
Emergence of Methicillin-Resistant Staphylococcus aureus ST239/241 SCCmec-III Mercury in Eastern Algeria

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Clinical Isolates
2.2. Antibiotic Susceptibility
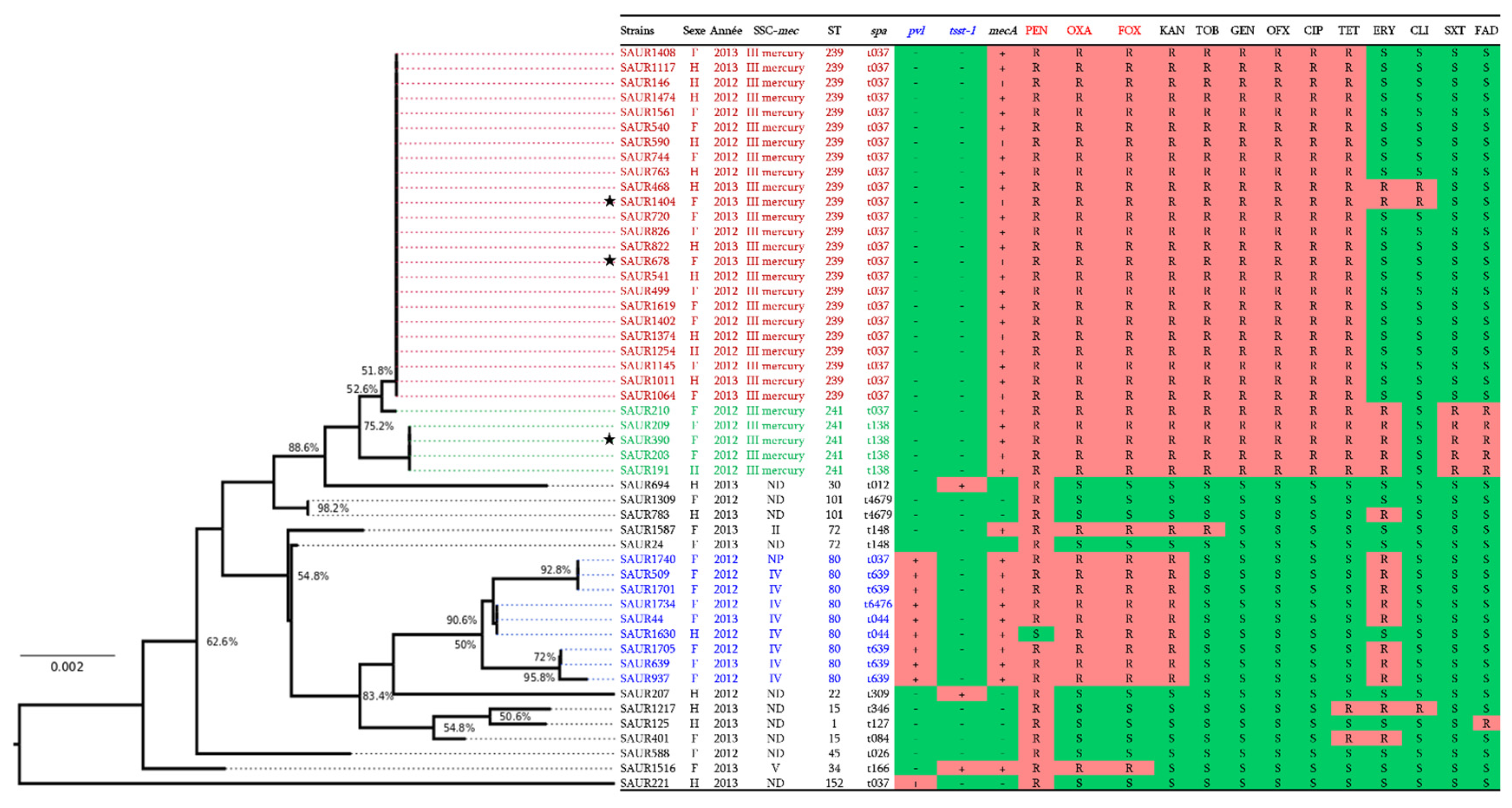
2.3. Molecular Characterisation
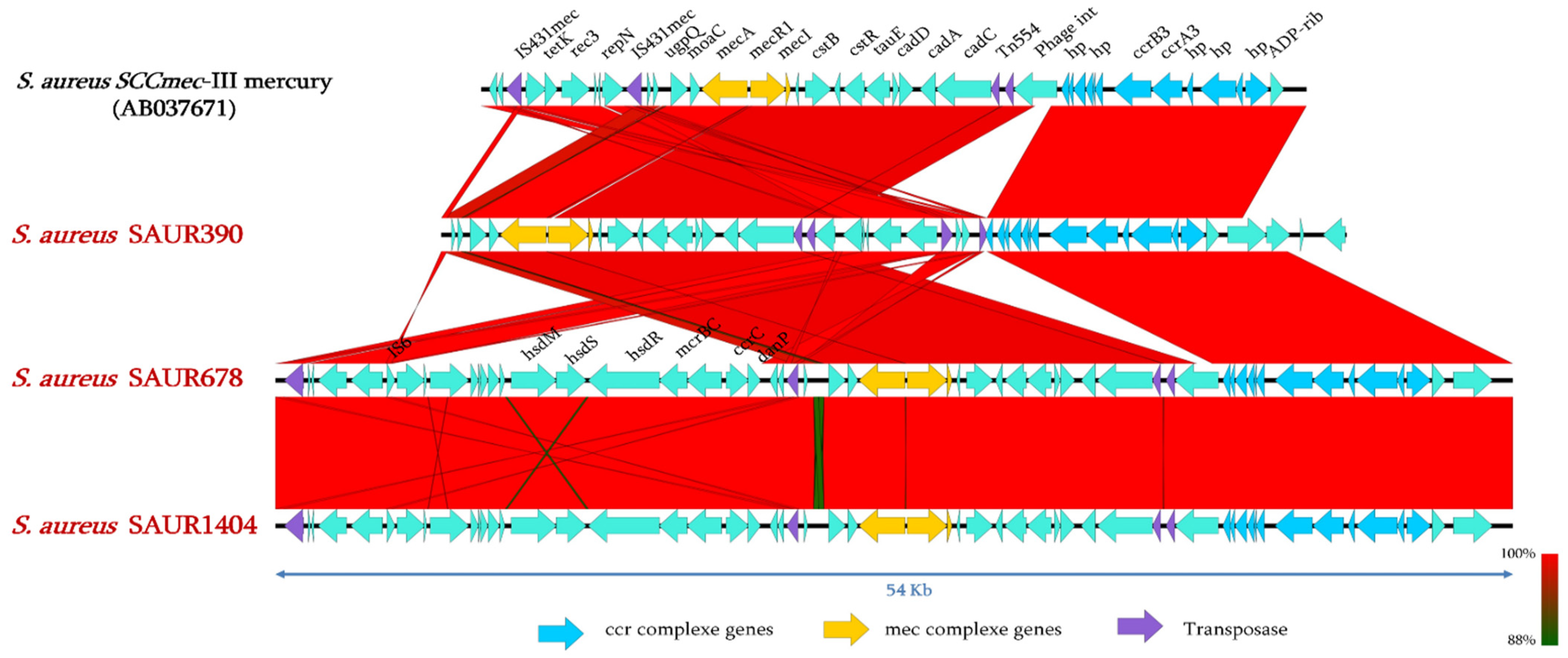
2.4. Genomic Analysis of SCCmec Cassettes from Sequenced Genomes
2.5. Resistance Genes Analysis of HA-MRSA SCCmec-III Mercury
2.6. Virulence Genes in HA-MRSA SCCmec-III Mercury
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Isolates and Identification
5.2. Antimicrobial Susceptibility Testing
5.3. Molecular Characterisation
5.4. Whole Genome Sequencing (WGS)
5.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Thomer, L.; Schneewind, O.; Missiakas, D. Pathogenesis of Staphylococcus aureus Bloodstream Infections. Annu. Rev. Pathol. Mech. Dis. 2016, 11, 343–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, A.M.S. What is behind the epidemiological difference between community-acquired and health-care associated methicillin-resistant Staphylococcus aureus? Virulence 2017, 8, 640–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, M.T.G.; Hsu, L.; Kurt, K.; Weinert, L.A.; Mather, A.E.; Harris, S.R.; Strommenger, B.; Layer, F.; Witte, W.; De Lencastre, H.; et al. A genomic portrait of the emergence, evolution, and global spread of methicillin-resistant Staphylococcus aureus pandemic. Genome Res. 2013, 23, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agaba, P.; Tumukunde, J.; Tindimwebwa, J.V.B.; Kwizera, A. Nosocomial bacterial infections and their antimicrobial susceptibility patterns among patients in Ugandan intensive care units: A cross sectional study. BMC Res. Notes 2017, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, B.J.; Tomasz, A. Low-Affinity Penicillin-Binding Protein Associated with β-Lactam Resistance in Staphylococcus aureus. J. Bacteriol. 1984, 158, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Katayama, Y.; Ito, T.; Hiramatsu, K. A New Class of Genetic Element, Staphylococcus Cassette Chromosome mec, Encodes Methicillin Resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2000, 44, 1549–1555. [Google Scholar] [CrossRef] [Green Version]
- Dupieux, C.; Bouchiat, C.; Larsen, A.R.; Pichon, B.; Holmes, M.; Teale, C.; Edwards, G.; Hill, R.; Decousser, J.-W.; Trouillet-Assant, S.; et al. Detection of mecC -Positive Staphylococcus aureus: What To Expect from Immunological Tests Targeting PBP2a? J. Clin. Microbiol. 2017, 55, 1961–1963. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Hiramatsu, K.; Oliveira, D.C.; De Lencastre, H.; Zhang, K.; Westh, H.; O’Brien, F.; Giffard, P.M.; Coleman, D.; Tenover, F.C.; et al. Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 53, 4961–4967. [Google Scholar]
- Wu, Z.; Li, F.; Liu, D.; Xue, H.; Zhao, X. Novel type XII staphylococcal cassette chromosome mec harbouring a new cassette chromosome recombinase, CcrC2. Antimicrob. Agents Chemother. 2015, 59, 7597–7601. [Google Scholar] [CrossRef] [Green Version]
- Baig, S.; Johannesen, T.B.; Overballe-petersen, S.; Larsen, J.; Larsen, A.R.; Stegger, M. Novel SCCmec type XIII (9A) identified in an ST152 methicillin-resistant Staphylococcus aureus. Infect. Genet. Evol. 2018, 61, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Urushibara, N.; Aung, M.S.; Kawaguchiya, M.; Kobayashi, N. Novel staphylococcal cassette chromosome mec (SCCmec) type XIV (5A) and a truncated SCCmec element in SCC composite islands carrying speG in ST5 MRSA in Japan. J. Antimicrob. Chemtherapy 2019, 75, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Deurenberg, R.H.; Vink, C.; Kalenic, S.; Friedrich, A.W.; Bruggeman, C.A.; Stobberingh, E.E. The molecular evolution of methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2007, 13, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; McClure, J.A.; Conly, J.M. Enhanced multiplex PCR assay for typing of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. Mol. Cell. Probes. 2012, 26, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, D.; Peters, B.M.; Li, L.; Li, B.; Xu, Z.; Shirliff, M.E. Staphylococcal chromosomal cassettes mec (SCCmec): A mobile genetic element in methicillin-resistant Staphylococcus aureus. Microb. Pathog. 2016, 101, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Rolo, J.; Worning, P.; Nielsen, J.B.; Bowden, R.; Bouchami, O.; Damborg, P.; Guardabassi, L.; Perreten, V.; Tomasz, A.; Westh, H.; et al. Evolutionary Origin of the Staphylococcal Cassette Chromosomemec (SCCmec). Antimicrob. Agents Chemother. 2017, 61, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Z.; Peng, C.; Lu, X. Diversity of SCCmec Elements in Methicillin-Resistant Coagulase-Negative Staphylococci Clinical Isolates. PLoS ONE 2011, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tulinski, P.; Fluit, A.C.; Wagenaar, J.A.; Mevius, D.; van de Vijver, L.; Duim, B. Methicillin-Resistant Coagulase-Negative Staphylococci on Pig Farms as a Reservoir of Heterogeneous Staphylococcal Cassette Chromosome mec Elements. Appl. Environ. Microbiol. 2012, 78, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Chen, L.; Wang, J.; Wang, W.; Chen, D.; Li, L.; Li, B.; Deng, Y.; Xu, Z. Current methodologies on genotyping for nosocomial pathogen methicillin-resistant Staphylococcus aureus (MRSA). Microb. Pathog. 2017, 107, 17–28. [Google Scholar] [CrossRef]
- Szabo, J. Molecular Methods in Epidemiology of Methicillin Resistant Staphylococcus aureus (MRSA): Advantages, Disadvantages of Different Techniques. J. Med. Microbiol. Diagn. 2014, 3, 1. [Google Scholar] [CrossRef]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. TheLancet. Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zheng, Y.; Mediavilla, J.R.; Chen, L.; Kreiswirth, B.N.; Song, Y.; Yang, R.; Du, H. Hospital Dissemination of tst-1-Positive Clonal Complex 5 (CC5) Methicillin-Resistant Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2017, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Abdulgader, S.M.; Shittu, A.O.; Nicol, M.P.; Kaba, M. Molecular epidemiology of Methicillin-resistant Staphylococcus aureus in Africa: A systematic review. Front. Microbiol. 2015, 6, 1–21. [Google Scholar] [CrossRef]
- Basset, P.; Amhis, W.; Blanc, D.S. Changing molecular epidemiology of methicillin-resistant Staphylococcus aureus in an algerian hospital. J. Infect. Dev. Ctries. 2015, 9, 206–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchenane, Z.; Agabou, A.; Smati, F.; Rolain, J.M.; Raoult, D. Staphylococcal cassette chromosome mec characterisation of methicillin-resistant Staphylococcus aureus strains isolated at the military hospital of Constantine/Algeria. Pathol.-Biol. 2013, 61, 280–281. [Google Scholar] [CrossRef] [PubMed]
- Rebiahi, S.A.; Abdelouahid, D.E.; Rahmoun, M.; Abdelali, S.; Azzaoui, H. Emergence of vancomycin-resistant Staphylococcus aureus identified in the Tlemcen university hospital (North-West Algeria). Médecine Mal. Infect. 2011, 41, 646–651. [Google Scholar] [CrossRef]
- Alioua, M.A.; Labid, A.; Amoura, K.; Bertine, M.; Gacemi-Kirane, D.; Dekhil, M. Emergence of the European ST80 clone of community-associated methicillin-resistant Staphylococcus aureus as a cause of healthcare-associated infections in Eastern Algeria. Médecine Mal. Infect. 2014, 44, 180–183. [Google Scholar] [CrossRef]
- Ramdani-Bouguessa, N.; Bes, M.; Meugnier, H.; Forey, F.; Reverdy, M.-E.; Lina, G.; Vandenesch, F.; Tazir, M.; Etienne, J. Detection of methicillin-resistant Staphylococcus aureus strains resistant to multiple antibiotics and carrying the Panton-Valentine leukocidin genes in an Algiers hospital. Antimicrob. Agents Chemother. 2006, 50, 1083–1085. [Google Scholar] [CrossRef] [Green Version]
- Bouchiat, C.; Curtis, S.; Spiliopoulou, I.; Bes, M.; Cocuzza, C.; Codita, I.; Dupieux, C.; Giormezis, N.; Kearns, A.; Laurent, F.; et al. MRSA infections among patients in the emergency department: A European multicentre study. J. Antimicrob. Chemother. 2017, 72, 372–375. [Google Scholar] [CrossRef]
- Simor, A.E.; Pelude, L.; Golding, G.; Fernandes, R.; Bryce, E.; Frenette, C.; Gravel, D.; Katz, K.; McGeer, A.; Mulvey, M.R.; et al. Determinants of Outcome in Hospitalized Patients With Methicillin-Resistant Staphylococcus aureus Bloodstream Infection: Results From National Surveillance in Canada, 2008–2012. Infect. Control. Hosp. Epidemiol. 2016, 37, 390–397. [Google Scholar] [CrossRef]
- Antri, K.; Rouzic, N.; Boubekri, I.; Dauwalder, O.; Beloufa, A.; Ziane, H.; Djennane, F.; Neggazi, M.; Benhabyles, B.; Bes, M.; et al. Forte prevalence des infections communautaires et nosocomiales àStaphylococus aureus résistant à la méticilline et portant le gène de la leucocidine de Panton-Valentine dans l’Algérois. Pathol.-Biol. 2010, 58, e15–e20. [Google Scholar] [CrossRef] [PubMed]
- Djoudi, F.; Bonura, C.; Benallaoua, S.; Touati, A.; Touati, D.; Aleo, A.; Cala, C.; Fasciana, T.; Mammina, C. Panton-Valentine leukocidin positive sequence type 80 methicillin-resistant Staphylococcus aureus carrying a staphylococcal cassette chromosome mec type IVc is dominant in neonates and children in an Algiers hospital. New Microbiol. 2013, 36, 49–56. [Google Scholar] [PubMed]
- Djahmi, N.; Messad, N.; Nedjai, S.; Moussaoui, A.; Mazouz, D.; Richard, J.L.; Sotto, A.; Lavigne, J.P. Molecular epidemiology of Staphylococcus aureus strains isolated from inpatients with infected diabetic foot ulcers in an Algerian University Hospital. Clin. Microbiol. Infect. 2013, 19, E398–E404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djoudi, F.; Benallaoua, S.; Aleo, A.; Touati, A.; Challal, M.; Bonura, C.; Mammania, C. Descriptive Epidemiology of Nasal Carriage of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus among Patients Admitted to Two Healthcare Facilities in Algeria. Microbial. Drug Resist. 2015, 21, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.; Yu, F.; Zhang, W.; Li, X.; Wang, H. Molecular Epidemiology and Antibiotic Resistance Profiles of Methicillin-Resistant Staphylococcus aureus Strains in a Tertiary Hospital in China. Front. Microbiol. 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Valsesia, G.; Rossi, M.; Bertschy, S.; Pfyffer, G.E. Emergence of SCCmec type IV and SCCmec type V methicillin-resistant Staphylococcus aureus containing the Panton-Valentine leukocidin genes in a large academic teaching hospital in Central Switzerland: External invaders or persisting circulators? J. Clin. Microbiol. 2010, 48, 720–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sit, P.S.; Teh, C.S.J.; Idris, N.; Sam, I.-C.; Syed Omar, S.F.; Sulaiman, H.; Thong, K.L.; Kamarulzaman, A.; Ponnampalavanar, S. Prevalence of methicillin-resistant Staphylococcus aureus (MRSA) infection and the molecular characteristics of MRSA bacteraemia over a two-year period in a tertiary teaching hospital in Malaysia. BMC Infect. Dis. 2017, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, B.; Rao, C.; Udo, E.E.; Gadepalli, R.; Vishnubhatla, S.; Kapil, A. Dissemination of methicillin-resistant Staphylococcus aureus SCCmec type IV and SCCmec type V epidemic clones in a tertiary hospital: Challenge to infection control. Epidemiol. Infect. 2015, 143, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Tavares, A.; Faria, N.A.; de Lencastre, H.; Miragaia, M. Population structure of methicillin-susceptible Staphylococcus aureus (MSSA) in Portugal over a 19-year period (1992–2011). Eur. J. Clin. Microbiol. Infect. Dis. 2014, 15, 423–432. [Google Scholar] [CrossRef]
- Benito, D.; Lozano, C.; Jiménez, E.; Albújar, M.; Gómez, A.; Rodríguez, J.M.; Torres, C. Characterisation of Staphylococcus aureus strains isolated from faeces of healthy neonates and potential mother-to-infant microbial transmission through breastfeeding. FEMS Microbiol. Ecol. 2015, 91, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, 1–24. [Google Scholar]
- Huang, V.; Cheung, C.M.; Kaatz, G.W.; Rybak, M.J. Evaluation of dalbavancin, tigecycline, minocycline, tetracycline, teicoplanin and vancomycin against community-associated and multidrug-resistant hospital-associated meticillin-resistant Staphylococcus aureus. Int. J. Antimicrob. Agents 2010, 35, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S. Molecular Typing of ST239-MRSA-III From Diverse Geographic Locations and the Evolution of the SCC Mec III Element During Its Intercontinental Spread. Front. Microbiol. 2018, 9, 1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.R.; Feil, E.J.; Holden, M.T.G.; Quail, M.A.; Emma, K.; Chantratita, N.; Gardete, S.; Tavares, A.; Day, N.; Jodi, A.; et al. Evolution of MRSA During Hospital Transmission and Intercontinental Spread. Science 2010, 327, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Soliman, M.S.; Soliman, N.S.; El-Manakhly, A.R.; ElBanna, S.A.; Aziz, R.K.; El-Kholy, A.A. Genomic Characterisation of Methicillin-Resistant Staphylococcus aureus (MRSA) by High-Throughput Sequencing in a Tertiary Care Hospital. Genes 2020, 11, 1219. [Google Scholar] [CrossRef] [PubMed]
- Elshimy, R.; Khattab, R.A.; Zedan, H.; Hosny, A.E.S.; Elmorsy, T.H. Study on prevalence and genetic discrimination of methicillin-resistant Staphylococcus aureus (MRSA) in Egyptian hospitals. Afr. J. Microbiol. Res. 2018, 12, 629–646. [Google Scholar]
- Gordon, N.C.; Price, J.R.; Cole, K.; Everitt, R.; Morgan, M.; Finney, J.; Kearns, A.M.; Pichon, B.; Young, B.; Wilson, D.J.; et al. Prediction of Staphylococcus aureus Antimicrobial Resistance By Whole-Genome Sequencing. J. Clin. Microbiol. 2014, 52, 1182–1191. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, F.; Katouli, M.; Pourshafie, M.R. Characteristics of hospital- and community-acquired meticillin-resistant Staphylococcus aureus in Tehran, Iran. J. Med. Microbiol. 2014, 63, 796–804. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, M.; Baserisalehi, M.; Bahador, N.; Taherikalani, M. Antibiotic Resistance Patterns and Virulence Determinants of Different SCCmec and Pulsotypes of Staphylococcus aureus Isolated from a Major Hospital in Ilam, Iran. Open Microbiol. J. 2017, 11, 211–223. [Google Scholar] [CrossRef]
- Bendary, M.; HEGAZY, W.; Sweed, A.; Samir, M.; Mosbah, R.; Ghaith, D.; Abdelaziz, A.; Ramadan, H.; Alshareef, W.; Abd El Hamid, M. Clonal Diversity and Epidemiological Characteristics of ST239-MRSA Strains. Authorea 2020. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial Resistance and Virulence: A Successful or Deleterious Association in the Bacterial World? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranovich, T.; Zaraket, H.; Shabana, I.I.; Nevzorova, V.; Turcutyuicov, V.; Suzuki, H. Molecular characterisation and susceptibility of methicillin-resistant and methicillin-susceptible Staphylococcus aureus isolates from hospitals and the community in Vladivostok, Russia. Clin. Microbiol. Infect. 2009, 80, 575–582. [Google Scholar]
- Howden, B.P.; Seemann, T.; Harrison, P.F.; Mcevoy, C.R.; Stanton, J.L.; Rand, C.J.; Mason, C.W.; Jensen, S.O.; Firth, N.; Davies, J.K.; et al. Complete Genome Sequence of Staphylococcus aureus Strain JKD6008, an ST239 Clone of Methicillin-Resistant Staphylococcus aureus with Intermediate-Level Vancomycin Resistance. J. Bacteriol. 2010, 192, 5848–5849. [Google Scholar] [CrossRef] [Green Version]
- Japoni, A.; Jamalidoust, M.; Farshad, S.; Ziyaeyan, M. Characterisation of SCCmec Types and Antibacterial Susceptibility Patterns of Methicillin-Resistant Staphylococcus aureus in Southern Iran. Jpn. J. Infect. Dis. 2011, 64, 28–33. [Google Scholar]
- Boyle-Vavra, S.; Li, X.; Alam, M.T.; Read, T.D.; Sieth, J.; Cywes-Bentley, C.; Dobbins, G.; David, M.Z.; Kumar, N.; Eells, S.J.; et al. USA300 and USA500 Clonal Lineages of Staphylococcus aureus Do Not Produce a Capsular Polysaccharide Due to Conserved Mutations in the cap5 Locus. MBio 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Spaan, A.N.; van Strijp, J.A.G.; Torres, V.J. Leukocidins: Staphylococcal bi-component pore-forming toxins find their receptors. Nat. Rev. Microbiol. 2017, 15, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.C.; Lina, G.; Bes, M.; Vandenesch, F.; Piémont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2018, 359, 753–759. [Google Scholar] [CrossRef]
- Melles, D.C.; Taylor, K.l.; Fattom, A.I.; Belkum, A. Serotyping of Dutch Staphylococcus aureus strains from carriage and infection. FEMS Immunol. Med. Microbiol. 2008, 52, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Li, D.Z.; Chen, Y.S.; Yang, J.P.; Zhang, W.; Hu, C.P.; Li, J.S.; Mu, L.; Hu, Y.H.; Geng, R.; Hu, K.; et al. Preliminary molecular epidemiology of the Staphylococcus aureus in lower respiratory tract infections: A multicenter study in China. Chin. Med. J. 2011, 124, 687–692. [Google Scholar] [CrossRef]
- Alibayov, B.; Zdeňková, K.; Purkrtová, S.; Demnerová, K.; Karpíšková, R. Detection of some phenotypic and genotypic characteristics of Staphylococcus aureus isolated from food items in the Czech Republic. Ann. Microbiol. 2014, 64, 1587–1596. [Google Scholar] [CrossRef]
- Li, H.; Llera, A.S.; Malchiodi, E.L.; Mariuzza, R.A. The structural basis of t cell activation by superantigens. Annu. Rev. Immunol. 1999, 17, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, T. Staphylococcal Superantigens: Pyrogenic Toxins Induce Toxic Shock. Toxins 2019, 1, 1–19. [Google Scholar]
- Seng, P.; Rolain, J.-M.; Fournier, P.E.; La Scola, B.; Drancourt, M.; Raoult, D. MALDI-TOF-mass spectrometry applications in clinical microbiology. Future Microbiol. 2010, 5, 1733–1754. [Google Scholar] [CrossRef] [PubMed]
- Cockerill, F. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2012; Volume 32, pp. 1–184. ISBN 1562387855. [Google Scholar]
- Sociéte Francaise de Microbiologie Comite de L’antibiogramme de la Societe Francaise de Microbiologie Recommandations. Comité de L’Antibio. 2013. Available online: https://www.sfm-microbiologie.org/wp-content/uploads/2020/07/CASFM_2013.pdf (accessed on 6 October 2021).
- Sharp, S.E.; Warren, J.A.; Thomson, R.B. Cefoxitin disk diffusion screen for confirmation of oxacillin-resistant Staphylococcus aureus isolates and utility in the clinical laboratory. Diagn. Microbiol. Infect. Dis. 2005, 51, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Felten, A.; Grandry, B.; Lagrange, P.H.; Casin, I. Evaluation of three techniques for detection of low-level methicillin-resistant Staphylococcus aureus (MRSA): A disk diffusion method with Cefoxitin and Moxalactam, the Vitek 2 system, and the MRSA-screen latex agglutination test. J. Clin. Microbiol. 2002, 40, 2766–2771. [Google Scholar] [CrossRef] [Green Version]
- Brennan, G.I.; Abbott, Y.; Burns, A.; Leonard, F.; McManus, B.A.; O’Connell, B.; Coleman, D.C.; Shore, A.C. The emergence and spread of multiple livestock-associated clonal complex 398 methicillin-resistant and methicillin-susceptible Staphylococcus aureus strains among animals and humans in the Republic of Ireland, 2010–2014. PLoS ONE 2016, 11, 1–11. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.J.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus Sequence Typing for Characterisation of Methicillin-Resistant and Methicillin-Susceptible Clones of Staphylococcus aureus Multilocus Sequence Typing for Characterisation of Methicillin- Resistant and Methicillin-Susceptible Clones of Staphyloco. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| All Strains | HA-MRSA | HA-MSSA | |
|---|---|---|---|
| n = 50 | n = 40 | n = 10 | |
| Samples | |||
| Bloodstream | 22 (44%) | 17 (42.5%) | 5 (50%) |
| Tracheal incubation | 20 (40%) | 15 (37.5%) | 5 (50%) |
| Central catheter | 2 (4%) | 2 (5%) | - |
| Chest drain | 2 (4%) | 2 (5%) | - |
| Urine | 2 (4%) | 2 (5%) | - |
| Pleural | 1 (2%) | 1 (2.5%) | - |
| Urinary catheter | 1 (2%) | 1 (2.5%) | - |
| Sex | |||
| Men | 19 (38%) | 13 (32.5%) | 6 (60%) |
| Women | 27 (54%) | 23 (57.5%) | 4 (40%) |
| Age (years) | |||
| 0–10 | 4 (8%) | 4 (10%) | - |
| 11–20 | 4 (8%) | 4 (10%) | - |
| 21–30 | 7(14%) | 4 (10%) | 3 (30%) |
| 31–40 | 5 (10%) | 3 (7.5%) | 2 (20%) |
| 41–50 | 8 (16%) | 8 (20%) | - |
| 51–60 | 3 (6%) | 3 (7.5%) | - |
| 61–70 | 6 (12%) | 4 (10%) | 2 (20%) |
| 71–80 | 7 (14%) | 5 (12.5%) | 2 (20%) |
| 81–90 | 2 (4%) | 1 (2.5%) | 1 (10%) |
| Antibiotics | HA-MRSA | HA-MSSA | Total | p Value |
|---|---|---|---|---|
| n = 40 | n = 10 | n = 50 | ||
| Penicillin (10 UI) | 39 (97.5%) | 10 (100%) | 49 (98%) | n.s |
| Oxacillin (1 µg) | 40 (100%) | - | 40 (80%) | <0.001 |
| Cefoxitin (30 µg) | 40 (100%) | - | 40 (80%) | <0.001 |
| Kanamycin (30 µg) | 39 (97.5%) | - | 39 (78%) | <0.001 |
| Tobramycin (10 µg) | 30 (75%) | - | 30 (60%) | <0.001 |
| Gentamicin (15 µg) | 29 (72.5%) | - | 29 (58%) | <0.001 |
| Erythromycin (15 µg) | 14 (35%) | 3 (30%) | 17 (36%) | n.s |
| Clindamycin (2 µg) | 2 (5%) | 1 (10%) | 3 (6%) | n.s |
| Ciprofloxacin (5 µg) | 29 (72.5%) | - | 29 (58%) | <0.001 |
| Ofloxacin (5 µg) | 29 (72.5%) | - | 29 (58%) | <0.001 |
| Trimethoprim-sulfamethoxazole (1.25 µg, 23.75 µg) | 5 (12.5%) | - | 5 (10%) | n.s |
| Fusidic acid (30 µg) | 5 (12.5%) | 1 (10%) | 6 (12%) | n.s |
| Tetracycline (15 µg) | 29 (72.5%) | 2 (20%) | 31 (62%) | 0.007 |
| Pristinamycin (15 µg) | - | - | - | - |
| Tigecycline (15 µg) | - | - | - | - |
| Rifampicin (5 µg) | - | - | - | - |
| Linezolid (10 µg) | - | - | - | - |
| Mupirocin (5 µg) | - | - | - | - |
| Vancomycin (30 µg) | - | - | - | - |
| Teicoplanin (30 µg) | - | - | - | - |
| MDR | 29 (72.5%) | - | 29 (58%) | <0.001 |
| Antibiotic families | SAUR390 | SAUR678 | SAUR1404 |
|---|---|---|---|
| Beta-lactams | mecA; blaZ; mecR1; mecI | mecA; blaZ; mecR1; mecI | mecA; blaZ; mecR1; mecI |
| Aminoglycosides | ant(9)-Ia; aph-Stph; aadC; aph(3′)-IIIa; aac(6′)-Ie-aph(2′’)-Ia | aph-Stph; aad(6); aac(6′)-Ie-aph(2′’)-Ia; aph(3′’)-Ib; aph(6)-Id; aph(3′)-Ia; aph(3′)-IIIa | aph-Stph; aph(6)-Id; aac(6′)-Ie-aph(2′’)-Ia |
| Streptothricins | Sat-4 | Sat-4 | / |
| Tetracyclines | tet(38); tet(K); tetM | tet(38); tet(K); tetM | tet(38); tet(K); tetM |
| Sulfamids | dfrC; dfrE; DfrG | dfrC; dfrE | dfrC; dfrE |
| Macrolids | erm(C); erm(A) | / | erm(C); erm(A) |
| Phenicols | dha1 | dha1 | dha1 |
| Quinolones | norA; norB; mgrA; arlR; arlS | norA; norB; mgrA; arlR; arlS | norA; norB; mgrA; arlR; arlS |
| Fosfomycin | fosB3 | fosB3 | fosB3 |
| Efflux pumps | Three copies of sav1866; mepA; mepR | Three copies of sav1866; mepA; mepR | Three copies of sav1866; mepA; mepR |
| Virulence Factors | SAUR390 | SAUR678 | SAUR1404 |
|---|---|---|---|
| Immunomodulators | spa; isdA; isdB; isdC; isdD; isdE; isdF; isdG; sbi; esaA; esaB; essA; essB; essC; esxA | spa; isdA;isdB;isdC; isdD; isdE; isdF; isdG; sbi; esaA; esaB;essA; essB; essC; esxA | spa; isdA;isdB;isdC; isdD; isdE; isdF; isdG; sbi; esaA; esaB;essA; essB; essC; esxA |
| Adhesins | Lap; sdrC; sdrD; sdrE; clfA; clfB; fnbA; fnbB; psaA; vwbp; ebp; map; icaA; icaB; icaC; icaD; icaR; cna | Lap; sdrC; sdrD; sdrE; clfA; clfB; fnbA; fnbB; psaA; vwbp; ebp; map; icaA; icaB; icaC; icaD; icaR; cna | Lap; sdrC; sdrD; sdrE; clfA; clfB; fnbA; fnbB; psaA; vwbp; ebp; map; icaA; icaB; icaC; icaD; icaR; cna |
| Exoenzyme | adsA; coa; geh; lip; srtB; katA; sspA; sspB; sspC; clpC; clpE; clpP; aur; ureB; ureG; hysA; cpsA; cpsJ; htpB; cap8A; cap8B; cap8C; cap8D; cap8E; cap8F; cap8G; cap8H; cap8I; cap8J; cap8K; cap8L; cap8M; cap8N; cap8O; cap8P | adsA; coa; geh; lip; srtB; kat; sspA; sspB; sspC; clpC; clpE; clpP; aur; ureB; ureG; hysA; cpsA; cpsJ; htpB; sak; scn; cap8D; cap8E; cap8F; cap8G; cap8H; cap8I; cap8J; cap8K; cap8L; cap8M; cap8N; cap8O; cap8P | adsA; coa; geh; lip; srtB; kat; sspA; sspB; sspC; clpC; clpE; clpP; aur; ureB; ureG; hysA; cpsA; cpsJ; htpB; sak; scn; cap8D; cap8E; cap8F; cap8G; cap8H; cap8I; cap8J; cap8K; cap8L; cap8M; cap8N; cap8O; cap8P |
| Toxins | hlb; hld; hlgA; hlgB; hlgC; lukF-PV | hlb; hld; hlgA; hlgB; hlgC; lukF-PV; sea; selk; selq | hlb; hld; hlgA; hlgB; hlgC; lukF-PV: sea; selk; selq |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aouati, H.; Hadjadj, L.; Aouati, F.; Agabou, A.; Ben Khedher, M.; Bousseboua, H.; Bentchouala, C.; Rolain, J.-M.; Diene, S.M. Emergence of Methicillin-Resistant Staphylococcus aureus ST239/241 SCCmec-III Mercury in Eastern Algeria. Pathogens 2021, 10, 1503. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111503
Aouati H, Hadjadj L, Aouati F, Agabou A, Ben Khedher M, Bousseboua H, Bentchouala C, Rolain J-M, Diene SM. Emergence of Methicillin-Resistant Staphylococcus aureus ST239/241 SCCmec-III Mercury in Eastern Algeria. Pathogens. 2021; 10(11):1503. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111503
Chicago/Turabian StyleAouati, Hanane, Linda Hadjadj, Farida Aouati, Amir Agabou, Mariem Ben Khedher, Hacène Bousseboua, Chafia Bentchouala, Jean-Marc Rolain, and Seydina M. Diene. 2021. "Emergence of Methicillin-Resistant Staphylococcus aureus ST239/241 SCCmec-III Mercury in Eastern Algeria" Pathogens 10, no. 11: 1503. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111503