
The Interplay between Salmonella and Intestinal Innate Immune Cells in Chickens
by
 , , , and
, , , and
Adil Ijaz
1 ,
,
Edwin J. A. Veldhuizen
1,
Femke Broere
1,
Victor P. M. G. Rutten
1,2 and
Christine A. Jansen
3,* 1
Division of Infectious Diseases and Immunology, Department of Biomolecular Health Sciences, Faculty of Veterinary Medicine, Utrecht University, Yalelaan 1, 3584 CL Utrecht, The Netherlands
2
Department of Veterinary Tropical Diseases, Faculty of Veterinary Science, University of Pretoria, Onderstepoort, Pretoria 0110, South Africa
3
Cell Biology and Immunology Group, Department of Animal Sciences, Wageningen University & Research, De Elst 1, 6708 PB Wageningen, The Netherlands
*
Author to whom correspondence should be addressed.
Pathogens 2021, 10(11), 1512; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111512
Submission received: 30 September 2021
/
Revised: 15 November 2021
/
Accepted: 15 November 2021
/
Published: 19 November 2021
(This article belongs to the Special Issue The Interaction between Pathogens and the Chicken Innate Immune System)
Abstract
:Salmonellosis is a common infection in poultry, which results in huge economic losses in the poultry industry. At the same time, Salmonella infections are a threat to public health, since contaminated poultry products can lead to zoonotic infections. Antibiotics as feed additives have proven to be an effective prophylactic option to control Salmonella infections, but due to resistance issues in humans and animals, the use of antimicrobials in food animals has been banned in Europe. Hence, there is an urgent need to look for alternative strategies that can protect poultry against Salmonella infections. One such alternative could be to strengthen the innate immune system in young chickens in order to prevent early life infections. This can be achieved by administration of immune modulating molecules that target innate immune cells, for example via feed, or by in-ovo applications. We aimed to review the innate immune system in the chicken intestine; the main site of Salmonella entrance, and its responsiveness to Salmonella infection. Identifying the most important players in the innate immune response in the intestine is a first step in designing targeted approaches for immune modulation.
1. Introduction
Salmonellosis is an intestinal bacterial infection of poultry. The disease may be caused by various serovars of Salmonella and infection can lead to severe symptoms such as gastroenteritis, septicemia, and typhoid fever and can cause mortality in young chickens. This makes Salmonella infection a major concern for the poultry industry [1]. The severity of infection depends on the serovar, the history of exposure, the age and the genotype of chickens. Some of the S. enterica serovars, such as S. ser. Gallinarium and S. ser. Pullorum, are host specific, but the majority of the Salmonella strains can infect multiple host species. Colonization of S. ser. Enteriditis and S. ser. Typhimurium in the ileum and cecum of the chicken causes enteric salmonellosis in young chickens of 2–3 days of age. S. ser. Pullorum and S. ser. Gallinarum infect chickens of all ages and cause a chronic typhoid-like disease resulting in significant mortality. S. enterica serovars induce systemic infection via lymphoid tissues such as Peyer’s patches and cecal tonsils. Salmonella infected phagocytes enter the lymphatics and bloodstream and disseminate bacteria to spleen, liver, bone marrow, and ovaries, thereby causing a second round of infection. In chickens, S. enterica serovars may reside in the spleen for months without showing obvious clinical signs and lead to infection of the reproductive tract [2]. Especially, S. ser. Pullorum colonizes the reproductive tract of the chickens and infects progeny through vertical transmission [3]. These examples clearly show the large diversity of infections and related disease caused by different Salmonella serovars, and thereby the complexity to describe and define a ‘Salmonella infection’.
Salmonella contaminated poultry meat products and eggs from Salmonella-infected chickens can lead to food-borne zoonosis in humans as poultry products are highly consumed food commodities of humans [4,5,6]. In 2010, more than 0.5 billion S. ser. Enteriditis contaminated eggs were recalled because of a nationwide outbreak of human food-borne salmonellosis in the US (https://www.cdc.gov/Salmonella/2010/shell-eggs-12-2-10.html, accessed on 26 August 2021). A recent report mentioned 87,923 confirmed cases of human salmonellosis in 2019, the 2nd highest human zoonosis in Europe [6]. In the EU, the overall economic burden due to human salmonellosis has been estimated to be over three billion euros annually (https://www.efsa.europa.eu/en/topics/topic/Salmonella, accessed on 26 August 2021). The worldwide incidence of salmonellosis is 1.3 billion, due to which 3 million people die annually [7]. Hence, controlling Salmonella infections in chickens is highly important to diminish the incidence of zoonotic Salmonella infections in humans.
To control Salmonella infections, the EU has adopted an integrated approach mainly focused on food safety by ensuring pathogen elimination from animal food products, including poultry meat and eggs (https://eur-lex.europa.eu/eli/reg/2012/1190/oj, accessed on 26 August 2021). The EU member states have initiated a Salmonella control program in poultry aiming to implement strict biosecurity, using vaccination against Salmonella in poultry, and destruction of infected eggs and birds in case of outbreaks in flocks (https://eur-lex.europa.eu/eli/reg/2006/1177/oj, accessed on 30 August 2021).
In poultry production, vaccination against Salmonella is nowadays common practice to provide protection to young birds. However, multiple studies have shown that vaccination doesn’t guarantee full protection and prevention of bacterial shedding in older birds, especially in layers. Moreover, Salmonella vaccines are not multivalent, so whereas those vaccines help to limit the spread of certain Salmonella serovars, they allow other harmful Salmonella strains to grow. Therefore, even after vaccination, a flock can be infected by Salmonella and transmit pathogen both horizontally and vertically.
Antibiotics as feed additives have proven to be an effective prophylactic option to control intestinal Salmonella infections [8], but due to limited effectiveness of antibiotics against Salmonellae and resistance issues in humans and animals, the use of antimicrobials has been banned in food animals in Europe [9]. Hence, there is an urgent need for alternative strategies to protect poultry from Salmonella infection. One alternative is to strengthen the innate immune system in young chickens to prevent early life intestinal infections. The focus on young chickens is specifically important because their adaptive immune system is not fully developed to generate an effective Salmonella specific immune response and therefore host defense mostly relies on innate immunity. Strengthening of the innate immune system in young chickens can, for example, be achieved by addition of immune modulating molecules to feed, or even by in-ovo application of immunostimulants.
Innate immunity that plays an important role against Salmonella invasion, may be affected by several factors. In chickens, the differentiation and proliferation of innate immune cells start during embryonic life and continue during the early days post-hatch, in addition, it has been shown that the competence of innate immune cells improves with increasing age [10]. Furthermore, the intestinal microbiota do contribute to innate immunity as it provides competition to the invading Salmonella serovars in the gut. Moreover, feed helps to attain diversity of intestinal microbiota that contribute to resistance to Salmonella colonization in the gut [11]. Also, the genetic background of the birds, as well as housing environment can affect innate immunity in chickens [12,13].
The objective of this review is to discuss the chicken intestinal innate immune system in order to determine how innate immune fitness can be increased and used to prevent infectious diseases like salmonellosis in chickens and hence to reduce the risk of zoonotic infections in humans.
2. The Chicken Intestine
The intestine is a complex organ that primarily functions to digest food into small particles and to transport it into the body to produce energy [14]. The chicken gut is home of a diverse microbial community, most of which are Gram-positive and facultative anaerobes [15]. The chicken gastrointestinal (GI) system is comparable to the digestive tract of mammals, but with several avian specific features like the crop, gizzard, ceca, and cloaca [14] (Figure 1a). The avian GI tract starts from the beak connected to the stomach by the esophagus. The crop, an out-pocket of the esophagus, used to store and process food for digestion [16], does so, due to its own microbiota, which includes Furmicutes, Bacteriodetes, Proteobacteria, and Actinobacteria [17]. The chicken stomach consists of the proventriculus, which is a glandular part, and the ventriculus or gizzard, which acts as a mechanical stomach because of its strong muscles and grinding action. The gizzard also acts as a microbial barrier because of its low pH [18]. The gizzard is followed by the small intestine, which consists of three sections, the duodenum, jejunum, and ileum, respectively. The duodenum plays an important role in food digestion. Bicarbonate and digestive enzymes from the pancreatic ducts, and bile from the liver help in food digestion. The absorption of nutrients takes place in the ileum. The most predominant bacterial species present in all three sections of the small intestine is Lactobacillus [18,19]. The Meckel’s diverticulum is a small structure present on the small intestine at the transition between jejunum and ileum. The ceca are two blind pouches attached to the small intestine, and involved in the fermentation of nutrients [20]. The cecal microbiota has greater richness and diversity as compared to the small intestinal microbiota [18,21,22]. It consists of Enterococcus, coliforms, Lactobacillus, and yeast [23,24]. The large intestine in chickens, the colon, terminates at the cloaca, which is a common cavity for the digestive and urogenital tract [14].
In the early days post-hatch, the intestine of the chickens is relatively sterile since the gut microbiota is still developing, which will result in 1010 bacteria/gram of digesta in the ileum and cecum, within 3 days [25]. During the early days, the chicken gut provides relatively low competitive exclusion for pathogen colonization and growth. Thus, it is a favorable site for Salmonella colonization [26,27]. External factors such as food and water can be a source of transferring Salmonella [28,29]. At this age exposure to Salmonella is detrimental to the growth of the natural gut microbiota, by causing reduction in microbial diversity, and hence allowing the growth of opportunistic pathogens in the gut [30]. The gut and its associated lymphoid tissue (GALT) provide protection against harmful effects of intestinal pathogens like Salmonella.
3. The Immune System of the Chicken Intestine
The chicken intestinal epithelium is a monolayer consisting of various cell types. Intercellular junctional complexes, tight junctions, present between the intestinal epithelial cells maintain the gut barrier integrity. Tight junctions hold the intestinal epithelial cells together to prevent the entry of enteric pathogens, for example, Salmonella, to the lamina propria, and thus maintain gut homeostasis [31]. In addition to the gut microbiota, which inhibits pathogen colonization; intestinal epithelial cells through their secretions also inhibit pathogen entry in the gut. For example, the outer surface of the intestinal epithelial cells is covered with a mucus layer, which provides a diffusive barrier between the intestinal microbiota and epithelial cells [32]. Mucus is secreted by intestinal goblet cells and is composed of 95% water and only 5% of the mucus contents contain salts, lipids, gel-forming mucin glycoproteins, defensins, and lysozyme [33,34,35,36,37,38]. Muc2, Muc5a, Muc5b, and Muc6 are gel forming mucins in chicken intestine [39]. The intestinal epithelium is the interface between the gut microbiota and the body [40]. The most abundant cell type of the intestinal epithelium, the enterocytes, represent more than 80% of the intestinal lining [41]. The primary function of the enterocytes is to digest and absorb food. Besides enterocytes, the monolayer contains the goblet cells, which secrete mucus and are abundantly present in the ileum (26%) and jejunum (23%) [42]. A third cell type present is the enteroendocrine cells, whose main function is to secrete enzymes to promote digestion of food components. These cells also produce hormones that are released in the bloodstream instead of the intestinal lumen. Enteroendocrine cells are sparsely present in the intestinal epithelium representing approximately 1% of the cells [43]. Paneth cells, specialized intestinal epithelial cells located at the base of the intestinal crypts of most of the vertebrates, are best known for their secretion of antimicrobial compounds such as phospholipase, lysozyme and α-defensins [44]. Three types of Paneth cells have been identified so far in poultry. These include, c-type -conventional or chicken type- mostly present in small intestine, and oviduct [45], g-type -goose type-, which are prevalent in the lungs and bone marrow [46], and g2-type which are expressed in the liver, kidney, and small intestine [45]. In chickens, however, the presence of the Paneth cells is controversial [45].
In addition to the gut epithelial cells, also immune cells are present along the length of the intestine as the GALT, part of the mucosa-associated lymphoid tissues (MALT) [47]. In contrast to mammals, chickens do not have encapsulated lymph nodes, instead mucosa-associated and diffused lymphoid tissues are present. The GALT is exposed to more antigens than any other tissue of the body and is present at strategically important locations along the length of the intestine. The chicken GALT includes Peyer’s patches (PP), cecal tonsils (CT), Meckel’s diverticulum, intraepithelial lymphocytes (IELs), and lamina propria lymphocytes (LPL), which are located beneath the intestinal epithelium as shown in Figure 1b [48,49,50].
The cellular architecture of the chicken GALT primarily consists of IELs (γδ T cells, αβ T cells, and NK cells), and phagocytic cells such as dendritic cells (DCs), and monocytes/macrophages. These cells either directly kill the invading pathogens or secret pro-inflammatory cytokines to attract other immune cells to mount the host immune response. DCs and macrophages also act as antigen presenting cells in the secondary lymphoid organs such as PP [51]. The immune cells of the GALT work in collaboration to initiate host defense responses during bacterial invasion. The innate immune cells are armed with sensors, specific for pathogens, so-called pattern recognition receptors (PRRs) [52]. The PRRs recognize pathogen-associated molecular patterns (PAMPs) and damage-associated molecular pattern (DAMPs). The PRRs can be cell-associated (Toll-like receptors, scavenger receptors, C-type lectins, intracellular receptors) or soluble components (pentraxins, LPS binding proteins, collectins). The PRRs are present at the cell surface in the plasma membrane, and in the cytoplasm and recognize most of the enteric pathogens. Probably the most important, or at least the best described PRRs are Toll-Like Receptors (TLR). These sensory receptors recognize pathogens through ligands present on bacteria (and/or other pathogens), which include, for example, flagellin (TLR5), LPS (TLR4), and CpG-DNA (TLR21) [53,54,55,56,57].
4. Innate Immune Cells in the Chicken Intestine
In the intestine, the IELs are embedded between the enterocytes of the intestinal epithelium. In adult chickens, the IEL population is comprised of NK cells, γδ, and αβ T cells [58,59]. Among the IEL population, NK and γδ T cells are innate immune cells and formulate the first line of host immune defense against invading pathogens [60]. An overview of the different mediators produced by intestinal innate immune cells as well as the intestinal epithelial cells, and the functions performed by them is shown in Table 1.
4.1. Natural Killer (NK) Cells of the Intestinal Immune System
NK cells are involved in the innate defense, they are scavenger cells and their primary function is to recognize the virally infected, transformed, and neoplastic host cells and kill them [61,62]. NK cells originate in the bone marrow and migrate towards blood and organs like the spleen, lungs, and intestine [63]. In the chicken intestine NK cells are embedded between the enterocytes as intraepithelial lymphocytes along the length of the gut. In intestinal intraepithelial lymphocytes NK cells comprise even higher numbers than CD8+ T cells and γδ T cells [59,64] suggesting an important role in the intestinal innate immune system [65].
Chicken NK cells are described as a population of cells that lack surface expression of CD3 or Ig [64] that is able to kill the NK susceptible cell line (LSCC-RP9) [59]. Based on these characteristics, many studies have reported the presence of NK cells in various tissues like spleen, lungs and intestine of chickens [66,67,68,69,70]. NK cells are also present in large numbers in the embryonic spleen and duodenum of chickens [59,67,70,71] indicating that they are indeed important in early life innate defense.
The induction of NK cell activation is mediated by an array of activating and inhibitory receptors which are present on their surface [72]. Also, in the chicken genome NK cell receptors have been reported to be present. The leucocyte receptor complex on chromosome 31 encodes for chicken Ig-like receptors (CHIR) [73]. Chromosome 1 harbors the NK gene complex which encodes NKG2/CD94 and syntenic to the mammalian NK complex region [73,74,75].
Upon crosslinking, activation receptors on NK cells signal via immunoreceptor tyrosine-based activation motifs (ITAMs) which initiates downstream signaling, while inhibitory receptors signal via immunoreceptor tyrosine-based inhibitory motifs (ITIMs). The delicate balance between activating and inhibitory signals received by NK cell receptors activates NK cells [76,77,78].
Once activated, NK cells release lytic granules that contain perforin and granzyme to lyse target cells [79,80]. The degranulation of NK cells upon activation can be detected using the CD107 assay [67], since enhanced surface expression of CD107a, the lysosomal-associated membrane protein-1 (LAMP-1), is associated with NK cell activation [66]. In chickens, enhanced CD107 surface expression was reported on CD3 negative cells that express surface markers such as 28-4, 20E5, CD11b/c, and 7C1 suggesting that these are markers of cells with NK cell function.
NK cells that express 28-4 (the chicken IL-2Ra orthologue) are mostly present in the duodenum of layer chickens and have been shown to able to kill susceptible target cells (LSCC-RP9) [59,81]. In other organs such as spleen, blood, and lung of layer chickens NK cells expressing 28-4 are less dominant, here the major populations express 5C7 and 20E5 [59,67,81,82].
In the intestine, the majority of NK cells express 28-4, while lower expression of NK cell markers 20E5 and 5C7 was observed. The number of 28-4 expressing NK cells was higher at day 1 post-hatch compared to levels at embryonic day 14 and 18, and remained stable throughout the life. Conversely, the number of 20E5+ and 5C7+ cells was lower in 1 day-old chickens compared to embryonic day 14 and 18 and increased until day 21 post-hatch. Among intestinal NK cells, surface expression of CD107 was also higher in the 28-4+ subset as compared to intestinal 20E5+ cells suggesting that this population may play a role in the intestinal innate immune response [70].
4.2. Heterophils of the Intestinal Immune System
Heterophils are the avian analogues of mammalian neutrophils and immediately appear at the site of infection to eliminate invasive pathogens [85,86]. Heterophils use multiple strategies such as phagocytosis, degranulation, and oxidative burst to kill the pathogens. In chickens, heterophils are the predominant granulocytes in circulating blood and the gut [87,88,89,90,91,92]. Heterophils are comparatively higher in number in young chickens as compared to the older ones [93]. However, in young birds heterophils are functionally less active as indicated by decreased phagocytosis, degranulation, oxidative burst and, subsequently, killing of bacteria.
In the chicken intestine, pro-inflammatory cytokines produced as a result of microorganism invasion attract heterophils to the site of infection [94]. Upon contact with pathogens, heterophils are activated through the interaction of TLRs with bacterial ligands such as LPS, peptidoglycan, flagellin, and lipoteichoic acid [53,95]. This activation of heterophils results in a sequence of events including phagocytosis, oxidative burst, degranulation, and cytokine (IL1β, IL6) and chemokine (CXCLi2) production through the NF-κB pathway [53,56]. Heterophils also become activated via avian triggering receptor expressed on myeloid cells-1 (TREM-A1), which modulates TLR signaling and causes phagocytosis [96].
Phagocytosis is the process by which heterophils internalize the pathogens. Following internalization, microorganisms are entrapped in a vacuole called the phagosome, which then immediately fuses with cytoplasmic granules leading to killing of entrapped pathogens by release of anti-microbicidal peptides and proteolytic enzymes from intracellular granules [97].
Degranulation is another host defense strategy used by heterophils to kill pathogens. Upon microbial agonist stimulation, heterophils release their cytoplasmic granules at the site of infection into the external environment to kill pathogens. Another important event closely linked with extracellular degranulation is the production of heterophil extracellular traps (HET) to trap pathogens. The HET is similar to the neutrophil extracellular trap [98]. This process allows heterophils to entrap and kill bacteria extracellularly. The HET mechanism is initiated by the formation of thin extracellular fibers (5–17 nm) between pathogens and heterophils [99]. This meshwork of fibers acts as a death trap and prevent the dissemination of pathogens in the tissues. Moreover, HET also destroy pathogens by highly concentrated microbicidal substances. Finally, phagocytes also use oxidative burst to produce reactive oxygen species for microbial killing. However, unlike neutrophils, avian heterophils are mostly dependent on non-oxidative killing of pathogens as they don’t generate strong oxidative burst as compared to their mammalian analogues [100,101].
4.3. Dendritic Cells (DCs) of the Intestinal Immune System
DCs act as an immunological bridge between innate and adaptive immunity since these cells are the key player of antigen presentation. The intestinal DCs take up and process antigens, and present these to adaptive immune cells, which initiates a pathogen-specific immune response [102]. In mice DCs are present underneath the intestinal epithelium together with macrophages [103]. Monocytes, macrophage progenitors, and DCs express colony stimulating factor 1 receptor [104]. The intestinal DCs in human and mice can be differentiated from intestinal monocyte-derived macrophages using CD64 expression [105].
Avian DCs are less well characterized compared to human and mice DCs in which the ontogeny and heterogenous nature is extensively explored [106,107,108]. Avian DCs are defined by the expression of several markers such as CD83, CD11c, DEC205, and MHC II [109,110,111,112]. In chickens, DCs expressing CD83 and DEC205 markers are particularly abundant in the spleen, cecal tonsils, thymus, and bursa of fabricius [111]. In the chicken intestine, DCs have been described in the clusters of the GALT. Follicular DCs are located in the germinal center of the cecal tonsil, and pyloric tonsils [113,114,115]. Follicular and interdigitating DCs have also been described in the PP [116].
Immature avian DCs are equipped with a variety of TLRs to recognize invading pathogen [117]. DCs also express the CCR6 chemokine receptor. Binding of of CCR6 to this receptor induces infiltration of the DCs at the infection site to encounter antigens, due to which immature DCs transform into the mature phenotype that presents antigens to T cells. Stimulation of chicken bone marrow derived DCs with LPS results into maturation of chicken DCs, which parallels increased expression of co-stimulatory molecules CD40, CD83, CD86, and a decrease in phagocytic activity [110,118]. DCs interact with LPS of the Gram-negative bacteria through TLR4, which activates CD14 dependent endocytosis of TLR4 [119]. In addition to antigen presentation, DCs also produce cytokines such as IL1β, IL6, 1L10, IL12p35, and TNFα, and CXCL chemokines to attract other immune cells and to enhance inflammatory processes [120,121].
4.4. Macrophages of the Intestinal Immune System
Macrophages are a type of innate immune cells, which can be involved in the defense against bacterial infections [122]. Macrophages actively recognize, phagocytize, and kill microbes by producing microbicidal substances like nitric oxide, reactive oxygen species, proteolytic enzymes, and lysozyme [123,124,125,126,127]. Upon contact with a pathogen, macrophages become activated and transform into either M1 or M2 phenotype [128,129,130]. The polarization of the macrophages into M1 or M2 phenotypes depends upon the activation pathway and the type of cytokines they are exposed to. Exposure of macrophages to intracellular bacteria and T helper cell type 1 (Th1) cytokines (interferon and TNFα) leads to M1 polarization [131], while T helper cell type 2 (Th2) cytokines (IL4, and 13) transform macrophages into the M2 phenotype [132,133,134].
Whether this polarization also occurs in chicken macrophages is currently not clear. It has been shown, in vitro, that cultured avian macrophages can have a more M1 like phenotype as described by Peng et al. [135] for monocyte-derived macrophages, while an IL4 induced polarization towards a more M2 phenotype has also been described [136]. However, evidence for the presence of M1 and M2 macrophages in vivo is still missing.
Chicken macrophages recognize pathogens by their phagocytic receptors [137], that may be opsonic (complement receptors, Fc receptors) or non-opsonic (TLRs, mannose receptors) [138,139,140,141]. Chicken intestinal macrophages possess a range of TLRs, and hence respond accordingly to a variety of bacterial ligands such as LPS, CpG oligonucleotides, and flagellin [142,143,144,145].
Upon pathogen recognition, macrophages activate intracellular signals (mitogen activated protein kinase p38) orchestrate the innate immune response by production of cytokines (TNFα, IL1, IL10), chemokines, and nitric oxide (NO) which has antibacterial properties [146,147,148,149,150,151,152,153]. In a recent study it has been shown that HD11 cells, a macrophage-like cell line, when stimulated with inactivated poultry vaccines, showed an increase in Fc-receptor driven phagocytosis as well as NO production when activated with TLR agonists [154]. In another study, HD11 cells stimulated with inactivated Avibacterium paragallinarium, led to NO production and express pro-inflammatory cytokines TNFα, IL1β, and IL12p40, as well as chemokines CXCLi1 and CXCLi2 [155].
In the chicken intestine, macrophages play a vital role in antigen presentation along with DCs, as well as act as regulators and effectors of immunity. Macrophages are present in the intestinal lamina propria in chickens and reach the invasion site after IFNγ production [156]. It has been shown that IFNγ stimulation increases the antiviral and phagocytic activity of macrophages and also induces IL12 and IL18 production which drives Th1 adaptive response [157,158].
4.5. γδ T Cells of the Intestinal Immune System
γδ T cells are unconventional CD3+ T cells having a unique T cell receptor (TCR) that consists of a γ chain and a δ chain. The number of γδ T cells in blood varies between 0.5 to 10% in humans, dogs, mice and monkeys [159,160], while in chickens, cattle and pigs γδ T cells ranges from 20–50% of the total circulating T cells in blood [161,162,163,164]. Apart from blood, γδ T cells have also been reported in, for example, the intestine and spleen and their frequency depends on age, sex, and strain of chickens [165,166].
The majority of the avian γδ T cells is activated in a MHC unrestricted way unlike αβ T cells [167]. Although toll-like receptors (TLR3 and 4) are present on chicken γδ T cells along with the scavenger receptor superfamily, their role in TCR independent activation of γδ T cells in chickens is still unclear [166,168]. Recently, Karunakaran et al. showed the TCR dependent activation of human γδ T cells. After stimulating γδ T cells with phospho-antigens, molecules present inside the infected cells, γδ T cells get activated to kill the target cell [169]. Activated γδ T cells also produce IL17 and IFNγ cytokines to attract other innate immune cells [170].
The primary function of γδ T cells is to recognize stressed, transformed, and tumor cells, and their killing by perforins and granzymes [171,172]. The mode of action of γδ T cells in chickens is still unclear, but a recent study demonstrated that γδ T cells show killing of LSCC-RP9 cells [166].
γδ T cells are abundantly present in the chicken intestine. It has been shown that the percentage of γδ T cells among IEL population in the ileum of the chicken intestine remained the same during late embryonic days and early post-hatch days but started to increase from day 14 to day 21 post-hatch. As compared to other T cell subsets in the chicken intestine such as CD8αα and CD8αβ T cells, the presence of γδ T cells is lower in chickens younger than 7 days of age, but from day 14 onwards their number becomes similar to CD8αα and CD8αβ T cells [70].
5. Interactions between Salmonella and Innate Immune Cells in the Chicken Intestine
Salmonella serovars enter chickens via the oral route through infected feed, water, litter or through vertical transmission from infected hens, and colonize the distal part of the ileum and cecum. Intestinal epithelial and immune cells embedded in the gut epithelium provide the first protective barrier against Salmonella. In the lumen of the chicken gut, Salmonella serovars outcompete the gut microflora and make the first contact with intestinal epithelial cells [177].
As Salmonella are facultative intracellular bacteria, and thus can invade intestinal epithelial cells, after adherence through TLR5. The flagella and fimbriae present on the bacterial cell surface facilitate the adhesion of Salmonella serovars to intestinal epithelial cells [178,179]. Once attached, Salmonella serovars establish themselves into an intracellular niche using the protein complex T3SS-1 (type 3 secretion system-1). T3SS-1 together with its effector proteins acts as a molecular syringe, and thereby ensures the efficient invasion of Salmonella serovars in the host cells [180,181,182,183,184,185,186]. Several kinds of effector proteins such as SopA/B/D/E/E2, AvrA) secreted by the T3SS-1 machinery into the host cell cytoplasm, particularly SopE/E2 and SopB which activate Rho GTPase and transform inactive GDP to an active GDP bound to initiate translocation of NF-κB in the nucleus [187,188,189]. The activation of the NF-κB pathway induces the expression of pro-inflammatory cytokines such as IL18, IL1β and CXCL chemokines which attract innate immune cells such as heterophils, macrophages, and DCs to limit Salmonella invasion as shown in Figure 1c [94,174,175,176]. In chickens, the fate of the initial inflammatory response and consequently disease progression due to Salmonella serovars (systemic or gut restricted) depend on the presence or absence of a flagella, a TLR5 agonist, [190]. It is well demonstrated, both in vitro and in vivo, that flagellated Salmonella serovars like S. ser. Gallinarum and S. ser. Pullorum elicited lower or no inflammatory signal during epithelial invasion compared to flagellated Salmonella serovars [54,191]. Aflagellated Salmonella serovars cross the gut epithelial barrier using stealth strategy and, therefore, evade recognition by intestinal epithelial cells, which leads to systemic infection [191]. Moreover, mutations in the flagellin gene of flagellated Salmonella serovars such as S. ser. Typhimurium also cause rapid epithelial invasion of Salmonella serovars with low inflammation [54,191].
Following Salmonella invasion, avian heterophils reach the site of infection during the initial stages and attack the invading pathogen by producing toxic reactive oxygen species and microbicidal peptides [100,192,193,194]. An in vivo study demonstrated the influx of heterophils at the site of inflammation which resulted in resistance against Salmonella infection in chickens [143]. Heterophils also use extracellular traps to kill Salmonella serovars. Jana et al. [195] have shown that exposure of S. ser. Enteriditis to heterophils results in their formation within 15 min in vitro. Moreover, heterophils interact with invading Salmonella serovars through TLR5, which is specific for bacterial flagellin [196]. The TLR5-flagellin interaction activates the MyD88-dependent downstream signaling pathways in host cell [197,198], which activates MAPK and NF-κB and consequently leads to secretion of pro-inflammatory cytokines that causes the influx of resident and circulating immune cells at the infection site [199,200,201,202,203].
Macrophages also reach the site of infection in the initial stages of infection and recognize Salmonella serovars through their surface TLRs. The activated macrophages then produce NO, which is often used as a functional readout of macrophage activation, to kill Salmonella [204]. This inflammatory response is age dependent and is more pronounced in young chickens (up to 2 weeks of age) compared to adults [205]. It has been shown, in vitro, that HD11 macrophages induced significant NO production after 24 h of infection with chicken serum opsonized S. ser. Gallinarum [206].
The extent to which macrophages respond against Salmonella serovars is correlated with the susceptibility of chickens against the infection [207]. The phagocytic potential of macrophages against invading pathogens and their extent to present antigens differs a lot among various chicken breeds [152]. Sun et al. [208] have studied the phagocytic potential of monocyte-macrophages of the Silky and Starbro chicken lines in vitro and found that the degree of adherence of S. ser. Pullorum to silky monocyte-macrophages was 1.5 times higher than to those of Starbro chickens. Similarly, the phagocytic potential of the macrophages of Silky chickens was also greater than Starbro chickens following S. ser. Pullorum inoculation [208].
Macrophages also activate the NF-κB pathway which induces release of inflammatory factors such as the cytokines IL6, IL8, and TNFα to strengthen early inflammatory responsiveness and to promote immune defense [209,210]. Recently, it has been shown that stimulation of HD11 macrophages with Salmonella serovars induced the production of pro-inflammatory cytokines such as IL1β, IFNγ, and CXCLi chemokines, in-vitro, during initial stages of infection [206]. In the meantime, DCs present in the GALT, also reach at the invasion site, recognize the invading Salmonella serovar and present it to the adaptive immune cells to initiate the adaptive immune response [211].
Salmonella serovars can also use macrophages and dendritic cells as a vehicles for systemic dissemination of infection by using Salmonella pathogenicity island (SPI) T3SS-2 [212,213]. Upon successful crossing of the intestinal epithelial barrier, Salmonella serovars invade the underlying lymphoid tissues and subsequently get endocytosed by host macrophages [214]. Inside the macrophages, Salmonella serovars reside in membrane bounded vesicles known as Salmonella containing vesicles (SCV), where T3SS-2 with its effector proteins is rapidly expressed to promote survival and replication of Salmonella serovars inside the host macrophages, and prevent the lysis of invading bacteria by the host cell lysosomes [215]. It is believed that the Salmonella serovars are carried by the macrophages and DC via the lymphatic system to the liver and spleen, where they replicate and cause systemic infection, although there is no experimental evidence for that [216]. Macrophages not only kill the bacteria but at the same time also transmit both soluble and contact dependent signals to the resident cytotoxic intraepithelial lymphocytes (NK cells, γδ T cells, CD8+ αβ T cells).
NK cells embedded between the enterocytes recognize the infected intestinal epithelial cells and are activated directly upon recognition of these transformed cells. NK cells are also activated indirectly by the soluble and contact signals transmitted by macrophages [217]. Once activated NK cells release cytoplasmic granules containing perforins and granzymes to kill Salmonella infected cells. In addition to that, NK cells also release the cytokine IFNγ to attract other cells to initiate phagocytosis to kill Salmonella and Salmonella infected cells. NK cells are the important source of IFNγ production during initial days of Salmonella infection in chickens, and they play an important role in determining the fate of disease progression in chickens [218].
In newly hatched chickens, innate immunity is even more important because host defense against pathogens actually largely depends on innate immune cells [219]. NK cells play a pivotal role in Salmonella infection in early days of life. Meijerink et al. [220] described that immediately after entry of Salmonella in the chicken gut (1 day post-infection), the numbers of intraepithelial lymphocytes including NK cell subsets and CD8+ T cells significantly increase. It has also been demonstrated that, in chickens, degranulation of NK cells and IFNγ production, which are the functional readouts of NK cell activation, are also increased during the initial days of S. ser. Enteriditis infection [220]. This not only helps to kill Salmonella, but also attracts additional innate immune cells to avoid Salmonella invasion and systemic infection. S. ser. Enteriditis infection in 1 day old pathogen free chickens results in the production of chemokines, which leads to macrophages infiltration to the intestine within 24 h of infection [191]. Moreover, Salmonella infection in young chickens also gives rise to increased presence of γδ T cells, CD8+ cytotoxic T cells, and CD4+ helper T cells within one week at the site of infection, beside the killing of Salmonella [218,221,222,223,224]. γδ T cells recognize the Salmonella infected cells through cellular stress markers or phospho-antigens and cause the killing of infected host cells. Activated γδ T cells also produce immune activation mediators such as IFNγ, and IL17 cytokines to initiate Th1 response [221,222,225]. This shows that innate immune cells play an important role in young chickens in the host defense against Salmonella infection.
6. Future Prospects
Many studies underscore the importance of intestinal innate immune cells in the defense against Salmonella infections. Several pre- and probiotic compounds have been reported to increase the abundance of beneficial bacteria in the gut, expression of the tight junction proteins, and TLR expression by the intestinal innate immune cells [226,227,228,229,230]. Supplementation of certain carbohydrate derivatives such as glucomannan modulates the gut microbiota that inhibit the Salmonella colonization in the gut of broiler chickens [11]. Moreover, oligosaccharide compounds because of their structural resemblance with LPS, a TLR4 agonist, have the potential to activate innate immune cells [227]. Targeting the innate immune cells using oligosaccharide compounds, either in-ovo or post-hatch through feed formulations, could be an alternative options to strengthen immune mediated disease resistance in chickens. The recent development of novel in vitro chicken intestine models [231,232] can be used to study immune response of novel carbohydrate compounds against intestinal pathogens including Salmonella. This would help us to identify compounds that can enhance the immune fitness of innate immune cells, which can be used as alternatives to antibiotics. Apart from being economically beneficial for the poultry industry, it will also limit risks of AMR resistance issues in poultry, and incidence of Salmonella infections in humans.
7. Conclusions
In conclusion, the literature described shows that the innate immune system plays a pivotal role in resisting the spread of Salmonella in the initial stages of infection. This is especially relevant in newly hatched chickens, where the adaptive immune system is still not fully developed. Specifically targeting these innate immune cells using for example oligosaccharide compounds may be a novel strategy to strengthen the immune mediated resistance in young chickens.
Author Contributions
Conceptualization, A.I., C.A.J., E.J.A.V.; writing—original draft preparation, A.I.; writing—review and editing, A.I., C.A.J., E.J.A.V., F.B., V.P.M.G.R. All authors have read and agreed to the published version of the manuscript.
Funding
A.I. is the recipient of an international PhD fellowship from the Punjab Educational Endowment Fund, Punjab, Pakistan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Castanheira, S.; García-Del Portillo, F. Salmonella populations inside host cells. Front. Cell. Infect. Microbiol. 2017, 7, 432. [Google Scholar] [CrossRef] [Green Version]
- Shivaprasad, H. Fowl typhoid and pullorum disease. Rev. Sci. Tech. (Int. Off. Epizoot.) 2000, 19, 405–424. [Google Scholar] [CrossRef]
- Snoeyenbos, G. Pullorum disease. In Diseases of Poultry; Iowa State University Press: Ames, IA, USA, 1991; pp. 72–86. [Google Scholar]
- Schleifer, J.; Juven, B.; Beard, C.; Cox, N. The susceptibility of chicks to Salmonella montevideo in artificially contaminated poultry feed. Avian Dis. 1984, 28, 497–503. [Google Scholar] [CrossRef]
- Jones, F.; Axtell, R.; Tarver, F.; Rives, D.; Scheideler, S.; Wineland, M. Environmental Factors Contributing to Salmonella Colonization of Chickens. In Colonization Control of Human Bacterial Enteropathogens in Poultry; Blankenship, L.C., Ed.; Academic Press, Inc.: San Diego, CA, USA, 1991; pp. 3–20. [Google Scholar]
- Authority, E.F.S. The European Union One Health 2019 Zoonoses Report. Efsa J. 2021, 19, e06406. [Google Scholar] [CrossRef] [Green Version]
- Chimalizeni, Y.; Kawaza, K.; Molyneux, E. The epidemiology and management of non typhoidal salmonella infections. In Hot Topics in Infection and Immunity in Children VI; Springer: New York, NY, USA, 2010; pp. 33–46. [Google Scholar] [CrossRef]
- Chen, H.-M.; Wang, Y.; Su, L.-H.; Chiu, C.-H. Nontyphoid Salmonella infection: Microbiology, clinical features, and antimicrobial therapy. Pediatrics Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Suresh, G.; Das, R.K.; Kaur Brar, S.; Rouissi, T.; Avalos Ramirez, A.; Chorfi, Y.; Godbout, S. Alternatives to antibiotics in poultry feed: Molecular perspectives. Crit. Rev. Microbiol. 2018, 44, 318–335. [Google Scholar] [CrossRef]
- Reemers, S.S.; van Leenen, D.; Koerkamp, M.J.G.; van Haarlem, D.; van de Haar, P.; van Eden, W.; Vervelde, L. Early host responses to avian influenza A virus are prolonged and enhanced at transcriptional level depending on maturation of the immune system. Mol. Immunol. 2010, 47, 1675–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijerink, N. Improving Health of Broiler Chickens by Early Life Modulation of Immune Responsiveness and Intestinal Microbiota: Recruiting Natural Killer Cells for Increased Resistance; Utrecht University: Utrecht, The Netherlands, 2021. [Google Scholar]
- Wells, C.A.; Ravasi, T.; Faulkner, G.J.; Carninci, P.; Okazaki, Y.; Hayashizaki, Y.; Sweet, M.; Wainwright, B.J.; Hume, D.A. Genetic control of the innate immune response. BMC Immunol. 2003, 4, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, T.; Schmucker, S.S.; Bessei, W.; Grashorn, M.; Stefanski, V. Impact of housing environment on the immune system in chickens: A review. Animals 2020, 10, 1138. [Google Scholar] [CrossRef] [PubMed]
- Svihus, B. Function of the digestive system. J. Appl. Poult. Res. 2014, 23, 306–314. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Zhong, T.; Pandya, Y.; Joerger, R.D. 16S rRNA-based analysis of microbiota from the cecum of broiler chickens. Appl. Environ. Microbiol. 2002, 68, 124–137. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A. The role of the crop in the feeding behaviour of the domestic chicken. Anim. Behav. 1970, 18, 633–639. [Google Scholar] [CrossRef]
- Choi, J.; Kim, G.; Cha, C. Spatial heterogeneity and stability of bacterial community in the gastrointestinal tracts of broiler chickens. Poult. Sci. 2014, 93, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Konsak, B.M.; Stanley, D.; Haring, V.R.; Geier, M.S.; Hughes, R.J.; Howarth, G.S.; Crowley, T.M.; Moore, R.J. Identification of differential duodenal gene expression levels and microbiota abundance correlated with differences in energy utilisation in chickens. Anim. Prod. Sci. 2013, 53, 1269–1275. [Google Scholar] [CrossRef] [Green Version]
- Clench, M.H.; Mathias, J.R. The avian cecum: A review. Wilson Bull. 1995, 107, 93–121. [Google Scholar]
- Gong, J.; Si, W.; Forster, R.J.; Huang, R.; Yu, H.; Yin, Y.; Yang, C.; Han, Y. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca. FEMS Microbiol. Ecol. 2007, 59, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Owens, B.; Tucker, L.; Collins, M.; McCracken, K. Effects of different feed additives alone or in combination on broiler performance, gut microflora and ileal histology. Br. Poult. Sci. 2008, 49, 202–212. [Google Scholar] [CrossRef]
- Savage, D.C. Microbial ecology of the gastrointestinal tract. Annu. Rev. Microbiol. 1977, 31, 107–133. [Google Scholar] [CrossRef]
- Gaskins, H.R. Intestinal bacteria and their influence on swine growth. In Swine Nutrition, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2000; pp. 585–608. [Google Scholar]
- Apajalahti, J.; Kettunen, A.; Graham, H. Characteristics of the gastrointestinal microbial communities, with special reference to the chicken. World’s Poult. Sci. J. 2004, 60, 223–232. [Google Scholar] [CrossRef]
- Juricova, H.; Videnska, P.; Lukac, M.; Faldynova, M.; Babak, V.; Havlickova, H.; Sisak, F.; Rychlik, I. Influence of Salmonella enterica serovar enteritidis infection on the development of the cecum microbiota in newly hatched chicks. Appl. Environ. Microbiol. 2013, 79, 745–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varmuzova, K.; Kubasova, T.; Davidova-Gerzova, L.; Sisak, F.; Havlickova, H.; Sebkova, A.; Faldynova, M.; Rychlik, I. Composition of gut microbiota influences resistance of newly hatched chickens to Salmonella Enteritidis infection. Front. Microbiol. 2016, 7, 957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanikawa, T.; Shoji, N.; Sonohara, N.; Saito, S.; Shimura, Y.; Fukushima, J.; Inamoto, T. Aging transition of the bacterial community structure in the chick ceca. Poult. Sci. 2011, 90, 1004–1008. [Google Scholar] [CrossRef]
- Stanley, D.; Geier, M.S.; Hughes, R.J.; Denman, S.E.; Moore, R.J. Highly variable microbiota development in the chicken gastrointestinal tract. PLoS ONE 2013, 8, e84290. [Google Scholar]
- Mon, K.K.; Saelao, P.; Halstead, M.M.; Chanthavixay, G.; Chang, H.-C.; Garas, L.; Maga, E.A.; Zhou, H. Salmonella enterica serovars Enteritidis infection alters the indigenous microbiota diversity in young layer chicks. Front. Vet. Sci. 2015, 2, 61. [Google Scholar] [CrossRef] [Green Version]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef]
- Bansil, R.; Turner, B.S. Mucin structure, aggregation, physiological functions and biomedical applications. Curr. Opin. Colloid Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Sellers, L.A.; Allen, A.; Morris, E.R.; Ross-Murphy, S.B. Mucus glycoprotein gels. Role of glycoprotein polymeric structure and carbohydrate side-chains in gel-formation. Carbohydr. Res. 1988, 178, 93–110. [Google Scholar] [CrossRef]
- Schumacher, G.; Kim, M.; Hosseinian, A.; Dupon, C. Immunoglobulins, proteinase inhibitors, albumin, and lysozyme in human cervical mucus: I. Communication: Hormonal profiles and cervical mucus changes—Methods and results. Am. J. Obstet. Gynecol. 1977, 129, 629–636. [Google Scholar] [CrossRef]
- Jones, D.E.; Bevins, C.L. Paneth cells of the human small intestine express an antimicrobial peptide gene. J. Biol. Chem. 1992, 267, 23216–23225. [Google Scholar] [CrossRef]
- Mukherjee, S.; Zheng, H.; Derebe, M.G.; Callenberg, K.M.; Partch, C.L.; Rollins, D.; Propheter, D.C.; Rizo, J.; Grabe, M.; Jiang, Q.-X. Antibacterial membrane attack by a pore-forming intestinal C-type lectin. Nature 2014, 505, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.; Bell, A.; Mantle, M.; Pearson, J.P. The structure and physiology of gastrointestinal mucus. In Mucus Health and Disease II; Springer: Boston, MA, USA, 1982; pp. 115–133. [Google Scholar]
- Lang, T.; Hansson, G.C.; Samuelsson, T. An inventory of mucin genes in the chicken genome shows that the mucin domain of Muc13 is encoded by multiple exons and that ovomucin is part of a locus of related gel-forming mucins. BMC Genom. 2006, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yegani, M.; Korver, D. Factors affecting intestinal health in poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef]
- Cheng, H.; Leblond, C. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine I. Columnar cell. Am. J. Anat. 1974, 141, 461–479. [Google Scholar] [CrossRef]
- Uni, Z.; Smirnov, A.; Sklan, D. Pre-and posthatch development of goblet cells in the broiler small intestine: Effect of delayed access to feed. Poult. Sci. 2003, 82, 320–327. [Google Scholar] [CrossRef]
- Rehfeld, J. A centenary of gastrointestinal endocrinology. Horm. Metab. Res. 2004, 36, 735–741. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Li, J., Jr.; Li, R.; Lv, C.; Li, S.; Mi, Y.; Zhang, C. Identification of the Paneth cells in chicken small intestine. Poult. Sci. 2016, 95, 1631–1635. [Google Scholar] [CrossRef] [PubMed]
- Nile, C.; Townes, C.; Michailidis, G.; Hirst, B.; Hall, J. Identification of chicken lysozyme g 2 and its expression in the intestine. Cell. Mol. Life Sci. 2004, 61, 2760–2766. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Graf, T. Goose-type lysozyme gene of the chicken: Sequence, genomic organization and expression reveals major differences to chicken-type lysozyme gene. Biochim. Biophys. Acta 1991, 1090, 273–276. [Google Scholar] [CrossRef]
- Smith, A.L.; Powers, C.; Beal, R.K. The avian enteric immune system in health and disease. In Avian Immunology, 2nd ed.; Elsevier: London, UK, 2014; pp. 227–250. [Google Scholar]
- Liebler-Tenorio, E.M.; Pabst, R. MALT structure and function in farm animals. Vet. Res. 2006, 37, 257–280. [Google Scholar] [CrossRef] [Green Version]
- Befus, A.D.; Johnston, N.; Leslie, G.; Bienenstock, J. Gut-associated lymphoid tissue in the chicken. I. Morphology, ontogeny, and some functional characteristics of Peyer’s patches. J. Immunol. 1980, 125, 2626–2632. [Google Scholar]
- Vervelde, L.; Jeurissen, S. Postnatal development of intra-epithelial leukocytes in the chicken digestive tract: Phenotypical characterization in situ. Cell Tissue Res. 1993, 274, 295–301. [Google Scholar] [CrossRef]
- Bockman, D.E.; Cooper, M.D. Pinocytosis by epithelium associated with lymphoid follicles in the bursa of Fabricius, appendix, and Peyer’s patches. An electron microscopic study. Am. J. Anat. 1973, 136, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Dalmaroni, M.J.; Gerswhin, M.E.; Adamopoulos, I.E. The critical role of toll-like receptors—from microbial recognition to autoimmunity: A comprehensive review. Autoimmun. Rev. 2016, 15, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H.; Iqbal, M.; He, H.; Philbin, V.; Kaiser, P.; Smith, A. Expression and function of Toll-like receptors in chicken heterophils. Dev. Comp. Immunol. 2005, 29, 791–807. [Google Scholar] [CrossRef]
- Kaiser, P.; Rothwell, L.; Galyov, E.E.; Barrow, P.A.; Burnside, J.; Wigley, P. Differential cytokine expression in avian cells in response to invasion by Salmonella typhimurium, Salmonella enteritidis and Salmonella gallinarum The GenBank accession numbers for the sequences reported in this paper are AI982185 for chicken IL-6 cDNA and AJ250838 for the partial chicken IL-6 genomic sequence, respectively. Microbiology 2000, 146, 3217–3226. [Google Scholar]
- Dil, N.; Qureshi, M. Differential expression of inducible nitric oxide synthase is associated with differential Toll-like receptor-4 expression in chicken macrophages from different genetic backgrounds. Vet. Immunol. Immunopathol. 2002, 84, 191–207. [Google Scholar] [CrossRef]
- Kogut, M.H.; Swaggerty, C.; He, H.; Pevzner, I.; Kaiser, P. Toll-like receptor agonists stimulate differential functional activation and cytokine and chemokine gene expression in heterophils isolated from chickens with differential innate responses. Microbes Infect. 2006, 8, 1866–1874. [Google Scholar] [CrossRef]
- Kogut, M.; Genovese, K.; He, H. Flagellin and lipopolysaccharide stimulate the MEK-ERK signaling pathway in chicken heterophils through differential activation of the small GTPases, Ras and Rap1. Mol. Immunol. 2007, 44, 1729–1736. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H. Analysis of Eimeria acervulina-induced changes in the intestinal T lymphocyte subpopulations in two chicken strains showing different levels of susceptibility to coccidiosis. Res. Vet. Sci. 1994, 56, 1–7. [Google Scholar] [CrossRef]
- Göbel, T.W.; Kaspers, B.; Stangassinger, M. NK and T cells constitute two major, functionally distinct intestinal epithelial lymphocyte subsets in the chicken. Int. Immunol. 2001, 13, 757–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kaer, L.; Olivares-Villagómez, D. Development, homeostasis, and functions of intestinal intraepithelial lymphocytes. J. Immunol. 2018, 200, 2235–2244. [Google Scholar] [CrossRef] [Green Version]
- Morvan, M.G.; Lanier, L.L. NK cells and cancer: You can teach innate cells new tricks. Nat. Rev. Cancer 2016, 16, 7–19. [Google Scholar] [CrossRef]
- Lodoen, M.B.; Lanier, L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006, 18, 391–398. [Google Scholar] [CrossRef]
- Bonanni, V.; Sciumè, G.; Santoni, A.; Bernardini, G. Bone marrow NK cells: Origin, distinctive features, and requirements for tissue localization. Front. Immunol. 2019, 10, 1569. [Google Scholar] [CrossRef] [Green Version]
- Göbel, T.W.; Chen, C.L.H.; Shrimpf, J.; Grossi, C.E.; Bernot, A.; Bucy, R.P.; Auffray, C.; Cooper, M.D. Characterization of avian natural killer cells and their intracellular CD3 protein complex. Eur. J. Immunol. 1994, 24, 1685–1691. [Google Scholar] [CrossRef] [Green Version]
- Chai, J.; Lillehoj, H. Isolation and functional characterization of chicken intestinal intra-epithelial lymphocytes showing natural killer cell activity against tumour target cells. Immunology 1988, 63, 111. [Google Scholar]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.A.; van de Haar, P.M.; van Haarlem, D.; van Kooten, P.; de Wit, S.; van Eden, W.; Viertlböck, B.C.; Göbel, T.W.; Vervelde, L. Identification of new populations of chicken natural killer (NK) cells. Dev. Comp. Immunol. 2010, 34, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Jansen, C.A.; De Geus, E.D.; Van Haarlem, D.A.; Van De Haar, P.M.; Löndt, B.Z.; Graham, S.P.; Göbel, T.W.; Van Eden, W.; Brookes, S.M.; Vervelde, L. Differential lung NK cell responses in avian influenza virus infected chickens correlate with pathogenicity. Sci. Rep. 2013, 3, 2478. [Google Scholar] [CrossRef]
- Vervelde, L.; Matthijs, M.; Van Haarlem, D.; de Wit, J.; Jansen, C. Rapid NK-cell activation in chicken after infection with infectious bronchitis virus M41. Vet. Immunol. Immunopathol. 2013, 151, 337–341. [Google Scholar] [CrossRef]
- Meijerink, N.; van Haarlem, D.A.; Velkers, F.C.; Stegeman, A.J.; Rutten, V.P.; Jansen, C.A. Analysis of chicken intestinal natural killer cells, a major IEL subset during embryonic and early life. Dev. Comp. Immunol. 2021, 114, 103857. [Google Scholar] [CrossRef]
- Zhang, L.; Katselis, G.S.; Moore, R.E.; Lekpor, K.; Goto, R.M.; Hunt, H.D.; Lee, T.D.; Miller, M.M. MHC class I target recognition, immunophenotypes and proteomic profiles of natural killer cells within the spleens of day-14 chick embryos. Dev. Comp. Immunol. 2012, 37, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Sivori, S.; Vacca, P.; Del Zotto, G.; Munari, E.; Mingari, M.C.; Moretta, L. Human NK cells: Surface receptors, inhibitory checkpoints, and translational applications. Cell. Mol. Immunol. 2019, 16, 430–441. [Google Scholar] [CrossRef]
- Viertlboeck, B.C.; Habermann, F.A.; Schmitt, R.; Groenen, M.A.; Du Pasquier, L.; Göbel, T.W. The chicken leukocyte receptor complex: A highly diverse multigene family encoding at least six structurally distinct receptor types. J. Immunol. 2005, 175, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Chiang, H.-I.; Zhou, H.; Raudsepp, T.; Jesudhasan, P.R.; Zhu, J.J. Chicken CD69 and CD94/NKG2-like genes in a chromosomal region syntenic to mammalian natural killer gene complex. Immunogenetics 2007, 59, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Viertlboeck, B.C.; Göbel, T.W. The chicken leukocyte receptor cluster. Vet. Immunol. Immunopathol. 2011, 144, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Biassoni, R. Natural killer cell receptors. In Multichain Immune Recognition Receptor Signaling; Springer: New York, NY, USA, 2008; pp. 35–52. [Google Scholar]
- Luci, C.; Reynders, A.; Ivanov, I.I.; Cognet, C.; Chiche, L.; Chasson, L.; Hardwigsen, J.; Anguiano, E.; Banchereau, J.; Chaussabel, D. Influence of the transcription factor RORγt on the development of NKp46+ cell populations in gut and skin. Nat. Immunol. 2009, 10, 75–82. [Google Scholar] [CrossRef]
- Mayassi, T.; Jabri, B. Human intraepithelial lymphocytes. Mucosal Immunol. 2018, 11, 1281–1289. [Google Scholar] [CrossRef]
- Orange, J.S. Formation and function of the lytic NK-cell immunological synapse. Nat. Rev. Immunol. 2008, 8, 713–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solana, R.; Tarazona, R.; Gayoso, I.; Lesur, O.; Dupuis, G.; Fulop, T. Innate immunosenescence: Effect of aging on cells and receptors of the innate immune system in humans. In Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2012; Volume 24, pp. 331–341. [Google Scholar]
- Jahromi, M.Z.; Bello, M.B.; Abdolmaleki, M.; Yeap, S.K.; Hair-Bejo, M.; Omar, A.R. Differential activation of intraepithelial lymphocyte-natural killer cells in chickens infected with very virulent and vaccine strains of infectious bursal disease virus. Dev. Comp. Immunol. 2018, 87, 116–123. [Google Scholar] [CrossRef]
- Deeg, C.A.; Degroote, R.L.; Giese, I.M.; Hirmer, S.; Amann, B.; Weigand, M.; Wiedemann, C.; Hauck, S.M. CD11d is a novel antigen on chicken leukocytes. J. Proteom. 2020, 225, 103876. [Google Scholar] [CrossRef] [PubMed]
- Bertzbach, L.D.; van Haarlem, D.A.; Härtle, S.; Kaufer, B.B.; Jansen, C.A. Marek’s disease virus infection of natural killer cells. Microorganisms 2019, 7, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poggi, A.; Benelli, R.; Venè, R.; Costa, D.; Ferrari, N.; Tosetti, F.; Zocchi, M.R. Human gut-associated natural killer cells in health and disease. Front. Immunol. 2019, 10, 961. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, S.A.; Tracey, D.E.; Henney, C.S. Induction of “natural killer” cells by BCG. Nature 1976, 262, 584–586. [Google Scholar] [CrossRef]
- Baron, E.J.; Proctor, R.A. Inefficient in vitro killing of virulent or nonvirulent Salmonella typhimurium by murine polymorphonuclear neutrophils. Can. J. Microbiol. 1984, 30, 1264–1270. [Google Scholar] [CrossRef]
- Maxwell, M.; Robertson, G. The avian heterophil leucocyte: A review. World’s Poult. Sci. J. 1998, 54, 155–178. [Google Scholar] [CrossRef]
- Kogut, M.; McGruder, E.; Hargis, B.; Corrier, D.; DeLoach, J. Dynamics of avian inflammatory response to Salmonella-immune lymphokines. Inflammation 1994, 18, 373–388. [Google Scholar] [CrossRef]
- Kogut, M.H.; McGruder, E.D.; Hargis, B.M.; Corner, D.E.; DeLoach, J.R. In vivo activation of heterophil function in chickens following injection with Salmonella enteritidis-immune lymphokines. J. Leukoc. Biol. 1995, 57, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H.; Mcgruder, E.D.; Hargis, B.M.; Corrier, D.E.; Deloach, J.R. Characterization of the pattern of inflammatory cell influx in chicks following the intraperitoneal administration of live Salmonella enteritidis and Salmonella enteritidis-immune lymphokines. Poult. Sci. 1995, 74, 8–17. [Google Scholar] [CrossRef]
- Kogut, M.; Holtzapple, C.; Lowry, V.; Genovese, K.; Stanker, L. Functional responses of neonatal chicken and turkey heterophils following stimulation by inflammatory agonists. Am. J. Vet. Res. 1998, 59, 1404–1408. [Google Scholar] [PubMed]
- Stabler, J.; McCormick, T.; Powell, K.; Kogut, M. Avian heterophils and monocytes: Phagocytic and bactericidal activities against Salmonella enteritidis. Vet. Microbiol. 1994, 38, 293–305. [Google Scholar] [CrossRef]
- Bar-Shira, E.; Friedman, A. Development and adaptations of innate immunity in the gastrointestinal tract of the newly hatched chick. Dev. Comp. Immunol. 2006, 30, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Matulova, M.; Varmuzova, K.; Sisak, F.; Havlickova, H.; Babak, V.; Stejskal, K.; Zdrahal, Z.; Rychlik, I. Chicken innate immune response to oral infection with Salmonella enterica serovar Enteritidis. Vet. Res. 2013, 44, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H.; Rothwell, L.; Kaiser, P. IFN-γ priming of chicken heterophils upregulates the expression of proinflammatory and Th1 cytokine mRNA following receptor-mediated phagocytosis of Salmonella enterica serovar enteritidis. J. Interferon Cytokine Res. 2005, 25, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Radsak, M.P.; Salih, H.R.; Rammensee, H.-G.; Schild, H. Triggering receptor expressed on myeloid cells-1 in neutrophil inflammatory responses: Differential regulation of activation and survival. J. Immunol. 2004, 172, 4956–4963. [Google Scholar] [CrossRef] [Green Version]
- Genovese, K.J.; He, H.; Swaggerty, C.L.; Kogut, M.H. The avian heterophil. Dev. Comp. Immunol. 2013, 41, 334–340. [Google Scholar] [CrossRef]
- Chuammitri, P.; Ostojić, J.; Andreasen, C.B.; Redmond, S.B.; Lamont, S.J.; Palić, D. Chicken heterophil extracellular traps (HETs): Novel defense mechanism of chicken heterophils. Vet. Immunol. Immunopathol. 2009, 129, 126–131. [Google Scholar] [CrossRef]
- Guimaraes-Costa, A.B.; Nascimento, M.T.; Wardini, A.B.; Pinto-da-Silva, L.H.; Saraiva, E.M. ETosis: A microbicidal mechanism beyond cell death. J. Parasitol. Res. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Wells, L.L.; Lowry, V.K.; Deloach, J.R.; Kogut, M.H. Age-dependent phagocytosis and bactericidal activities of the chicken heterophil. Dev. Comp. Immunol. 1998, 22, 103–109. [Google Scholar] [CrossRef]
- Daimon, T.; Caxton-Martins, A. Electron microscopic and enzyme cytochemical studies on granules of mature chicken granular leucocytes. J. Anat. 1977, 123, 553. [Google Scholar]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Farache, J.; Zigmond, E.; Shakhar, G.; Jung, S. Contributions of dendritic cells and macrophages to intestinal homeostasis and immune defense. Immunol. Cell Biol. 2013, 91, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Garceau, V.; Smith, J.; Paton, I.R.; Davey, M.; Fares, M.A.; Sester, D.P.; Burt, D.W.; Hume, D.A. Pivotal Advance: Avian colony-stimulating factor 1 (CSF-1), interleukin-34 (IL-34), and CSF-1 receptor genes and gene products. J. Leukoc. Biol. 2010, 87, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamoutounour, S.; Henri, S.; Lelouard, H.; de Bovis, B.; de Haar, C.; van der Woude, C.J.; Woltman, A.M.; Reyal, Y.; Bonnet, D.; Sichien, D. CD 64 distinguishes macrophages from dendritic cells in the gut and reveals the T h1-inducing role of mesenteric lymph node macrophages during colitis. Eur. J. Immunol. 2012, 42, 3150–3166. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gálvez, S.G.; Tovar-Torres, S.M.; Tron-Gómez, M.S.; Weiser-Smeke, A.E.; Álvarez-Hernández, D.A.; Franyuti-Kelly, G.A.; Tapia-Moreno, M.; Ibarra, A.; Gutiérrez-Kobeh, L.; Vázquez-López, R. Human dendritic cells: Ontogeny and their subsets in health and disease. Med Sci. 2018, 6, 88. [Google Scholar] [CrossRef] [Green Version]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The dendritic cell lineage: Ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [Green Version]
- Nagy, N.; Bódi, I.; Oláh, I. Avian dendritic cells: Phenotype and ontogeny in lymphoid organs. Dev. Comp. Immunol. 2016, 58, 47–59. [Google Scholar] [CrossRef]
- Hansell, C.; Zhu, X.W.; Brooks, H.; Sheppard, M.; Withanage, S.; Maskell, D.; McConnell, I. Unique features and distribution of the chicken CD83+ cell. J. Immunol. 2007, 179, 5117–5125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Rothwell, L.; Young, J.R.; Kaufman, J.; Butter, C.; Kaiser, P. Generation and characterization of chicken bone marrow-derived dendritic cells. Immunology 2010, 129, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Staines, K.; Young, J.R.; Butter, C. Expression of chicken DEC205 reflects the unique structure and function of the avian immune system. PLoS ONE 2013, 8, e51799. [Google Scholar] [CrossRef] [Green Version]
- Manh, T.-P.V.; Marty, H.; Sibille, P.; Le Vern, Y.; Kaspers, B.; Dalod, M.; Schwartz-Cornil, I.; Quéré, P. Existence of conventional dendritic cells in Gallus gallus revealed by comparative gene expression profiling. J. Immunol. 2014, 192, 4510–4517. [Google Scholar] [CrossRef] [PubMed]
- Jeurissen, S.; Janse, E.; Lehrbach, P.; Haddad, E.; Avakian, A.; Whitfill, C. The working mechanism of an immune complex vaccine that protects chickens against infectious bursal disease. Immunology 1998, 95, 494. [Google Scholar] [CrossRef] [PubMed]
- Olah, I.; Glick, B. Structure of the germinal centers in the chicken caecal tonsil: Light and electron microscopic and autoradiographic studies. Poult. Sci. 1979, 58, 195–210. [Google Scholar] [CrossRef]
- Nagy, N.; Oláh, I. Pyloric tonsil as a novel gut-associated lymphoepithelial organ of the chicken. J. Anat. 2007, 211, 407–411. [Google Scholar] [CrossRef]
- Gallego, M.; Del Cacho, E.; Bascuas, J. Antigen-binding cells in the cecal tonsil and Peyer’s patches of the chicken after bovine serum albumin administration. Poult. Sci. 1995, 74, 472–479. [Google Scholar] [CrossRef]
- Abasht, B.; Kaiser, M.; Lamont, S. Toll-like receptor gene expression in cecum and spleen of advanced intercross line chicks infected with Salmonella enterica serovar Enteritidis. Vet. Immunol. Immunopathol. 2008, 123, 314–323. [Google Scholar] [CrossRef]
- Van den Biggelaar, R.H.; Arkesteijn, G.J.; Rutten, V.P.; van Eden, W.; Jansen, C.A. In vitro chicken bone marrow-derived dendritic cells comprise subsets at different states of maturation. Front. Immunol. 2020, 11, 141. [Google Scholar] [CrossRef]
- Zanoni, I.; Ostuni, R.; Marek, L.R.; Barresi, S.; Barbalat, R.; Barton, G.M.; Granucci, F.; Kagan, J.C. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell 2011, 147, 868–880. [Google Scholar] [CrossRef] [Green Version]
- Yitbarek, A.; Echeverry, H.; Brady, J.; Hernandez-Doria, J.; Camelo-Jaimes, G.; Sharif, S.; Guenter, W.; House, J.; Rodriguez-Lecompte, J. Innate immune response to yeast-derived carbohydrates in broiler chickens fed organic diets and challenged with Clostridium perfringens. Poult. Sci. 2012, 91, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Singh, M.; Chander, V.; Sharma, G.K.; Mahawar, M.; Teeli, A.S.; Goswami, T.K. Differential responses of chicken monocyte-derived dendritic cells infected with Salmonella Gallinarum and Salmonella Typhimurium. Sci. Rep. 2021, 11, 17214. [Google Scholar] [CrossRef]
- Rehman, M.U.; Yoshihisa, Y.; Miyamoto, Y.; Shimizu, T. The anti-inflammatory effects of platinum nanoparticles on the lipopolysaccharide-induced inflammatory response in RAW 264.7 macrophages. Inflamm. Res. 2012, 61, 1177–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, M.; Lillehoj, H.S.; Raybourne, R.B.; Babu, U.S.; Heckert, R.A.; Tani, H.; Sasai, K.; Baba, E.; Lillehoj, E.P. Differential responses of macrophages to Salmonella enterica serovars Enteritidis and Typhimurium. Vet. Immunol. Immunopathol. 2005, 107, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Withanage, G.; Mastroeni, P.; Brooks, H.; Maskell, D.; McConnell, I. Oxidative and nitrosative responses of the chicken macrophage cell line MQ-NCSU to experimental Salmonella infection. Br. Poult. Sci. 2005, 46, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Babu, U.S.; Gaines, D.W.; Lillehoj, H.; Raybourne, R.B. Differential reactive oxygen and nitrogen production and clearance of Salmonella serovars by chicken and mouse macrophages. Dev. Comp. Immunol. 2006, 30, 942–953. [Google Scholar] [CrossRef]
- He, H.; Genovese, K.J.; Nisbet, D.J.; Kogut, M.H. Profile of Toll-like receptor expressions and induction of nitric oxide synthesis by Toll-like receptor agonists in chicken monocytes. Mol. Immunol. 2006, 43, 783–789. [Google Scholar] [CrossRef] [PubMed]
- He, H.; MacKinnon, K.M.; Genovese, K.J.; Kogut, M.H. CpG oligodeoxynucleotide and double-stranded RNA synergize to enhance nitric oxide production and mRNA expression of inducible nitric oxide synthase, pro-inflammatory cytokines and chemokines in chicken monocytes. Innate Immun. 2011, 17, 137–144. [Google Scholar]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000prime Rep. 2014, 6, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Novak, M.L.; Koh, T.J. Macrophage phenotypes during tissue repair. J. Leukoc. Biol. 2013, 93, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Ma, J.; Chen, T.; Mandelin, J.; Ceponis, A.; Miller, N.; Hukkanen, M.; Ma, G.; Konttinen, Y. Regulation of macrophage activation. Cell. Mol. Life Sci. 2003, 60, 2334–2346. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luzina, I.G.; Keegan, A.D.; Heller, N.M.; Rook, G.A.; Shea-Donohue, T.; Atamas, S.P. Regulation of inflammation by interleukin-4: A review of “alternatives”. J. Leukoc. Biol. 2012, 92, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rőszer, T. Understanding the mysterious M2 macrophage through activation markers and effector mechanisms. Mediat. Inflamm. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; van den Biggelaar, R.H.; Jansen, C.A.; Haagsman, H.P.; Veldhuizen, E.J. A method to differentiate chicken monocytes into macrophages with proinflammatory properties. Immunobiology 2020, 225, 152004. [Google Scholar] [CrossRef]
- Chaudhari, A.A.; Kim, W.H.; Lillehoj, H.S. Interleukin-4 (IL-4) may regulate alternative activation of macrophage-like cells in chickens: A sequential study using novel and specific neutralizing monoclonal antibodies against chicken IL-4. Vet. Immunol. Immunopathol. 2018, 205, 72–82. [Google Scholar] [CrossRef]
- Gordon, S. Pattern recognition receptors: Doubling up for the innate immune response. Cell 2002, 111, 927–930. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.E.; Douglas, S.R. An experimental investigation of the role of the blood fluids in connection with phagocytosis. Proc. R. Soc. Lond. 1904, 72, 357–370. [Google Scholar] [CrossRef]
- Janeway, C.A. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54, 1–13. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, L.M.; Ezekowitz, R.A.B. Phagocytosis: Elegant complexity. Immunity 2005, 22, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Yamamoto, M.; Takeda, K. Toll-like receptor family: Receptors essential for microbial recognition and immune responses. Arthritis Res Ther 2003, 5, 1–54. [Google Scholar] [CrossRef]
- He, H.; Genovese, K.J.; Nisbet, D.J.; Kogut, M.H. Synergy of CpG oligodeoxynucleotide and double-stranded RNA (poly I: C) on nitric oxide induction in chicken peripheral blood monocytes. Mol. Immunol. 2007, 44, 3234–3242. [Google Scholar] [CrossRef]
- Hartley, C.; Salisbury, A.-M.; Wigley, P. CpG oligonucleotides and recombinant interferon-γ in combination improve protection in chickens to Salmonella enterica serovar Enteritidis challenge as an adjuvant component, but have no effect in reducing Salmonella carriage in infected chickens. Avian Pathol. 2012, 41, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Lillehoj, H.; Kim, C.-H.; Keeler, C., Jr.; Babu, U.; Zhang, M. Transcriptional response of chicken macrophages to Salmonella enterica serovar enteritidis infection. In Animal Genomics for Animal Health; Karger Publishers: Berlin, Germany, 2008; Volume 132, pp. 141–151. [Google Scholar]
- Kim, Y.-J.; Shin, Y.; Lee, K.H.; Kim, T.-J. Anethum graveloens flower extracts inhibited a lipopolysaccharide-induced inflammatory response by blocking iNOS expression and NF-κB activity in macrophages. Biosci. Biotechnol. Biochem. 2012, 76, 1122–1127. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Scheenstra, M.R.; van Dijk, A.; Veldhuizen, E.J.; Haagsman, H.P. A new and efficient culture method for porcine bone marrow-derived M1-and M2-polarized macrophages. Vet. Immunol. Immunopathol. 2018, 200, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Djeraba, A.; Musset, E.; van Rooijen, N.; Quéré, P. Resistance and susceptibility to Marek’s disease: Nitric oxide synthase/arginase activity balance. Vet. Microbiol. 2002, 86, 229–244. [Google Scholar] [CrossRef]
- He, H.; Kogut, M.H. CpG-ODN-induced nitric oxide production is mediated through clathrin-dependent endocytosis, endosomal maturation, and activation of PKC, MEK1/2 and p38 MAPK, and NF-κB pathways in avian macrophage cells (HD11). Cell. Signal. 2003, 15, 911–917. [Google Scholar] [CrossRef]
- Khatri, M.; Palmquist, J.M.; Cha, R.M.; Sharma, J.M. Infection and activation of bursal macrophages by virulent infectious bursal disease virus. Virus Res. 2005, 113, 44–50. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Li, G. Nitric oxide production by macrophages stimulated with coccidia sporozoites, lipopolysaccharide, or interferon-γ, and its dynamic changes in SC and TK strains of chickens infected with Eimeria tenella. Avian Dis. 2004, 48, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M. Avian macrophage and immune response: An overview. Poult. Sci. 2003, 82, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.K.; Kaiser, P.; Rothwell, L.; Humphrey, T.; Barrow, P.A.; Jones, M.A. Campylobacter jejuni-induced cytokine responses in avian cells. Infect. Immun. 2005, 73, 2094–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Biggelaar, R.H.; Van Eden, W.; Rutten, V.P.; Jansen, C.A. Nitric oxide production and Fc receptor-mediated phagocytosis as functional readouts of macrophage activity upon stimulation with inactivated poultry vaccines in vitro. Vaccines 2020, 8, 332. [Google Scholar] [CrossRef]
- Van den Biggelaar, R.H.; van Eden, W.; Rutten, V.P.; Jansen, C.A. Macrophage Activation Assays to Evaluate the Immunostimulatory Capacity of Avibacterium paragallinarum in A Multivalent Poultry Vaccine. Vaccines 2020, 8, 671. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Trout, J.M. Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 1996, 9, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Wigley, P.; Hulme, S.; Rothwell, L.; Bumstead, N.; Kaiser, P.; Barrow, P. Macrophages isolated from chickens genetically resistant or susceptible to systemic salmonellosis show magnitudinal and temporal differential expression of cytokines and chemokines following Salmonella enterica challenge. Infect. Immun. 2006, 74, 1425–1430. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Genovese, K.J.; Kogut, M.H. Modulation of chicken macrophage effector function by TH1/TH2 cytokines. Cytokine 2011, 53, 363–369. [Google Scholar] [CrossRef]
- Malkovsky, M.; Bartz, S.R.; Mackenzie, D.; Radtke, B.E.; Wallace, M.; Manning, J.; Pauza, C.D.; Fisch, P. Are γδ T cells important for the elimination of virus-infected cells? J. Med Primatol. 1992, 21, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Hayday, A.C. γδ cells: A right time and a right place for a conserved third way of protection. Annu. Rev. Immunol. 2000, 18, 975–1026. [Google Scholar] [CrossRef] [PubMed]
- Sowder, J.T.; Chen, C.; Ager, L.L.; Chan, M.M.; Cooper, M. A large subpopulation of avian T cells express a homologue of the mammalian T gamma/delta receptor. J. Exp. Med. 1988, 167, 315–322. [Google Scholar] [CrossRef]
- Mackay, C.R.; Hein, W.R. A large proportion of bovine T cells express the γδ T cell receptor and show a distinct tissue distribution and surface phenotype. Int. Immunol. 1989, 1, 540–545. [Google Scholar] [CrossRef]
- Yang, H.; Parkhouse, R. Phenotypic classification of porcine lymphocyte subpopulations in blood and lymphoid tissues. Immunology 1996, 89, 76–83. [Google Scholar] [CrossRef]
- Kubota, T.; Wang, J.-y.; Göbel, T.W.; Hockett, R.D.; Cooper, M.D.; Chen-lo, H.C. Characterization of an avian (Gallus gallus domesticus) TCR αδ gene locus. J. Immunol. 1999, 163, 3858–3866. [Google Scholar]
- Pieper, J.; Methner, U.; Berndt, A. Heterogeneity of avian γδ T cells. Vet. Immunol. Immunopathol. 2008, 124, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Fenzl, L.; Göbel, T.W.; Neulen, M.-L. γδ T cells represent a major spontaneously cytotoxic cell population in the chicken. Dev. Comp. Immunol. 2017, 73, 175–183. [Google Scholar] [CrossRef]
- Holderness, J.; Hedges, J.F.; Ramstead, A.; Jutila, M.A. Comparative biology of γδ T cell function in humans, mice, and domestic animals. Annu. Rev. Anim. Biosci. 2013, 1, 99–124. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, C.L.; Yirsaw, A.; Gillespie, A.; Le Page, L.; Zhang, F.; Damani-Yokota, P.; Telfer, J.C. γδ T cells in livestock: Responses to pathogens and vaccine potential. Transbound. Emerg. Dis. 2020, 67, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, M.M.; Willcox, C.R.; Salim, M.; Paletta, D.; Fichtner, A.S.; Noll, A.; Starick, L.; Nöhren, A.; Begley, C.R.; Berwick, K.A. Butyrophilin-2A1 directly binds germline-encoded regions of the Vγ9Vδ2 TCR and is essential for phosphoantigen sensing. Immunity 2020, 52, 487–498.e486. [Google Scholar] [CrossRef] [PubMed]
- Shibata, K.; Yamada, H.; Nakamura, M.; Hatano, S.; Katsuragi, Y.; Kominami, R.; Yoshikai, Y. IFN-γ–Producing and IL-17–Producing γδ T Cells Differentiate at Distinct Developmental Stages in Murine Fetal Thymus. J. Immunol. 2014, 192, 2210–2218. [Google Scholar] [CrossRef] [Green Version]
- Price, S.J.; Sopp, P.; Howard, C.J.; Hope, J.C. Workshop cluster 1+ γδ T-cell receptor+ T cells from calves express high levels of interferon-γ in response to stimulation with interleukin-12 and-18. Immunology 2007, 120, 57–65. [Google Scholar] [CrossRef]
- Alvarez, A.; Endsley, J.; Werling, D.; Mark Estes, D. WC1+ γδ T Cells Indirectly Regulate Chemokine Production During Mycobacterium bovis Infection in SCID-bo Mice. Transbound. Emerg. Dis. 2009, 56, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, M.Z.; Yan, L.; Lillehoj, H.; Pace, L.W.; Zhang, S. Induction of CXC chemokine messenger-RNA expression in chicken oviduct epithelial cells by salmonella enterica serovar enteritidis via the type three secretion system–1. Avian Dis. 2009, 53, 396–404. [Google Scholar] [CrossRef]
- Withanage, G.; Wigley, P.; Kaiser, P.; Mastroeni, P.; Brooks, H.; Powers, C.; Beal, R.; Barrow, P.; Maskell, D.; McConnell, I. Cytokine and chemokine responses associated with clearance of a primary Salmonella enterica serovar Typhimurium infection in the chicken and in protective immunity to rechallenge. Infect. Immun. 2005, 73, 5173–5182. [Google Scholar] [CrossRef] [Green Version]
- Setta, A.; Barrow, P.; Kaiser, P.; Jones, M. Early immune dynamics following infection with Salmonella enterica serovars Enteritidis, Infantis, Pullorum and Gallinarum: Cytokine and chemokine gene expression profile and cellular changes of chicken cecal tonsils. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 397–410. [Google Scholar] [CrossRef]
- Withanage, G.; Kaiser, P.; Wigley, P.; Powers, C.; Mastroeni, P.; Brooks, H.; Barrow, P.; Smith, A.; Maskell, D.; McConnell, I. Rapid expression of chemokines and proinflammatory cytokines in newly hatched chickens infected with Salmonella enterica serovar typhimurium. Infect. Immun. 2004, 72, 2152–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gart, E.V.; Suchodolski, J.S.; Welsh, T.H., Jr.; Alaniz, R.C.; Randel, R.D.; Lawhon, S.D. Salmonella typhimurium and multidirectional communication in the gut. Front. Microbiol. 2016, 7, 1827. [Google Scholar] [CrossRef] [Green Version]
- Darwin, K.H.; Miller, V.L. Molecular basis of the interaction of Salmonella with the intestinal mucosa. Clin. Microbiol. Rev. 1999, 12, 405–428. [Google Scholar] [CrossRef] [Green Version]
- Van Asten, A.J.; van Dijk, J.E. Distribution of “classic” virulence factors among Salmonella spp. FEMS Immunol. Med Microbiol. 2005, 44, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Marlovits, T.C.; Stebbins, C.E. Type III secretion systems shape up as they ship out. Curr. Opin. Microbiol. 2010, 13, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.; Lynne, A.; Nayak, R. Salmonella challenges: Prevalence in swine and poultry and potential pathogenicity of such isolates. J. Anim. Sci. 2008, 86, E149–E162. [Google Scholar] [CrossRef]
- Winnen, B.; Schlumberger, M.C.; Sturm, A.; Schüpbach, K.; Siebenmann, S.; Jenny, P.; Hardt, W.-D. Hierarchical effector protein transport by the Salmonella Typhimurium SPI-1 type III secretion system. PLoS ONE 2008, 3, e2178. [Google Scholar] [CrossRef]
- Schlumberger, M.C.; Hardt, W.-D. Salmonella type III secretion effectors: Pulling the host cell’s strings. Curr. Opin. Microbiol. 2006, 9, 46–54. [Google Scholar] [CrossRef]
- Marlovits, T.C.; Kubori, T.; Sukhan, A.; Thomas, D.R.; Galán, J.E.; Unger, V.M. Structural insights into the assembly of the type III secretion needle complex. Science 2004, 306, 1040–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galán, J.E. Salmonella interactions with host cells: Type III secretion at work. Annu. Rev. Cell Dev. Biol. 2001, 17, 53–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lostroh, C.P.; Lee, C.A. The Salmonella pathogenicity island-1 type III secretion system. Microbes Infect. 2001, 3, 1281–1291. [Google Scholar] [CrossRef]
- Zhou, D.; Chen, L.M.; Hernandez, L.; Shears, S.B.; Galán, J.E. A Salmonella inositol polyphosphatase acts in conjunction with other bacterial effectors to promote host cell actin cytoskeleton rearrangements and bacterial internalization. Mol. Microbiol. 2001, 39, 248–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardt, W.-D.; Chen, L.-M.; Schuebel, K.E.; Bustelo, X.R.; Galán, J.E.S. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells. Cell 1998, 93, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Friebel, A.; Ilchmann, H.; Aepfelbacher, M.; Ehrbar, K.; Machleidt, W.; Hardt, W.-D. SopE and SopE2 from Salmonella typhimurium activate different sets of RhoGTPases of the host cell. J. Biol. Chem. 2001, 276, 34035–34040. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Philbin, V.J.; Withanage, G.; Wigley, P.; Beal, R.K.; Goodchild, M.J.; Barrow, P.; McConnell, I.; Maskell, D.J.; Young, J. Identification and functional characterization of chicken toll-like receptor 5 reveals a fundamental role in the biology of infection with Salmonella enterica serovar typhimurium. Infect. Immun. 2005, 73, 2344–2350. [Google Scholar] [CrossRef] [Green Version]
- Chappell, L.; Kaiser, P.; Barrow, P.; Jones, M.A.; Johnston, C.; Wigley, P. The immunobiology of avian systemic salmonellosis. Vet. Immunol. Immunopathol. 2009, 128, 53–59. [Google Scholar] [CrossRef]
- Kogut, M.H.; Genovese, K.J.; Lowry, V.K. Differential activation of signal transduction pathways mediating phagocytosis, oxidative burst, and degranulation by chicken heterophils in response to stimulation with opsonized Salmonella enteritidis. Inflammation 2001, 25, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Swaggerty, C.L.; Pevzner, I.Y.; Lowry, V.K.; Farnell, M.B.; Kogut, M.H. Functional comparison of heterophils isolated from commercial broiler chickens. Avian Pathol. 2003, 32, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, A.; Tersteeg-Zijderveld, M.H.; Tjeerdsma-van Bokhoven, J.L.; Jansman, A.J.; Veldhuizen, E.J.; Haagsman, H.P. Chicken heterophils are recruited to the site of Salmonella infection and release antibacterial mature Cathelicidin-2 upon stimulation with LPS. Mol. Immunol. 2009, 46, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Pieper, J.; Locke, M.; Ruzaike, G.; Voigt, S.; Methner, U.; Berndt, A. In vitro and in vivo generation of heterophil extracellular traps after Salmonella exposure. Vet. Immunol. Immunopathol. 2017, 188, 1–11. [Google Scholar] [CrossRef]
- Steiner, T.S. How flagellin and toll-like receptor 5 contribute to enteric infection. Infect. Immun. 2007, 75, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.-S.; Lee, H.; Lee, J.-O. The structural basis of lipopolysaccharide recognition by the TLR4–MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar] [CrossRef]
- Yoon, S.-i.; Kurnasov, O.; Natarajan, V.; Hong, M.; Gudkov, A.V.; Osterman, A.L.; Wilson, I.A. Structural basis of TLR5-flagellin recognition and signaling. Science 2012, 335, 859–864. [Google Scholar] [CrossRef] [Green Version]
- McDermott, P.F.; Ciacci-Woolwine, F.; Snipes, J.A.; Mizel, S.B. High-affinity interaction between gram-negative flagellin and a cell surface polypeptide results in human monocyte activation. Infect. Immun. 2000, 68, 5525–5529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaves-Pyles, T.; Murthy, K.; Liaudet, L.; Virág, L.; Ross, G.; Soriano, F.G.; Szabó, C.; Salzman, A.L. Flagellin, a novel mediator of Salmonella-induced epithelial activation and systemic inflammation: IκBα degradation, induction of nitric oxide synthase, induction of proinflammatory mediators, and cardiovascular dysfunction. J. Immunol. 2001, 166, 1248–1260. [Google Scholar] [CrossRef] [Green Version]
- Gewirtz, A.T.; Navas, T.A.; Lyons, S.; Godowski, P.J.; Madara, J.L. Cutting edge: Bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J. Immunol. 2001, 167, 1882–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Means, T.K.; Hayashi, F.; Smith, K.D.; Aderem, A.; Luster, A.D. The Toll-like receptor 5 stimulus bacterial flagellin induces maturation and chemokine production in human dendritic cells. J. Immunol. 2003, 170, 5165–5175. [Google Scholar] [CrossRef]
- Khan, M.A.; Kang, J.; Steiner, T.S. Enteroaggregative Escherichia coli flagellin-induced interleukin-8 secretion requires Toll-like receptor 5-dependent p38 MAP kinase activation. Immunology 2004, 112, 651–660. [Google Scholar] [CrossRef]
- Crippen, T.L. The selective inhibition of nitric oxide production in the avian macrophage cell line HD11. Vet. Immunol. Immunopathol. 2006, 109, 127–137. [Google Scholar] [CrossRef]
- Beal, R.; Wigley, P.; Powers, C.; Hulme, S.; Barrow, P.; Smith, A. Age at primary infection with Salmonella enterica serovar Typhimurium in the chicken influences persistence of infection and subsequent immunity to re-challenge. Vet. Immunol. Immunopathol. 2004, 100, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Fresno, A.H.; Skov, S.; Olsen, J.E. Dynamics and outcome of macrophage interaction between Salmonella gallinarum, Salmonella Typhimurium, and Salmonella dublin and macrophages from chicken and cattle. Front. Cell. Infect. Microbiol. 2020, 9, 420. [Google Scholar] [CrossRef]
- Swaggerty, C.L.; Kogut, M.H.; Ferro, P.J.; Rothwell, L.; Pevzner, I.Y.; Kaiser, P. Differential cytokine mRNA expression in heterophils isolated from Salmonella-resistant and-susceptible chickens. Immunology 2004, 113, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Pan, Q.; Hui, X.; Zhang, B.; Wu, H.; Li, H.; Xu, W.; Zhang, Q.; Li, J.; Deng, X. Stronger in vitro phagocytosis by monocytes-macrophages is indicative of greater pathogen clearance and antibody levels in vivo. Poult. Sci. 2008, 87, 1725–1733. [Google Scholar] [CrossRef]
- Johannessen, M.; Askarian, F.; Sangvik, M.; Sollid, J.E. Bacterial interference with canonical NFκB signalling. Microbiology 2013, 159, 2001. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Flohé, L. Basic principles and emerging concepts in the redox control of transcription factors. Antioxid. Redox Signal. 2011, 15, 2335–2381. [Google Scholar] [CrossRef] [Green Version]
- Bekiaris, V.; Persson, E.K.; Agace, W.W. Intestinal dendritic cells in the regulation of mucosal immunity. Immunol. Rev. 2014, 260, 86–101. [Google Scholar] [CrossRef]
- Vazquez-Torres, A.; Jones-Carson, J.; Bäumler, A.J.; Falkow, S.; Valdivia, R.; Brown, W.; Le, M.; Berggren, R.; Parks, W.T.; Fang, F.C. Extraintestinal dissemination of Salmonella by CD18-expressing phagocytes. Nature 1999, 401, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Swart, A.L.; Hensel, M. Interactions of Salmonella enterica with dendritic cells. Virulence 2012, 3, 660–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSorley, S.J.; Asch, S.; Costalonga, M.; Reinhardt, R.L.; Jenkins, M.K. Tracking salmonella-specific CD4 T cells in vivo reveals a local mucosal response to a disseminated infection. Immunity 2002, 16, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.L.; Raffatellu, M.; Bevins, C.L.; Adams, L.G.; Tükel, Ç.; Tsolis, R.M.; Bäumler, A.J. Life in the inflamed intestine, Salmonella style. Trends Microbiol. 2009, 17, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Mastroeni, P.; Ménager, N. Development of acquired immunity to Salmonella. J. Med Microbiol. 2003, 52, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Haller, D.; Serrant, P.; Granato, D.; Schiffrin, E.; Blum, S. Activation of human NK cells by staphylococci and lactobacilli requires cell contact-dependent costimulation by autologous monocytes. Clin. Vaccine Immunol. 2002, 9, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Carvajal, B.G.; Methner, U.; Pieper, J.; Berndt, A. Effects of Salmonella enterica serovar Enteritidis on cellular recruitment and cytokine gene expression in caecum of vaccinated chickens. Vaccine 2008, 26, 5423–5433. [Google Scholar] [CrossRef]
- Klasing, K.; Leshchinsky, T. Functions, costs, and benefits of the immune system during development and growth. Ostrich 1999, 69, 32. [Google Scholar]
- Meijerink, N.; van den Biggelaar, R.H.; van Haarlem, D.A.; Stegeman, J.A.; Rutten, V.P.; Jansen, C.A. A detailed analysis of innate and adaptive immune responsiveness upon infection with Salmonella enterica serotype Enteritidis in young broiler chickens. Vet. Res. 2021, 52, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Berndt, A.; Pieper, J.; Methner, U. Circulating γδ T Cells in Response to Salmonella enterica Serovar Enteritidis Exposure in Chickens. Infect. Immun. 2006, 74, 3967–3978. [Google Scholar] [CrossRef] [Green Version]
- Sekelova, Z.; Polansky, O.; Stepanova, H.; Fedr, R.; Faldynova, M.; Rychlik, I.; Vlasatikova, L. Different roles of CD4, CD8 and γδ T-lymphocytes in naive and vaccinated chickens during Salmonella Enteritidis infection. Proteomics 2017, 17, 1700073. [Google Scholar] [CrossRef]
- Schokker, D.; Peters, T.; Hoekman, A.; Rebel, J.; Smits, M. Differences in the early response of hatchlings of different chicken breeding lines to Salmonella enterica serovar Enteritidis infection. Poult. Sci. 2012, 91, 346–353. [Google Scholar] [CrossRef]
- Van Hemert, S.; Hoekman, A.J.; Smits, M.A.; Rebel, J.M. Immunological and gene expression responses to a Salmonella infection in the chicken intestine. Vet. Res. 2007, 38, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Deseke, M.; Prinz, I. Ligand recognition by the γδ TCR and discrimination between homeostasis and stress conditions. Cell. Mol. Immunol. 2020, 17, 914–924. [Google Scholar] [CrossRef]
- Bai, S.; Wu, A.; Ding, X.; Lei, Y.; Bai, J.; Zhang, K.; Chio, J. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poult. Sci. 2013, 92, 663–670. [Google Scholar] [CrossRef]
- Meijerink, N.; de Oliveira, J.E.; van Haarlem, D.A.; Hosotani, G.; Lamot, D.M.; Stegeman, J.A.; Rutten, V.P.; Jansen, C.A. Glucose Oligosaccharide and Long-Chain Glucomannan Feed Additives Induce Enhanced Activation of Intraepithelial NK Cells and Relative Abundance of Commensal Lactic Acid Bacteria in Broiler Chickens. Vet. Sci. 2021, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Teng, P.Y.; Lee, T.T.; Yu, B. Effects of multi-strain probiotic supplementation on intestinal microbiota, tight junctions, and inflammation in young broiler chickens challenged with Salmonella enterica subsp. enterica. Asian Australas. J. Anim. Sci. 2020, 33, 1797. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cai, K.; Mishra, R.; Jha, R. In ovo supplementation of chitooligosaccharide and chlorella polysaccharide affects cecal microbial community, metabolic pathways, and fermentation metabolites in broiler chickens. Poult. Sci. 2020, 99, 4776–4785. [Google Scholar] [CrossRef] [PubMed]
- Stefaniak, T.; Madej, J.P.; Graczyk, S.; Siwek, M.; Łukaszewicz, E.; Kowalczyk, A.; Sieńczyk, M.; Bednarczyk, M. Selected prebiotics and synbiotics administered in ovo can modify innate immunity in chicken broilers. BMC Vet. Res. 2019, 15, 1–9. [Google Scholar] [CrossRef]
- Nash, T.J.; Morris, K.M.; Mabbott, N.A.; Vervelde, L. Inside-out chicken enteroids with leukocyte component as a model to study host–pathogen interactions. Commun. Biol. 2021, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Oost, M.J.; Velkers, F.C.; Kraneveld, A.D.; Venema, K. Development of the in vitro Cecal Chicken ALIMEntary tRact mOdel-2 to Study Microbiota Composition and Function. Front. Microbiol. 2021, 12, 726447. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Schematic representation of the chicken gastrointestinal tract, including the avian specific crop, gizzard, ceca, and cloaca; (b) Anatomical features of chicken intestinal epithelium and innate immune cells embedded in intestinal epithelium, lamina propria, and Peyer’s patches; (c) Cross-talk between host innate immune cells and Salmonella serovars.
Figure 1.
(a) Schematic representation of the chicken gastrointestinal tract, including the avian specific crop, gizzard, ceca, and cloaca; (b) Anatomical features of chicken intestinal epithelium and innate immune cells embedded in intestinal epithelium, lamina propria, and Peyer’s patches; (c) Cross-talk between host innate immune cells and Salmonella serovars.


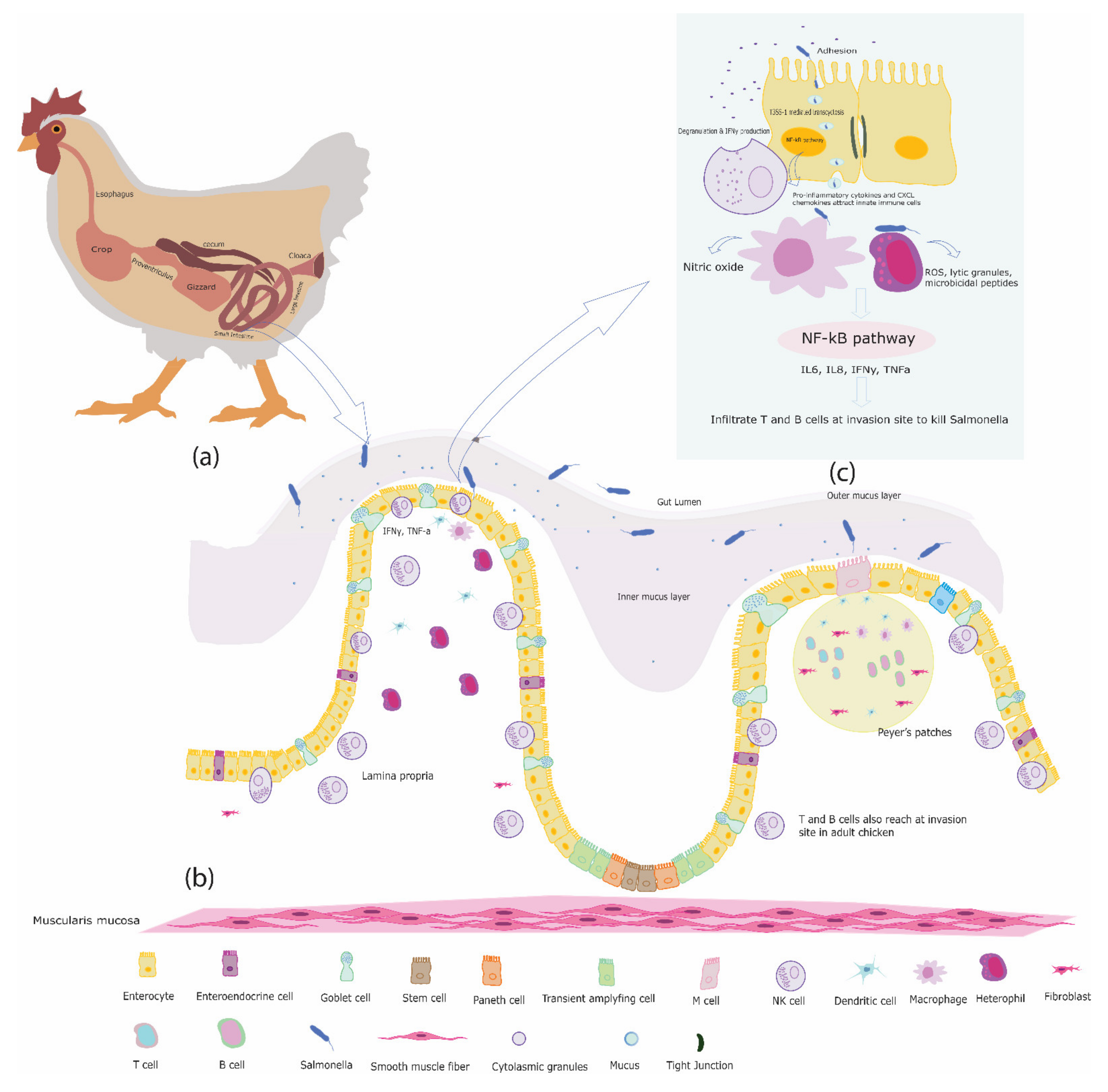{kind=link}
Table 1.
Overview of functions performed by intestinal innate immune cells against invading pathogen.
Table 1.
Overview of functions performed by intestinal innate immune cells against invading pathogen.
| Host Cell | Mediators | Functions | References |
|---|---|---|---|
| Intestinal epithelial cells | anti-microbial peptides (AMPs), Mucus, CXC chemokines, IL12, IL18, IL1β | Provide diffusive barrier to pathogen; Attract underlying innate immune cells; shape the activity of the immune cells | [173,174,175,176] |
| NK cells | perforins, granzymes, IFNγ, TNFα | Cytolysis of transformed and infected host cells; shape the activity of innate and adaptive immune cells | [79,84] |
| Heterophils | CXCLi, IL6, IL1β, cytoplasmic granules, HET | Direct Killing of invading pathogen by phagocytosis, degranulation, HET; Attract immune cells from peripheral blood and nearby organs | [56,97,98] |
| DCs | CXCLi1, CXCLi2, IL1β, IL10, IL6, IL12p35, TNFα | Antigen presentation to adaptive immune cells to initiate specific immune response; attract and shape the function of innate immune cells | [121] |
| Macrophages | CXCLi1, CXCLi2, IL10, IL12p40, IL12, IL18, TNFα, iNOS | Direct killing of pathogen by producing reactive oxygen species, nitric oxide production; Drive Th1 adaptive immune response | [146,149,152,155,157] |
| γδ T cells | Perforins, granzymes, IL17, IFNγ | Cytolysis of infected host cells; Attract other innate immune cells at site of infection and shape their immune function | [170,171] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ijaz, A.; Veldhuizen, E.J.A.; Broere, F.; Rutten, V.P.M.G.; Jansen, C.A. The Interplay between Salmonella and Intestinal Innate Immune Cells in Chickens. Pathogens 2021, 10, 1512. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111512
AMA Style
Ijaz A, Veldhuizen EJA, Broere F, Rutten VPMG, Jansen CA. The Interplay between Salmonella and Intestinal Innate Immune Cells in Chickens. Pathogens. 2021; 10(11):1512. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111512
Chicago/Turabian StyleIjaz, Adil, Edwin J. A. Veldhuizen, Femke Broere, Victor P. M. G. Rutten, and Christine A. Jansen. 2021. "The Interplay between Salmonella and Intestinal Innate Immune Cells in Chickens" Pathogens 10, no. 11: 1512. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111512
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.