
Tsetse Bloodmeal Analyses Incriminate the Common Warthog Phacochoerus africanus as an Important Cryptic Host of Animal Trypanosomes in Smallholder Cattle Farming Communities in Shimba Hills, Kenya

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
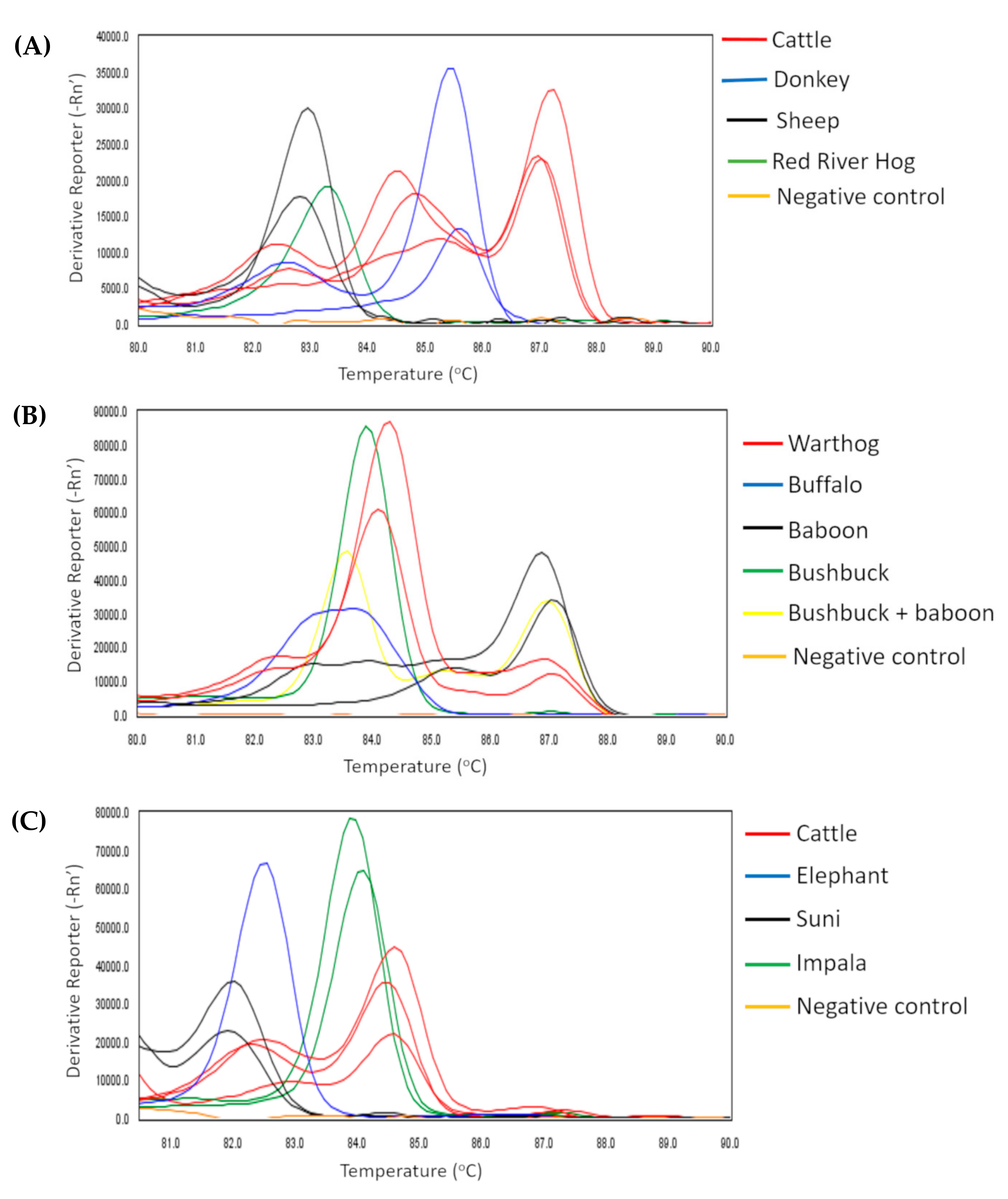
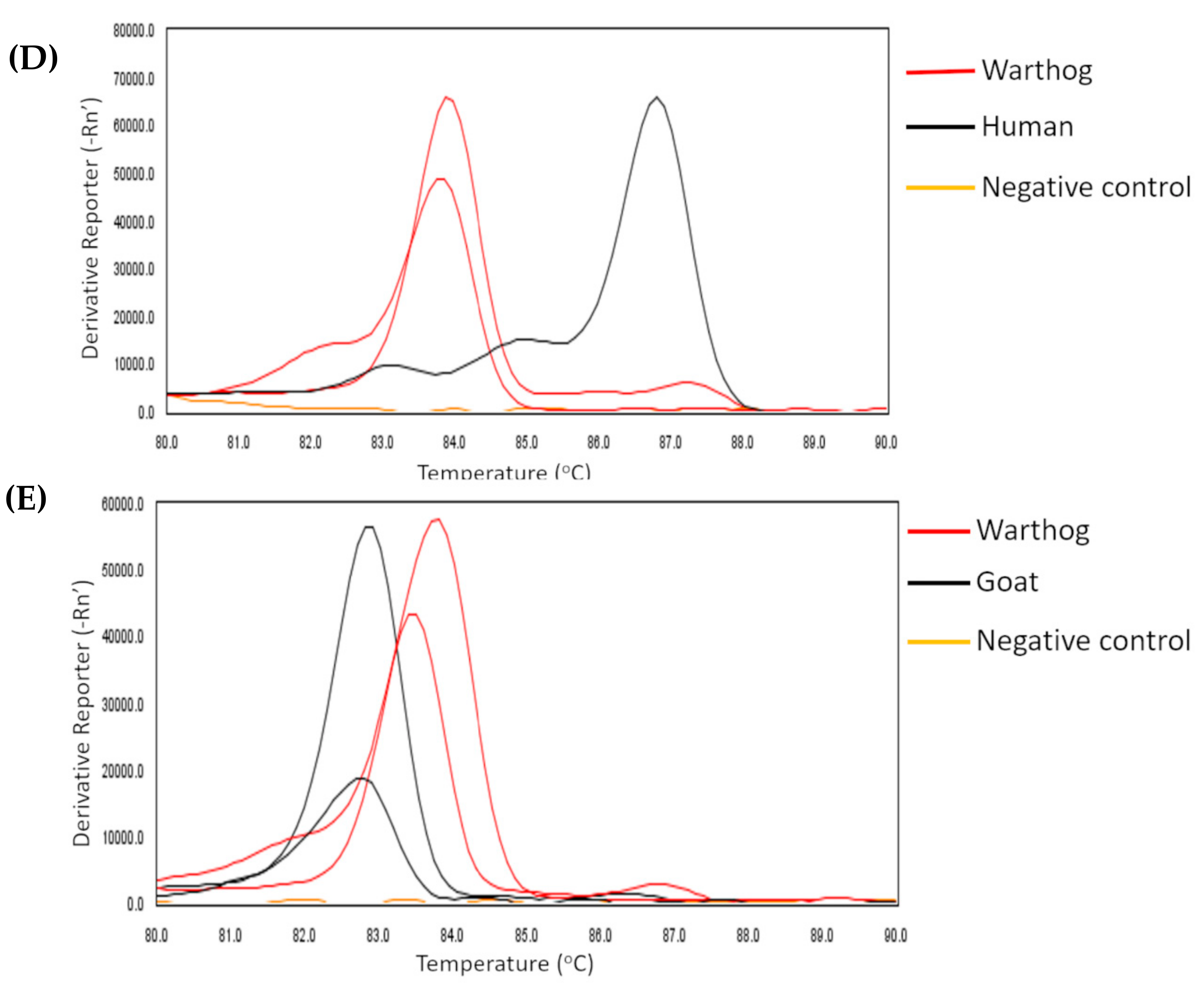
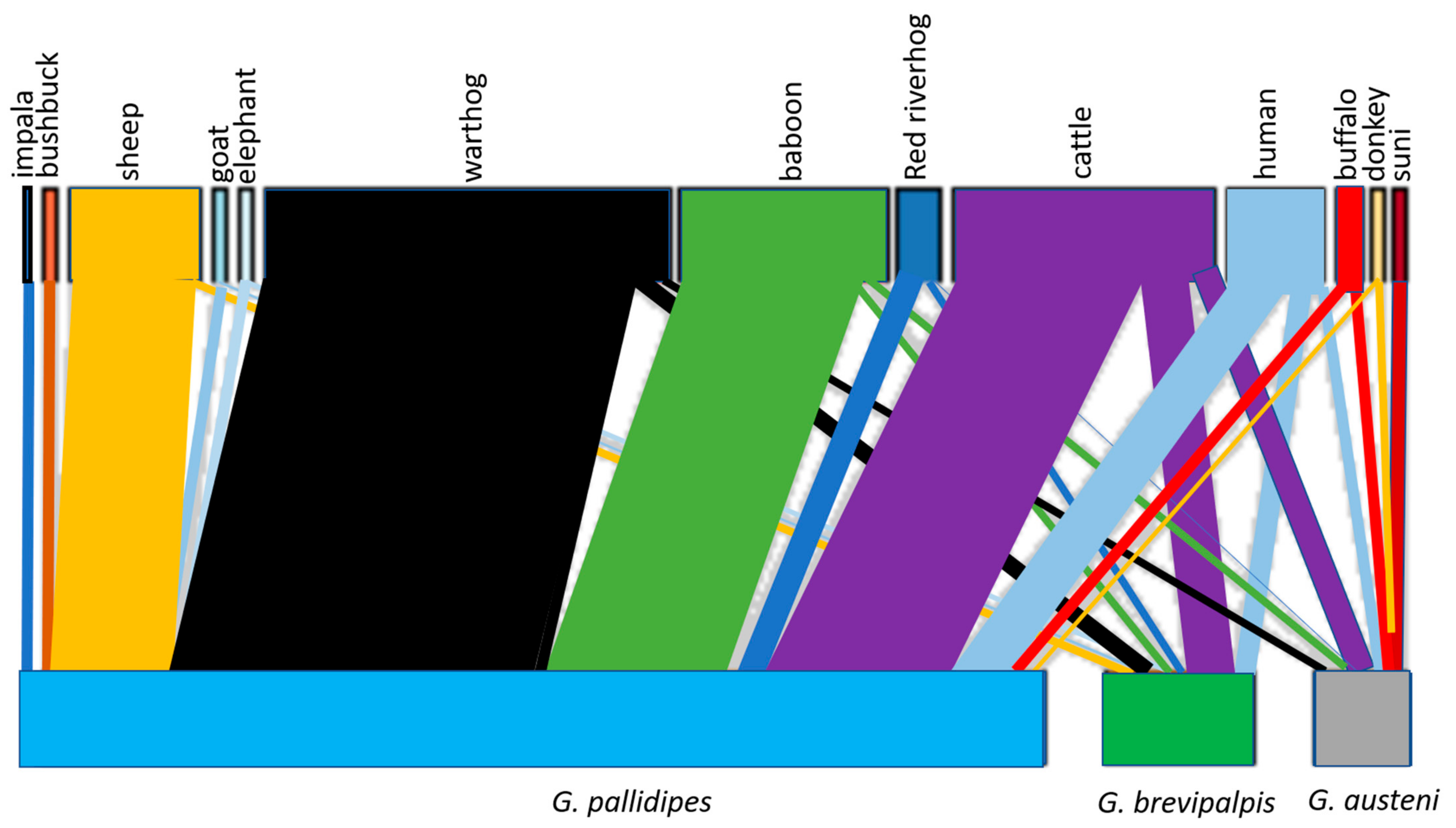
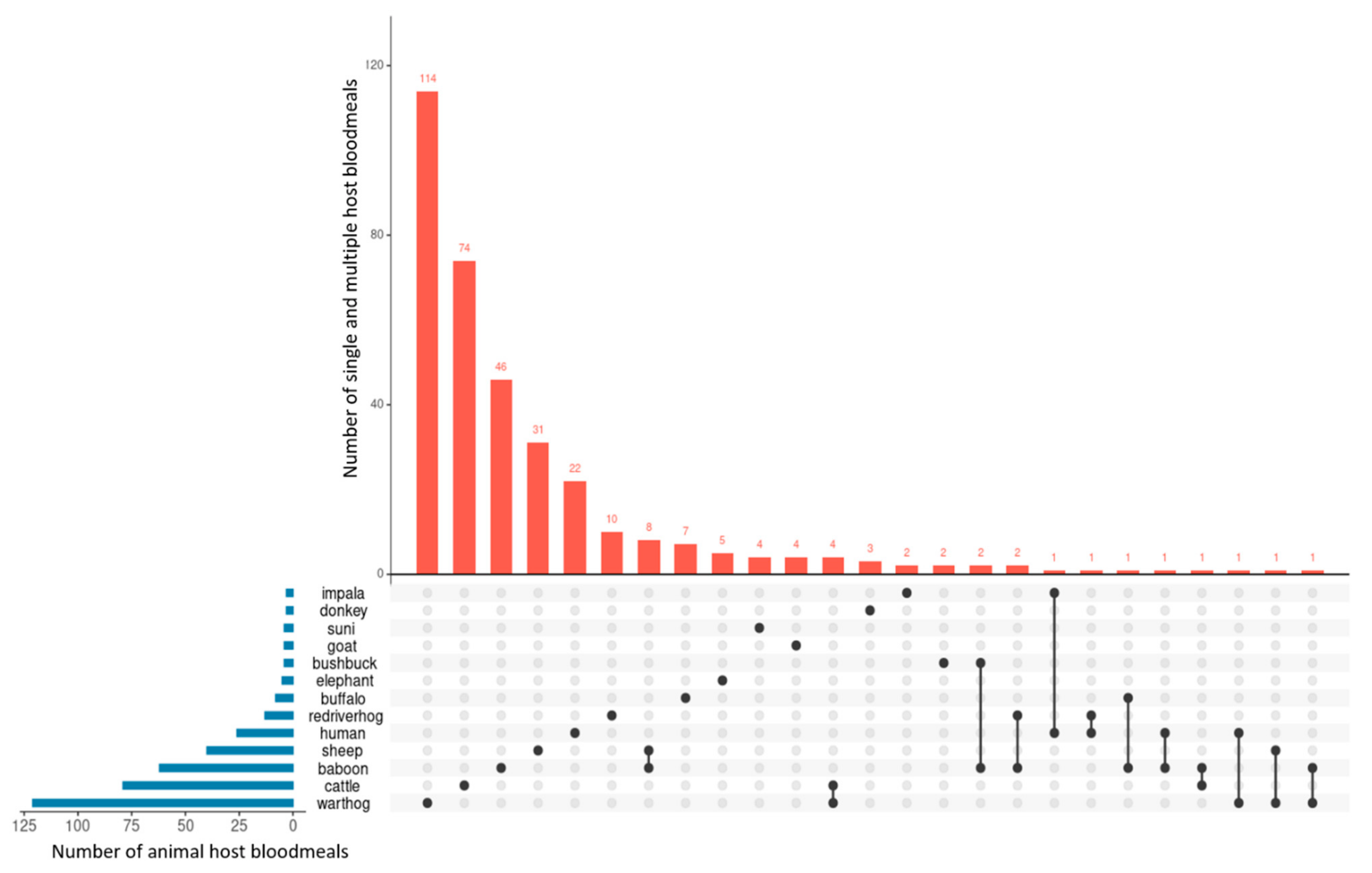
2.1. Animal Bloodmeals in Tsetse Flies
2.2. Trypanosome Infections in Blood-Fed Tsetse Flies
3. Discussion
4. Materials and Methods
4.1. Ethical Consent
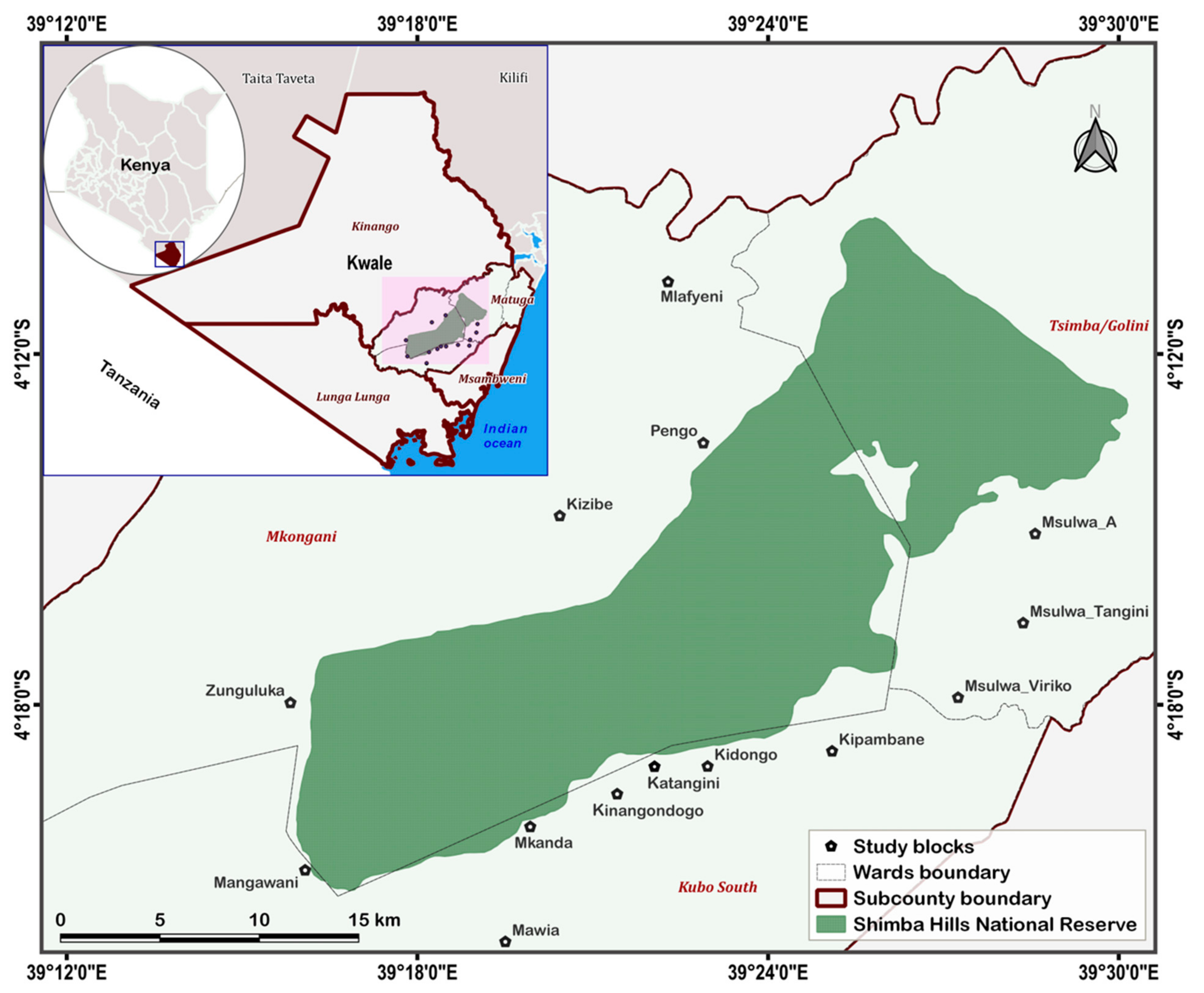
4.2. Study Area
4.3. Tsetse Fly Collection and Characterization
4.4. Identification of Vertebrate Bloodmeal Sources in Tsetse Flies
4.5. Molecular Identification of Trypanosomes in Tsetse Flies
4.6. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and disease emergence: Dynamics at the wildlife–livestock–human interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.E.; Mubanga, J.; Fevre, E.M.; Picozzi, K.; Eisler, M.C.; Thomas, R.; Welburn, S.C. Characterisation of the wildlife reservoir community for human and animal trypanosomiasis in the Luangwa Valley, Zambia. PLoS Negl. Trop. Dis. 2011, 5, e1211. [Google Scholar] [CrossRef] [Green Version]
- Auty, H.; Morrison, L.J.; Torr, S.J.; Lord, J. Transmission dynamics of Rhodesian sleeping sickness at the interface of wildlife and livestock areas. Trends Parasitol. 2016, 32, 608–621. [Google Scholar] [CrossRef] [PubMed]
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human african trypanosomiasis. Lancet 2017, 390, 2397–2409. [Google Scholar] [CrossRef]
- Diall, O.; Cecchi, G.; Wanda, G.; Argilés-Herrero, R.; Vreysen, M.J.; Cattoli, G.; Viljoen, G.J.; Mattioli, R.; Bouyer, J. Developing a progressive control pathway for African animal trypanosomosis. Trends Parasitol. 2017, 33, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Kaare, M.T.; Picozzi, K.; Mlengeya, T.; Fèvre, E.M.; Mellau, L.S.; Mtambo, M.M.; Cleaveland, S.; Welburn, S.C. Sleeping sickness—A re-emerging disease in the Serengeti? Travel Med. Infect. Dis. 2007, 5, 117–124. [Google Scholar] [CrossRef]
- Claxton, J.R.; Faye, J.A.; Rawlings, P. Trypanosome infections in warthogs (Phacochoerus aethiopicus) in the Gambia. Vet. Parasitol. 1992, 41, 179–187. [Google Scholar] [CrossRef]
- Muturi, C.N.; Ouma, J.O.; Malele, I.I.; Ngure, R.M.; Rutto, J.J.; Mithöfer, K.M.; Enyaru, J.; Masiga, D.K. Tracking the feeding patterns of tsetse flies (Glossina genus) by analysis of bloodmeals using mitochondrial cytochromes genes. PLoS ONE 2011, 6, e17284. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Siamudaala, V.; Munyeme, M.; Nalubamba, K.S. A review of ecological factors associated with the epidemiology of wildlife trypanosomiasis in the Luangwa and Zambezi valley ecosystems of Zambia. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 372523. [Google Scholar] [CrossRef]
- Squarre, D.; Hayashida, K.; Gaithuma, A.; Chambaro, H.; Kawai, N.; Moonga, L.; Namangala, B.; Sugimoto, C.; Yamagishi, J. Diversity of trypanosomes in wildlife of the Kafue ecosystem, Zambia. Int. J. Parasitol. Parasites Wildl. 2020, 12, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Makhulu, E.E.; Villinger, J.; Adunga, V.O.; Jeneby, M.M.; Kimathi, E.M.; Mararo, E.; Oundo, J.W.; Musa, A.A.; Wambua, L. Tsetse blood-meal sources, endosymbionts and trypanosome-associations in the Maasai Mara National Reserve, a wildlife-human-livestock interface. PLoS Negl. Trop. Dis. 2021, 15, e0008267. [Google Scholar] [CrossRef]
- Channumsin, M.; Ciosi, M.; Masiga, D.; Auty, H.; Turner, C.M.; Kilbride, E.; Mable, B.K. Blood meal analysis of tsetse flies (Glossina pallidipes: Glossinidae) reveals higher host fidelity on wild compared with domestic hosts. Wellcome Open Res. 2021, 6, 213. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Orindi, B.O.; Mbahin, N.; Andoke, J.A.; Muasa, P.N.; Mbuvi, D.M.; Muya, C.M.; Pickett, J.A.; Borgemeister, C.W. Protecting cows in small holder farms in East Africa from tsetse flies by mimicking the odor profile of a non-host bovid. PLoS Negl. Trop. Dis. 2017, 11, e0005977. [Google Scholar]
- Navia-Gine, W.G.; Loaiza, J.R.; Miller, M.J. Mosquito-host interactions during and after an outbreak of equine viral encephalitis in eastern Panama. PLoS ONE 2013, 8, e81788. [Google Scholar] [CrossRef]
- Escobar, D.; Ascencio, K.; Ortiz, A.; Palma, A.; Sánchez, A.; Fontecha, G. Blood meal sources of Anopheles spp. in malaria endemic areas of Honduras. Insects 2020, 11, 450. [Google Scholar] [CrossRef] [PubMed]
- Weitz, B. The feeding habits of Glossina. Bull. World Health Organ. 1963, 28, 711. [Google Scholar] [PubMed]
- Weitz, B.; Glasgow, J.P. The natural hosts of some species of Glossina in East Africa. Trans. R. Soc. Trop. Med. Hyg. 1956, 50, 593–612. [Google Scholar] [CrossRef] [Green Version]
- Auty, H.; Cleaveland, S.; Malele, I.; Masoy, J.; Lembo, T.; Bessell, P.; Torr, S.; Picozzi, K.; Welburn, S.C. Quantifying heterogeneity in host-vector contact: Tsetse (Glossina swynnertoni and G. pallidipes) host choice in Serengeti National Park, Tanzania. PLoS ONE 2016, 11, e0161291. [Google Scholar] [CrossRef] [PubMed]
- Lamprey, H.F.; Glasgow, J.P.; Lee-Jones, F.; Weitz, B. A simultaneous census of the potential and actual food sources of the tsetse fly Glossina swynnertoni Austen. J. Anim. Ecol. 1962, 31, 151–156. [Google Scholar] [CrossRef]
- Ngene, S.M.; Mukeka, J. Aerial Census of Elephants and Other Mammals in Shimba Hills, Mkongani, and Mwaluganje Conservation Areas; Kenya Wildlife Service: Nairobi, Kenya, 2012. [Google Scholar]
- De Beer, C.J.; Venter, G.J.; Potgieter, F.T. Developing Quality Control Procedures to Sustain a Supply of High Quality Blood for Mass Rearing Tsetse Flies; IAEA: Vienna, Austria, 2012. [Google Scholar]
- Späth, J. Natural host odours as possible attractants for Glossina tachinoides and G. longipalpis (Diptera: Glossinidae). Acta Trop. 1997, 68, 149–158. [Google Scholar] [CrossRef]
- Hargrove, J.W.; Vale, G.A. The effect of host odour concentration on catches of tsetse flies (Glossinidae) and other Diptera in the field. Bull. Entomol. Res. 1978, 68, 607–612. [Google Scholar] [CrossRef]
- Brightwell, B.; Dransfield, B.; Maudlin, I.; Stevenson, P.; Shaw, A. Reality vs. rhetoric–a survey and evaluation of tsetse control in East Africa. Agric. Hum. Values 2001, 18, 219–233. [Google Scholar] [CrossRef]
- Leak, S.G.; Ejigu, D.; Vreysen, M.J. Collection of Entomological Baseline Data for Tsetse Area-Wide Integrated Pest Management Programmes; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2008. [Google Scholar]
- Vale, G.A. The responses of tsetse flies (Diptera, Glossinidae) to mobile and stationary baits. Bull. Entomol. Res. 1974, 64, 545–588. [Google Scholar] [CrossRef]
- Rutto, J.J.; Karuga, J.W. Temporal and spatial epidemiology of sleeping sickness and use of geographical information system (GIS) in Kenya. J. Vector Borne Dis. 2009, 46, 18. [Google Scholar] [PubMed]
- Olaide, O.Y.; Tchouassi, D.P.; Yusuf, A.A.; Pirk, C.W.; Masiga, D.K.; Saini, R.K.; Torto, B. Zebra skin odor repels the savannah tsetse fly, Glossina pallidipes (Diptera: Glossinidae). PLoS Negl. Trop. Dis. 2019, 13, e0007460. [Google Scholar] [CrossRef] [Green Version]
- Bett, M.K.; Saini, R.K.; Hassanali, A. Repellency of tsetse-refractory waterbuck (Kobus defassa) body odour to Glossina pallidipes (Diptera: Glossinidae): Assessment of relative contribution of different classes and individual constituents. Acta Trop. 2015, 146, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Waage, J.K. How the zebra got its stripes-biting flies as selective agents in the evolution of zebra coloration. J. Entomol. Soc. S. Afr. 1981, 44, 351–358. [Google Scholar]
- Gibson, G. Do tsetse flies ‘see’ zebras? A field study of the visual response of tsetse to striped targets. Physiol. Entomol. 1992, 17, 141–147. [Google Scholar] [CrossRef]
- Caro, T.; Izzo, A.; Reiner, R.C.; Walker, H.; Stankowich, T. The function of zebra stripes. Nat. Commun. 2014, 5, 3535. [Google Scholar] [CrossRef]
- Gaithuma, A.; Yamagishi, J.; Hayashida, K.; Kawai, N.; Namangala, B.; Sugimoto, C. Blood meal sources and bacterial microbiome diversity in wild-caught tsetse flies. Sci. Rep. 2020, 10, 5005. [Google Scholar] [CrossRef] [Green Version]
- Ouso, D.O.; Otiende, M.Y.; Jeneby, M.M.; Oundo, J.W.; Bargul, J.L.; Miller, S.E.; Wambua, L.; Villinger, J. Three-gene PCR and high-resolution melting analysis for differentiating vertebrate species mitochondrial DNA for biodiversity research and complementing forensic surveillance. Sci. Rep. 2020, 10, 4741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wamwiri, F.N.; Alam, U.; Thande, P.C.; Aksoy, E.; Ngure, R.M.; Aksoy, S.; Ouma, J.O.; Murilla, G.A. Wolbachia, Sodalis and trypanosome co-infections in natural populations of Glossina austeni and Glossina pallidipes. Parasites Vectors 2013, 6, 232. [Google Scholar] [CrossRef] [Green Version]
- Kubi, C.; Billiouw, M.; Van den Bossche, P. Age prevalence of trypanosomal infections in female Glossina morsitans morsitans (Diptera: Glossinidae) on the plateau area of eastern Zambia. Onderstepoort J. Vet. Res. 2007, 74, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Vale, G.A.; Hargrove, J.W.; Solano, P.; Courtin, F.; Rayaisse, J.B.; Lehane, M.J.; Esterhuizen, J.; Tirados, I.; Torr, S.J. Explaining the host-finding behavior of blood-sucking insects: Computerized simulation of the effects of habitat geometry on tsetse fly movement. PLoS Negl. Trop. Dis. 2014, 8, e2901. [Google Scholar] [CrossRef] [PubMed]
- Mwamuye, M.M.; Kariuki, E.; Omondi, D.; Kabii, J.; Odongo, D.; Masiga, D.; Villinger, J. Novel Rickettsia and emergent tick-borne pathogens: A molecular survey of ticks and tick-borne pathogens in Shimba Hills National Reserve, Kenya. Ticks tick-borne Dis. 2017, 8, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Jagero, G.O. Molecular Evidence of Arthropod-Borne Pathogens in Wildlife and Livestock in Shimba Hills National Reserve (SHNR), Kwale County, Kenya. Master’s Dissertation, The University of Edinburgh, Edinburgh, UK, 2015. [Google Scholar]
- Knickerbocker, T.J.; Waithaka, J. People and elephants in the Shimba Hills, Kenya. Conserv. Biol. Ser.-Camb. 2005, 9, 224. [Google Scholar]
- Challier, A.; Laveissiere, C. A new trap for catching Glossina: Description and field trials. Cah. ORSTOM Ser. Entomol. Med et Parasitol. 1973, 11, 251–262. [Google Scholar]
- Pollock, J.N. Training Manual for Tsetse Control Personnel; Food and Agriculture Organization of the United Nations: Rome, Italy, 1982. [Google Scholar]
- Jackson, C.H. An artificially isolated generation of tsetse flies (Diptera). Bull. Entomol. Res. 1946, 37, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Omondi, D.; Masiga, D.K.; Ajamma, Y.U.; Fielding, B.C.; Njoroge, L.; Villinger, J. Unraveling host-vector-arbovirus interactions by two-gene high resolution melting mosquito bloodmeal analysis in a Kenyan wildlife-livestock interface. PLoS ONE 2015, 10, e0134375. [Google Scholar] [CrossRef] [Green Version]
- Boakye, D.A.; Tang, J.; Truc, P.; Merriweather, A.; Unnasch, T.R. Identification of bloodmeals in haematophagous Diptera by cytochrome B heteroduplex analysis. Med. Vet. Entomol. 1999, 13, 282–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Njiru, Z.K.; Constantine, C.C.; Guya, S.; Crowther, J.; Kiragu, J.M.; Thompson, R.C.; Dávila, A.M. The use of ITS1 rDNA PCR in detecting pathogenic African trypanosomes. Parasitol. Res. 2005, 95, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Dunn, P.K.; Smyth, G.K. Models for Proportions: Binomial GLMs. In Generalized Linear Models with Examples in R; Springer: New York, NY, USA, 2018; pp. 333–369. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S.; Hothorn, M.T. Package ‘multcomp’. In Simultaneous Inference in General Parametric Models; Project for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Team RC. R: A Language and Environment for Statistical Computing; Vienna, Austria. 2013. Available online: https://www.R-project.org/ (accessed on 3 November 2021).
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite package: Analysing ecological networks. Interaction 2008, 8, 8–11. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Tsetse Flies Screened | % Feeding Rate | 95% CI | |
|---|---|---|---|
| Fly sex ‡ | |||
| Female | 458 | 54.80 a | 50.23–59.38 |
| Male | 238 | 40.76 b | 34.47–47.04 |
| Fly species ‡ | |||
| G. austeni | 40 | 62.50 b | 46.82–78.18 |
| G. brevipalpis | 130 | 32.31 a | 24.16–40.45 |
| G. pallidipes | 526 | 53.42 b | 49.15–57.70 |
| Fly age † | |||
| Juvenile | 186 | 51.61 a | 44.36–58.86 |
| Old | 155 | 55.48 a | 47.57–63.40 |
| Young | 355 | 46.76 a | 41.55–51.98 |
| Landscape † | |||
| Cultivated field | 144 | 47.92 a | 39.66–56.17 |
| Forest | 55 | 50.91 a | 37.27–64.55 |
| Fruit-Orchard | 110 | 53.64 a | 44.17–63.10 |
| Grassland | 161 | 50.93 a | 43.13–58.74 |
| Peridomicilliary | 11 | 54.55 a | 19.46–89.63 |
| Shrubs | 215 | 48.37 a | 41.64–55.11 |
| Distance from the SHNR † | |||
| <1000 m | 614 | 50.65 a | 46.69–54.62 |
| 1000 to 1999 m | 61 | 44.26 a | 31.44–57.09 |
| >2000 m | 21 | 47.62 a | 24.32–70.91 |
| Number of Tsetse Flies Screened | % Feeding Rate | 95% CI | |
|---|---|---|---|
| Cluster † | |||
| Katangini | 25 | 44.00 a | 23.09–64.91 |
| Kidongo | 12 | 50.00 a | 16.82–83.18 |
| Kinangondogo | 17 | 47.06 a | 20.61–73.51 |
| Kipambane | 15 | 53.33 a | 24.74–81.93 |
| Kizibe | 134 | 47.76 a | 39.19–56.33 |
| Mangawani | 36 | 52.78 a | 35.65–69.91 |
| Mawia | 36 | 44.44 a | 27.39–61.50 |
| Mkanda | 8 | 12.50 a | −17.06–42.06 |
| Mlafyeni | 160 | 55.00 a | 47.21–62.79 |
| Msulwa A | 27 | 66.67 a | 47.66–85.67 |
| Msulwa Tangini | 2 | 100.00 a | 100.00–100.00 |
| Msulwa Viriko | 6 | 33.33 a | −20.86–87.53 |
| Pengo | 196 | 46.43 a | 39.38–53.47 |
| Zunguluka | 22 | 63.64 a | 41.81–85.47 |
| Sample ID (GenBank Accession No.) | Block | Latitude | Longitude | Fly Species | Fly Sex | Sequence Length (bp) | Closest Match on GenBank (Location) | Aimal Host Species | Sequence Identity (%) |
|---|---|---|---|---|---|---|---|---|---|
| GP370 (MZ816958) | Mlafyeni | −4.17453 | 39.39222 | G. pallidipes | F | 667 | DQ409327 (Africa) | Phacochoerus africanus | 99.55 |
| GP536 (MZ816959) | Mlafyeni | −4.20606 | 39.40222 | G. pallidipes | F | 595 | MN124266 (Kenya) | Phacochoerus africanus | 100.00 |
| GP411 (MZ816967) | Pengo | −4.20742 | 39.37234 | G. pallidipes | M | 607 | MN124266 (Kenya) | Potamochoerus porcus | 99.34 |
| GB412 (MZ816968) | Pengo | −4.25076 | 39.36938 | G. brevipalpis | F | 607 | MN124266 (Kenya) | Potamochoerus porcus | 99.01 |
| GP425 (MZ816969) | Pengo | −4.25076 | 39.36938 | G. pallidipes | F | 607 | MN124266 (Kenya) | Potamochoerus porcus | 99.38 |
| GB762 (MZ816966) | Pengo | −4.22782 | 39.37926 | G. brevipalpis | F | 607 | MN124266 (Kenya) | Potamochoerus porcus | 99.34 |
| GP362 (MZ816960) | Mlafyeni | −4.25085 | 39.36904 | G. pallidipes | F | 652 | MN124245 (Kenya) | Bos taurus | 99.85 |
| GP89 (MZ816962) | Mangawani | −4.3584 | 39.27996 | G. pallidipes | F | 652 | MT576844 (China) | Bos taurus | 100.00 |
| GP888 (MZ816961) | Mangawani | −4.3584 | 39.27996 | G. pallidipes | F | 652 | MT576844 (China) | Bos taurus | 100.00 |
| GB349 (MZ816970) | kinangondogo | −4.33653 | 39.34352 | G. brevipalpis | F | 396 | MN124271 (Kenya) | Loxodonta africana | 98.99 |
| GA379 (MZ816964) | katangini | −4.33402 | 39.35677 | G. austeni | F | 638 | JN645581 (Gabon) | Neotragus moschatus | 99.84 |
| GB545 (MZ816965) | Kizibe | −4.27812 | 39.31002 | G. brevipalpis | F | 538 | MF437212 (UAE) | Homo sapiens | 100.00 |
| GP665 (MZ816963) | Pengo | −4.28013 | 39.35485 | G. pallidipes | F | 662 | MN124246 (Kenya) | Capra hircus | 100.00 |
| GP344 (MZ816971) | Katangini | −4.31766 | 39.36762 | G. pallidipes | F | 470 | MN124256 (Kenya) | Syncerus caffer | 98.94 |
| G. austeni (n = 40) † | G. brevipalpis (n = 130) † | G. pallidipes (n = 526) ‡ | |||||||
|---|---|---|---|---|---|---|---|---|---|
| No | % | 95% CI | No | % | 95% CI | No | % | 95% CI | |
| Baboon | 4 | 10 a | 0.28–19.72 | 4 | 3.08 a | 0.07–6.09 | 54 | 10.27 c | 7.66–12.87 |
| Buffalo | 2 | 5 a | −2.06–12.06 | 0 | 0 | NA | 6 | 1.14 a | 0.23–2.05 |
| Bushbuck | 0 | 0 | NA | 0 | 0 | NA | 4 | 0.76 a | 0.02–1.51 |
| Cattle | 7 | 17.5 a | 5.19–21.81 | 14 | 10.77 a | 5.37–16.17 | 58 | 11.03 c | 8.34–13.71 |
| Donkey | 1 | 2.5 a | −2.56–7.56 | 0 | 0 | NA | 2 | 0.38 a | −0.15–0.91 |
| Elephant | 0 | 0 | NA | 3 | 2.31 a | −0.31–4.92 | 2 | 0.38 a | −0.15–0.91 |
| Goat | 0 | 0 | NA | 1 | 0.77 a | −0.75–2.29 | 3 | 0.57 a | −0.08–1.22 |
| Human | 3 | 7.5 a | −1.03–16.03 | 6 | 4.62 a | 0.96–8.27 | 17 | 3.23 ab | 1.72–4.75 |
| Impala | 0 | 0 | NA | 0 | 0 | NA | 3 | 0.57 a | −0.08–1.22 |
| Red Riverhog | 1 | 2.5 a | −2.56–7.56 | 4 | 3.08 a | 0.07–6.09 | 8 | 1.52 a | 0.47–2.57 |
| Sheep | 0 | 0 | NA | 5 | 6.65 a | 0.50–7.20 | 35 | 6.65 bc | 4.52–8.79 |
| Suni | 4 | 10 a | 0.28–19.72 | 0 | 0 | NA | 0 | 0 | NA |
| Warthog | 6 | 15 a | 3.43–26.57 | 7 | 5.38 a | 1.45–9.32 | 108 | 20.53 d | 17.07–23.10 |
| Host Family | No. of Tsetse Flies | Feeding Rate (%) | 95% CI |
|---|---|---|---|
| Bovidae | 142 | 20.40 a | 17.40–23.40 |
| Suidae | 134 | 19.25 a | 16.32–22.19 |
| Cercopithecidae | 62 | 8.91 b | 6.79–11.03 |
| Hominidae | 26 | 3.74 c | 2.32–5.15 |
| Elephantidae | 5 | 0.72 d | 0.09–1.35 |
| Equidae | 3 | 0.43 d | −0.06–0.92 |
| Host Species | |||
| Warthog | 121 | 17.39 f | 14.56–20.21 |
| Cattle | 79 | 11.35 ef | 8.99–13.71 |
| Baboon | 62 | 8.91 de | 6.79–11.03 |
| Sheep | 40 | 5.75 cd | 4.01–7.48 |
| Human | 26 | 3.74 bc | 2.32–5.15 |
| Red River Hog | 13 | 1.87 ab | 0.86–2.88 |
| Buffalo | 8 | 1.15 ab | 0.36–1.94 |
| Elephant | 5 | 0.72 a | 0.09–1.35 |
| Bushbuck | 4 | 0.57 a | 0.01–1.14 |
| Goat | 4 | 0.57 a | 0.01–1.14 |
| Suni | 4 | 0.57 a | 0.01–1.14 |
| Donkey | 3 | 0.43 a | −0.06–0.92 |
| Impala | 3 | 0.43 a | −0.06–0.92 |
| Bloodmeal-Positive Tsetse Flies | T. vivax | T. simiae Tsavo | T. simiae | ||||
|---|---|---|---|---|---|---|---|
| % (95% CI) | p-Value | % (95% CI) | p-Value | % (95% CI) | p-Value | ||
| Bovidae | 142 | 2.82 (0.06–5.57) | 0.746 | 1.41 (−0.55–3.37) | 0.844 | 0.70 (−0.69–2.10) | 0.982 |
| Suidae ‡ | 134 | 5.22 (1.41–9.04) | 0.027 | 1.49 (−0.59–3.57) | 0.479 | 1.49 (−0.59–3.57) | 0.259 |
| Cercopithecidae | 62 | 1.61 (−1.61–4.84) | 0.660 | 1.61 (−1.61–4.84) | 0.886 | NA | - |
| Hominidae | 26 | 3.85 (−4.08–11.77) | 0.640 | 3.85 (−4.08–11.77) | 0.093 | NA | - |
| Elephantidae | 5 | NA | - | NA | - | NA | - |
| Equidae | 3 | NA | - | NA | - | NA | - |
| Warthog ‡ | 121 | 5.79 (1.57–10.00) | 0.014 | 0.83 (−0.81–2.46) | 0.865 | 1.65 (−0.65–3.96) | 0.205 |
| Cattle | 79 | 1.27 (−1.25–3.79) | 0.482 | 2.53 (−1.01–6.07) | 0.313 | 1.27 (−1.25–3.79) | 0.548 |
| Baboon | 62 | 1.61 (−1.61–4.84) | 0.660 | 1.61 (−1.61–4.84) | 0.886 | NA | - |
| Sheep | 40 | 5.00 (−2.06–12.06) | 0.294 | NA | - | NA | - |
| Human | 26 | 3.85 (−4.08–11.77) | 0.640 | 3.85 (−4.08–11.77) | 0.093 | NA | - |
| Red River Hog | 13 | NA | - | 7.69 (−9.07–9.07) | 0.084 | NA | - |
| Buffalo | 8 | 12.50 (−17.06–42.06) | 0.103 | NA | - | NA | - |
| Elephant | 5 | NA | - | NA | - | NA | - |
| Bushbuck | 4 | NA | - | NA | - | NA | - |
| Goat | 4 | NA | - | NA | - | NA | - |
| Suni | 4 | NA | - | NA | - | NA | - |
| Donkey | 3 | NA | - | NA | - | NA | - |
| Impala | 3 | NA | - | NA | - | NA | - |
| Bloodmeal-Positive Tsetse Flies | T. godfreyi | T. congolense⁋ | T. brucei sl | ||||
| % (95% CI) | p- Value | % (95% CI) | p- Value | % (95% CI) | p- Value | ||
| Bovidae | 142 | 0.70 (−0.69–2.10) | 0.689 | 4.93 (−1.33–8.53) | 0.345 | NA | - |
| Suidae ‡ | 134 | 1.49 (−0.59–3.57) | 0.535 | 6.72 (−2.42–11.01) | 0.060 | 2.24 (−0.30–4.78) | 0.043 |
| Cercopithecidae | 62 | NA | - | NA | - | NA | - |
| Hominidae | 26 | NA | - | NA | - | NA | - |
| Elephantidae | 5 | NA | - | NA | - | NA | - |
| Equidae | 3 | NA | - | NA | - | NA | - |
| Warthog ‡ | 121 | 0.83 (−0.81–2.46) | 0.828 | 7.44 (2.70–12.18) | 0.033 | 2.48 (−0.33–5.29) | 0.031 |
| Cattle | 79 | 1.27 (−1.25–3.79) | 0.806 | 3.80 (−0.51–8.11) | 0.994 | NA | - |
| Baboon | 62 | NA | - | NA | - | NA | - |
| Sheep | 40 | NA | - | 2.50 (−2.56–7.56) | 0.785 | NA | - |
| Human | 26 | NA | - | NA | - | NA | - |
| Red River Hog ‡ | 13 | 7.69 (−9.07–9.07) | 0.046 | NA | - | NA | - |
| Buffalo | 8 | NA | - | NA | - | NA | - |
| Elephant | 5 | NA | - | NA | - | NA | - |
| Bushbuck | 4 | NA | - | NA | - | NA | - |
| Goat | 4 | NA | - | NA | - | NA | - |
| Suni ‡ | 4 | NA | - | 75.00 (4.56–154.56) | 0.0004 | NA | - |
| Donkey | 3 | NA | - | NA | - | NA | - |
| Impala | 3 | NA | - | NA | - | NA | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebhodaghe, F.I.; Okal, M.N.; Kalayou, S.; Bastos, A.D.S.; Masiga, D.K. Tsetse Bloodmeal Analyses Incriminate the Common Warthog Phacochoerus africanus as an Important Cryptic Host of Animal Trypanosomes in Smallholder Cattle Farming Communities in Shimba Hills, Kenya. Pathogens 2021, 10, 1501. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111501
Ebhodaghe FI, Okal MN, Kalayou S, Bastos ADS, Masiga DK. Tsetse Bloodmeal Analyses Incriminate the Common Warthog Phacochoerus africanus as an Important Cryptic Host of Animal Trypanosomes in Smallholder Cattle Farming Communities in Shimba Hills, Kenya. Pathogens. 2021; 10(11):1501. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111501
Chicago/Turabian StyleEbhodaghe, Faith I., Michael N. Okal, Shewit Kalayou, Armanda D. S. Bastos, and Daniel K. Masiga. 2021. "Tsetse Bloodmeal Analyses Incriminate the Common Warthog Phacochoerus africanus as an Important Cryptic Host of Animal Trypanosomes in Smallholder Cattle Farming Communities in Shimba Hills, Kenya" Pathogens 10, no. 11: 1501. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111501