detection of small ruminant lentiviruses. Brazilian J. Microbiol. 2018, 49, 83–92. [Google Scholar] [CrossRef]
- Chassalevris, T.; Chaintoutis, S.C.; Apostolidi, E.D.; Giadinis, N.D.; Vlemmas, I.; Brellou, G.D.; Dovas, C.I. A highly sensitive semi-nested real-time PCR utilizing oligospermine-conjugated degenerate primers for the detection of diverse strains of small ruminant lentiviruses. Mol. Cell. Probes 2020, 51, 101528. [Google Scholar] [CrossRef]
- Dolfini, T.; Conrad, L.F.; Flores, I.V.C.; Ravazzolo, A.P. Comparison of primer pairs: Greater degeneracy improves small ruminant lentivirus (SRLVs) detection by seminested PCR. Small Rumin. Res. 2015, 123, 189–192. [Google Scholar] [CrossRef]
- Eltahir, Y.M.; Dovas, C.I.; Papanastassopoulou, M.; Koumbati, M.; Giadinis, N.; Verghese-Nikolakaki, S.; Koptopoulos, G. Development of a semi-nested PCR using degenerate primers for the generic detection of small ruminant lentivirus proviral DNA. J. Virol. Methods 2006, 135, 240–246. [Google Scholar] [CrossRef]
- Heaton, M.P.; Clawson, M.L.; Chitko-Mckown, C.G.; Leymaster, K.A.; Smith, T.P.L.; Harhay, G.P.; White, S.N.; Herrmann-Hoesing, L.M.; Mousel, M.R.; Lewis, G.S.; et al. Reduced Lentivirus Susceptibility in Sheep with TMEM154 Mutations. PLoS Genet. 2012, 8, e1002467. [Google Scholar] [CrossRef] [Green Version]
- Heaton, M.P.; Kalbfleisch, T.S.; Petrik, D.T.; Simpson, B.; Kijas, J.W.; Clawson, M.L.; Chitko-McKown, C.G.; Harhay, G.P.; Leymaster, K.A. Genetic Testing for TMEM154 Mutations Associated with Lentivirus Susceptibility in Sheep. PLoS ONE 2013, 8, e55490. [Google Scholar] [CrossRef] [Green Version]
- White, S.N.; Mousel, M.R.; Herrmann-Hoesing, L.M.; Reynolds, J.O.; Leymaster, K.A.; Neibergs, H.L.; Lewis, G.S.; Knowles, D.P. Genome-Wide Association Identifies Multiple Genomic Regions Associated with Susceptibility to and Control of Ovine Lentivirus. PLoS ONE 2012, 7, e47829. [Google Scholar] [CrossRef] [Green Version]
- Cecchi, F.; Dadousis, C.; Bozzi, R.; Fratini, F.; Russo, C.; Bandecchi, P.; Cantile, C.; Mazzei, M. Genome scan for the possibility of identifying candidate resistance genes for goat lentiviral infections in the Italian Garfagnina goat breed. Trop. Anim. Health Prod. 2019, 51, 729–733. [Google Scholar] [CrossRef]
- Alshanbari, F.A.; Mousel, M.R.; Reynolds, J.O.; Herrmann-Hoesing, L.M.; Highland, M.A.; Lewis, G.S.; White, S.N. Mutations in Ovis aries TMEM154 are associated with lower small ruminant lentivirus proviral concentration in one sheep flock. Anim. Genet. 2014, 45, 565–571. [Google Scholar] [CrossRef]
- Leymaster, K.A.; Chitko-McKown, C.G.; Clawson, M.L.; Harhay, G.P.; Heaton, M.P. Effects of TMEM154 haplotypes 1 and 3 on susceptibility to ovine progressive pneumonia virus following natural exposure in sheep1,2,3. J. Anim. Sci. 2013, 91, 5114–5121. [Google Scholar] [CrossRef]
- Molaee, V.; Eltanany, M.; Lühken, G. First survey on association of TMEM154 and CCR5 variants with serological maedi-visna status of sheep in German flocks. Vet. Res. 2018, 49, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Molaee, V.; Otarod, V.; Abdollahi, D.; Lühken, G. Lentivirus Susceptibility in Iranian and German Sheep Assessed by Determination of TMEM154 E35K. Animals 2019, 9, 685. [Google Scholar] [CrossRef] [Green Version]
- Yaman, Y.; Keleş, M.; Aymaz, R.; Sevim, S.; Sezenler, T.; Önaldı, A.T.; Kaptan, C.; Başkurt, A.; Koncagül, S.; Öner, Y.; et al. Association of TMEM154 variants with visna/maedi virus infection in Turkish sheep. Small Rumin. Res. 2019, 177, 61–67. [Google Scholar] [CrossRef]
- Andrésdóttir, V.; Skraban, R.; Matthíasdóttir, S.; Lutley, R.; Agnarsdóttir, G.; Thorsteinsdóttir, H. Selection of antigenic variants in maedi-visna virus infection. J. Gen. Virol. 2002, 83, 2543–2551. [Google Scholar] [CrossRef]
- Michiels, R.; Adjadj, N.R.; De Regge, N. Phylogenetic Analysis of Belgian Small Ruminant Lentiviruses Supports Cross Species Virus Transmission and Identifies New Subtype B5 Strains. Pathogens 2020, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Michiels, R.; Van Mael, E.; Quinet, C.; Adjadj, N.; Cay, A.; De Regge, N. Comparative Analysis of Different Serological and Molecular Tests for the Detection of Small Ruminant Lentiviruses (SRLVs) in Belgian Sheep and Goats. Viruses 2018, 10, 696. [Google Scholar] [CrossRef] [Green Version]
- Sanjosé, L.; Pinczowski, P.; Crespo, H.; Pérez, M.; Glaria, I.; Gimeno, M.; de Andrés, D.; Amorena, B.; Luján, L.; Reina, R. Diagnosing infection with small ruminant lentiviruses of genotypes A and B by combining synthetic peptides in ELISA. Vet. J. 2015, 204, 88–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reina, R.; Grego, E.; Profiti, M.; Glaria, I.; Robino, P.; Quasso, A.; Amorena, B.; Rosati, S. Development of specific diagnostic test for small ruminant lentivirus genotype E. Vet. Microbiol. 2009, 138, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leginagoikoa, I.; Juste, R.A.; Barandika, J.; Amorena, B.; De Andrés, D.; Luján, L.; Badiola, J.; Berriatua, E. Extensive rearing hinders Maedi-Visna Virus (MVV) infection in sheep. Vet. Res. 2006, 37, 767–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, H.; Reina, R.; Bertolotti, L.; Cenoz, A.; Hernández, M.M.; San Román, B.; Glaria, I.; de Andrés, X.; Crespo, H.; Jáuregui, P.; et al. Study of compartmentalization in the visna clinical form of small ruminant lentivirus infection in sheep. BMC Vet. Res. 2012, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Colitti, B.; Coradduzza, E.; Puggioni, G.; Capucchio, M.T.; Reina, R.; Bertolotti, L.; Rosati, S. A new approach for small ruminant lentivirus full genome characterization revealed the circulation of divergent strains. PLoS ONE 2019, 14, e0212585. [Google Scholar] [CrossRef] [PubMed]
- Pisoni, G.; Bertoni, G.; Puricelli, M.; Maccalli, M.; Moroni, P. Demonstration of Coinfection with and Recombination by Caprine Arthritis-Encephalitis Virus and Maedi-Visna Virus in Naturally Infected Goats. J. Virol. 2007, 81, 4948–4955. [Google Scholar] [CrossRef] [Green Version]
- Rosati, S.; Mannelli, A.; Merlo, T.; Ponti, N. Characterization of the immunodominant cross-reacting epitope of visna maedi virus and caprine arthritis-encephalitis virus capsid antigen. Virus Res. 1999, 61, 177–183. [Google Scholar] [CrossRef]
- Kwang, J.; Keen, J.; Cutlip, R.C.; Kim, H.S.; de la Concha-Bermejillo, A. Serological diagnosis of caprine lentivirus infection by recombinant immunoassays. Small Rumin. Res. 1995, 16, 171–177. [Google Scholar] [CrossRef]
- Varea, R.; Monleón, E.; Pacheco, C.; Luján, L.; Bolea, R.; Vargas, M.A.; Van Eynde, G.; Saman, E.; Dickson, L.; Harkiss, G.; et al. Early detection of maedi-visna (ovine progressive pneumonia) virus seroconversion in field sheep samples. J. Vet. Diagn. Investig. 2001, 13, 301–307. [Google Scholar] [CrossRef]
- Saman, E.; Van Eynde, G.; Lujan, L.; Extramiana, B.; Harkiss, G.; Tolari, F.; Gonzàlez, L.; Amorena, B.; Watt, N.; Badiola, J. A new sensitive serological assay for detection of lentivirus infections in small ruminants. Clin. Diagn. Lab. Immunol. 1999, 6, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Lühken, G. Phylogenetic analysis of small ruminant lentiviruses in Germany and Iran suggests their expansion with domestic sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef] [Green Version]
- Gjerset, B.; Storset, A.K.; Rimstad, E. Genetic diversity of small-ruminant lentiviruses: Characterization of Norwegian isolates of Caprine arthritis encephalitis virus. J. Gen. Virol. 2006, 87, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Grego, E.; Bertolotti, L.; Quasso, A.; Profiti, M.; Lacerenza, D.; Muz, D.; Rosati, S. Genetic characterization of small ruminant lentivirus in Italian mixed flocks: Evidence for a novel genotype circulating in a local goat population. J. Gen. Virol. 2007, 88, 3423–3427. [Google Scholar] [CrossRef] [PubMed]
- Kuhar, U.; Barlič-Maganja, D.; Grom, J. Development and validation of TaqMan probe based real time PCR assays for the specific detection of genotype A and B small ruminant lentivirus strains. BMC Vet. Res. 2013, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barquero, N.; Gomez-Lucia, E.; Arjona, A.; Toural, C.; Heras, A.; Fernández-Garayzabal, J.; Domenech, A. Evolution of Specific Antibodies and Proviral DNA in Milk of Small Ruminants Infected by Small Ruminant Lentivirus. Viruses 2013, 5, 2614–2623. [Google Scholar] [CrossRef] [PubMed]
- Leginagoikoa, I.; Daltabuit-Test, M.; Álvarez, V.; Arranz, J.; Juste, R.A.; Amorena, B.; de Andrés, D.; Luján, L.L.; Badiola, J.J.; Berriatua, E. Horizontal Maedi-Visna virus (MVV) infection in adult dairy-sheep raised under varying MVV-infection pressures investigated by ELISA and PCR. Res. Vet. Sci. 2006, 80, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Juste, R.A.; Villoria, M.; Leginagoikoa, I.; Ugarte, E.; Minguijon, E. Milk production losses in Latxa dairy sheep associated with small ruminant lentivirus infection. Prev. Vet. Med. 2020, 176, 104886. [Google Scholar] [CrossRef] [PubMed]
- Clawson, M.L.; Redden, R.; Schuller, G.; Heaton, M.P.; Workman, A.; Chitko-Mckown, C.G.; Smith, T.P.; Leymaster, K.A. Genetic subgroup of small ruminant lentiviruses that infects sheep homozygous for TMEM154 frameshift deletion mutation A4Δ53. Vet. Res. 2015, 46, 22. [Google Scholar] [CrossRef] [Green Version]
- Muz, D.; Oǧuzoǧlu, T.Ç.; Rosati, S.; Reina, R.; Bertolotti, L.; Burgu, I. First molecular characterization of visna/maedi viruses from naturally infected sheep in Turkey. Arch. Virol. 2013, 158, 559–570. [Google Scholar] [CrossRef]
- Li, X.; Howard, T.D.; Ampleford, E.J.; Peters, S.P.; Bleecker, E.R.; Meyers, D.A. Genome-wide Association Study of Asthma Severity Using Proportional Odds Model Identifies TMEM154. In Proceedings of the American Thoracic Society International Conference Meetings Abstracts American Thoracic Society International Conference Meetings, New Orleans, LA, USA, 14–19 May 2010; p. A3728. [Google Scholar]
- Mahajan, A.; Go, M.J.; Zhang, W.; Below, J.E.; Gaulton, K.J.; Ferreira, T.; Horikoshi, M.; Johnson, A.D.; Ng, M.C.Y.; Prokopenko, I.; et al. Genome-wide trans-ancestry meta-analysis provides insight into the genetic architecture of type 2 diabetes susceptibility. Nat. Genet. 2014, 46, 234–244. [Google Scholar] [CrossRef]
- Harder, M.N.; Appel, E.V.R.; Grarup, N.; Gjesing, A.P.; Ahluwalia, T.S.; Jørgensen, T.; Christensen, C.; Brandslund, I.; Linneberg, A.; Sørensen, T.I.A.; et al. The Type 2 Diabetes Risk Allele of TMEM154-rs6813195 Associates with Decreased Beta Cell Function in a Study of 6,486 Danes. PLoS ONE 2015, 10, e0120890. [Google Scholar] [CrossRef] [Green Version]
- Matsuba, R.; Imamura, M.; Tanaka, Y.; Iwata, M.; Hirose, H.; Kaku, K.; Maegawa, H.; Watada, H.; Tobe, K.; Kashiwagi, A.; et al. Replication study in a japanese population of six susceptibility loci for type 2 diabetes originally identified by a transethnic meta-analysis of genome-wide association studies. PLoS ONE 2016, 11, e0154093. [Google Scholar] [CrossRef] [PubMed]



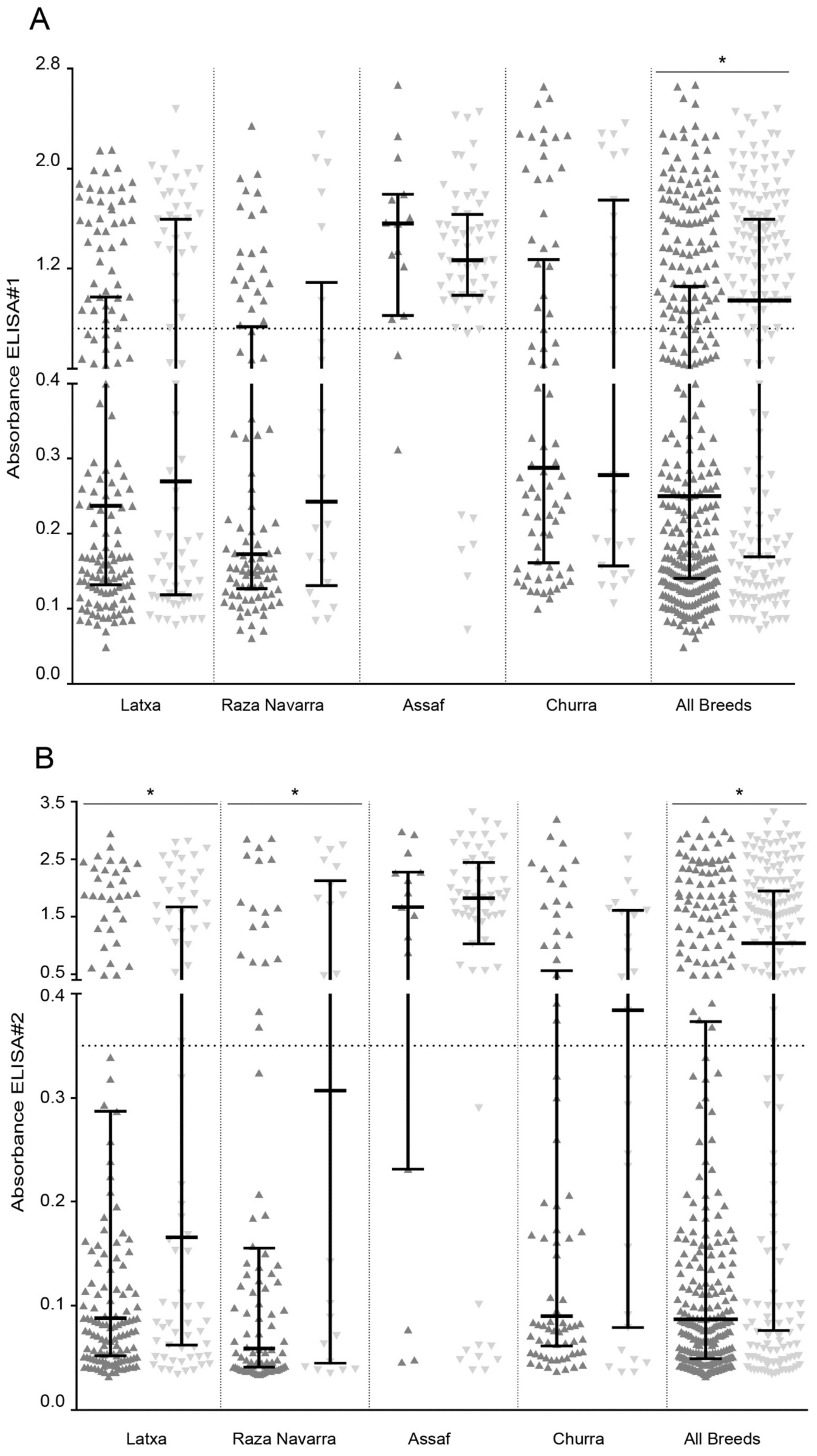
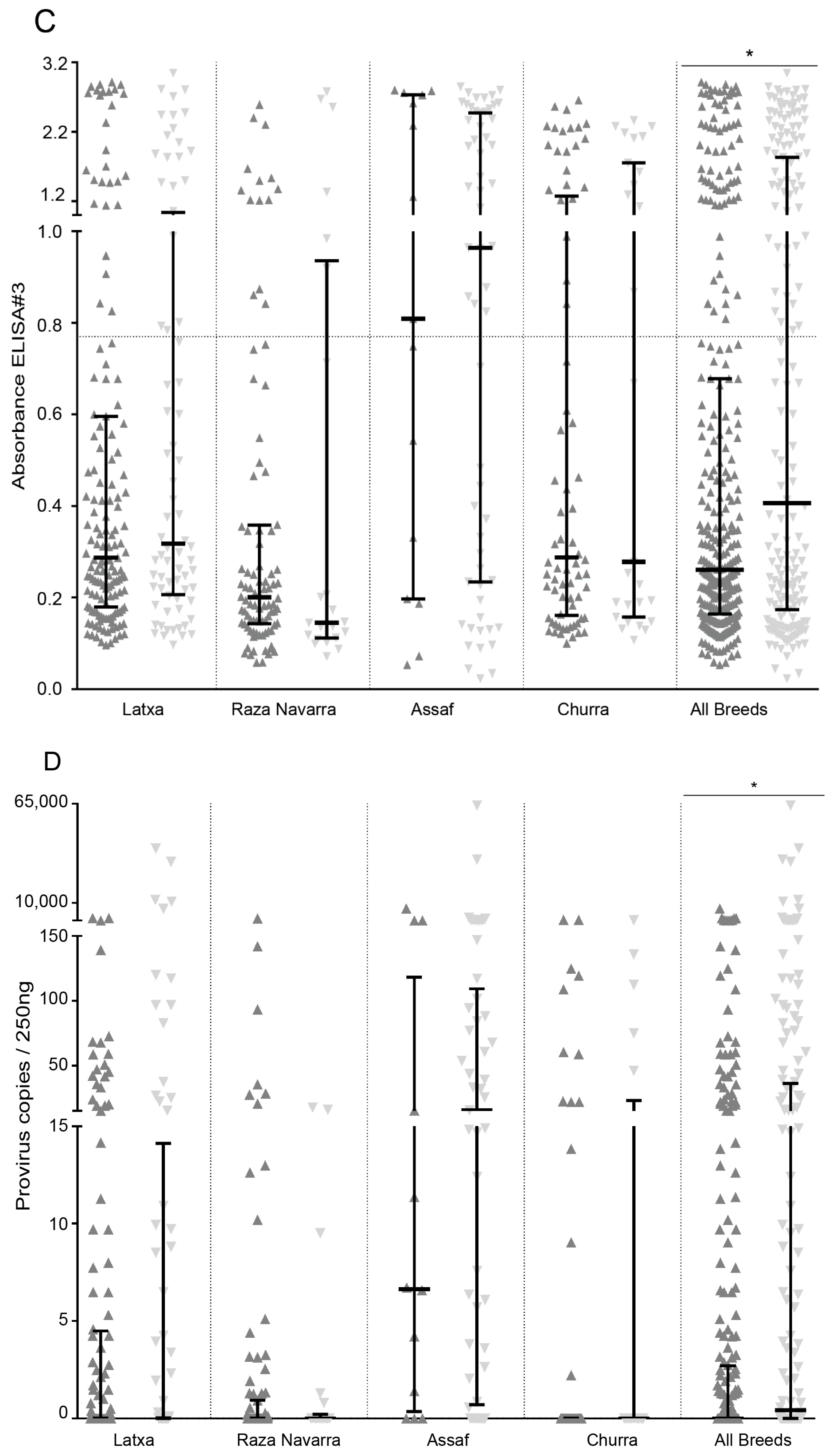
 ) or E/K and E/E (
) or E/K and E/E (  ) and analyzed by ELISA and qPCR. Samples were grouped by individual breeds and combined (All breeds). Average cut-off values of individual ELISA are represented as a horizontal dotted line (* Mann–Whitney, p < 0.05).
) or E/K and E/E ( ) and analyzed by ELISA and qPCR. Samples were grouped by individual breeds and combined (All breeds). Average cut-off values of individual ELISA are represented as a horizontal dotted line (* Mann–Whitney, p < 0.05).
) and analyzed by ELISA and qPCR. Samples were grouped by individual breeds and combined (All breeds). Average cut-off values of individual ELISA are represented as a horizontal dotted line (* Mann–Whitney, p < 0.05).
) or E/K and E/E ( ) and analyzed by ELISA and qPCR. Samples were grouped by individual breeds and combined (All breeds). Average cut-off values of individual ELISA are represented as a horizontal dotted line (* Mann–Whitney, p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TEST | Raza Navarra | Latxa Navarra | Assaf | Churra | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Positive | Efficiency % | n | Positive | Efficiency % | n | Positive | Efficiency % | n | Positive | Efficiency % | |||||
| n | % | n | % | n | % | n | % | |||||||||
| ELISA#1 | 114 | 29 | 25.4 | 55.8 | 194 | 76 | 39.2 | 73.8 | 74 | 66 | 89.2 | 91.7 | 101 | 32 | 31.7 | 47.8 |
| ELISA#2 | 114 | 28 | 24.6 | 53.9 | 194 | 60 | 30.9 | 58.3 | 74 | 61 | 82.4 | 84.7 | 101 | 42 | 41.6 | 62.7 |
| ELISA#3 | 114 | 20 | 17.5 | 38.5 | 194 | 60 | 30.9 | 58.3 | 74 | 46 | 62.2 | 63.9 | 101 | 61 | 60.4 | 91.0 |
| Total ELISA | 114 | 52 | 45.6 | 100.0 | 194 | 103 | 53.1 | 100.0 | 74 | 72 | 97.3 | 100.0 | 101 | 67 | 66.3 | 100.0 |
| TEST | Raza Navarra | Latxa | Assaf | Churra | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Positive | Efficiency % | n | Positive | Efficiency % | n | Positive | Efficiency % | n | Positive | Efficiency % | |||||
| n | % | n | % | n | % | n | % | |||||||||
| qPCR | 111 | 33 | 29.7 | 45.2 | 191 | 90 | 47.1 | 65.7 | 74 | 57 | 77.0 | 79.2 | 82 | 18 | 22.0 | 32.1 |
| Total infected | 114 | 75 | 65.8 | 100.0 | 194 | 139 | 71.6 | 100.0 | 74 | 72 | 97.3 | 100.0 | 101 | 67 | 66.3 | 100.0 |
| Probes/Primers Sequences | Product Size (Base Pairs) | Purpose |
|---|---|---|
| Fw 5′-CTGCCTTTGTGGGAGATTTA-3′ | 335 | Amplification and sequencing for verification of genotyping results |
| Rv 5′-TTCTGTGGTCACTGAAGCAA-3′ | ||
| Fw 5′-TTCGTCTCCATGACAAGTCTCAAT-3′ | 121 | Determination of nucleotide substitution G/A, resulting in amino acid substitution E35K. |
| Rv 5′-GCTTAGGGCCTCTGACTCTTCA-3′ | ||
| HEX-AGGACACAGAACTGT-BHQ-1 | ||
| 6-FAM-AGGACACAAAACTGT-BHQ-1 |
| Genotype | Raza Navarra | Latxa | Assaf | Churra | ||||
|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | n | % | |
| K/K | 92 | 80.7 | 134 | 69.1 | 15 | 20.3 | 75 | 74.3 |
| E/K | 18 | 15.8 | 56 | 28.9 | 32 | 43.2 | 24 | 23.8 |
| E/E | 4 | 3.5 | 4 | 2.1 | 27 | 36.5 | 2 | 2.0 |
| Total | 114 | 100 | 194 | 100 | 74 | 100 | 101 | 100 |
| TEST | SRLV | Raza Navarra | Latxa Navarra | Assaf | Churra | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TMEM154 Genotype (%) | TMEM154 Genotype (%) | TMEM154 Genotype (%) | TMEM154 Genotype (%) | ||||||||||||||||||
| p | RR | P’ | p | RR | P’ | p | RR | P’ | p | RR | P’ | ||||||||||
| KK | EK/EE | (95%CI) | KK | EK/EE | (95%CI) | KK | EK/EE | (95%CI) | KK | EK/EE | (95%CI) | ||||||||||
| ELISA#1 | Negative | 69 | 16 | 0.792 | 1.09 (0.51–2.35) | 0.824 | 95 | 23 | <0.0001 | 2.12 (1.52–2.95) | <0.0001 | 2 | 6 | 0.66 | 1.04 (0.83–1.29) | 0.745 | 53 | 16 | 0.338 | 1.35 (0.74–2.48) | 0.326 |
| Positive | 23 | 6 | 39 | 37 | 13 | 53 | 21 | 10 | |||||||||||||
| ELISA#2 | Negative | 74 | 12 | 0.024 | 2.32 (1.25–4.31) | 0.008 | 111 | 23 | <0.0001 | 3.59 (2.36–5.48) | <0.0001 | 4 | 9 | 0.446 | 1.16 (0.84–1.60) | 0.381 | 50 | 9 | 0.005 | 2.02 (1.31–3.10) | 0.002 |
| Positive | 18 | 10 | 23 | 37 | 11 | 50 | 24 | 17 | |||||||||||||
| ELISA#3 | Negative | 78 | 16 | 0.214 | 1.79 (0.78–4.13) | 0.171 | 101 | 33 | 0.007 | 1.83 (1.22–2.75) | 0.0037 | 5 | 23 | 0.772 | 0.91 (0.60–1.38) | 0.674 | 31 | 9 | 0.644 | 1.11 (0.79–1.56) | 0.530 |
| Positive | 14 | 6 | 33 | 27 | 10 | 36 | 44 | 17 | |||||||||||||
| TOTALELISA | Negative | 52 | 10 | 0.475 | 1.25 (0.80–1.96) | 0.003 | 73 | 18 | 0.002 | 1.54 (1.20–1.98) | 0.0007 | 0 | 2 | 1 | 0.97 (0.92–1.01) | 0.157 | 28 | 6 | 0.232 | 1.23 (0.93–1.61) | 0.142 |
| Positive | 40 | 12 | 61 | 42 | 15 | 57 | 47 | 20 | |||||||||||||
| qPCR | Negative | 62 | 16 | 1 | 0.90 (0.42–1.91) | 0.781 | 72 | 29 | 0.437 | 1.15 (0.84–1.56) | 0.384 | 3 | 14 | 1 | 0.95 (0.71–1.27) | 0.747 | 48 | 16 | 0.770 | 1.12 (0.45–2.76) | 0.810 |
| Positive | 27 | 6 | 59 | 31 | 12 | 45 | 13 | 5 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez, H.; Echeverría, I.; Benito, A.A.; Glaria, I.; Benavides, J.; Pérez, V.; de Andrés, D.; Reina, R. Accurate Diagnosis of Small Ruminant Lentivirus Infection Is Needed for Selection of Resistant Sheep through TMEM154 E35K Genotyping. Pathogens 2021, 10, 83. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10010083
Ramírez H, Echeverría I, Benito AA, Glaria I, Benavides J, Pérez V, de Andrés D, Reina R. Accurate Diagnosis of Small Ruminant Lentivirus Infection Is Needed for Selection of Resistant Sheep through TMEM154 E35K Genotyping. Pathogens. 2021; 10(1):83. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10010083
Chicago/Turabian StyleRamírez, Hugo, Irache Echeverría, Alfredo A. Benito, Idoia Glaria, Julio Benavides, Valentín Pérez, Damián de Andrés, and Ramsés Reina. 2021. "Accurate Diagnosis of Small Ruminant Lentivirus Infection Is Needed for Selection of Resistant Sheep through TMEM154 E35K Genotyping" Pathogens 10, no. 1: 83. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10010083