
Identification and Comparison of Colletotrichum Secreted Effector Candidates Reveal Two Independent Lineages Pathogenic to Soybean

 , , and
, , and
Abstract
:
1. Introduction
2. Results
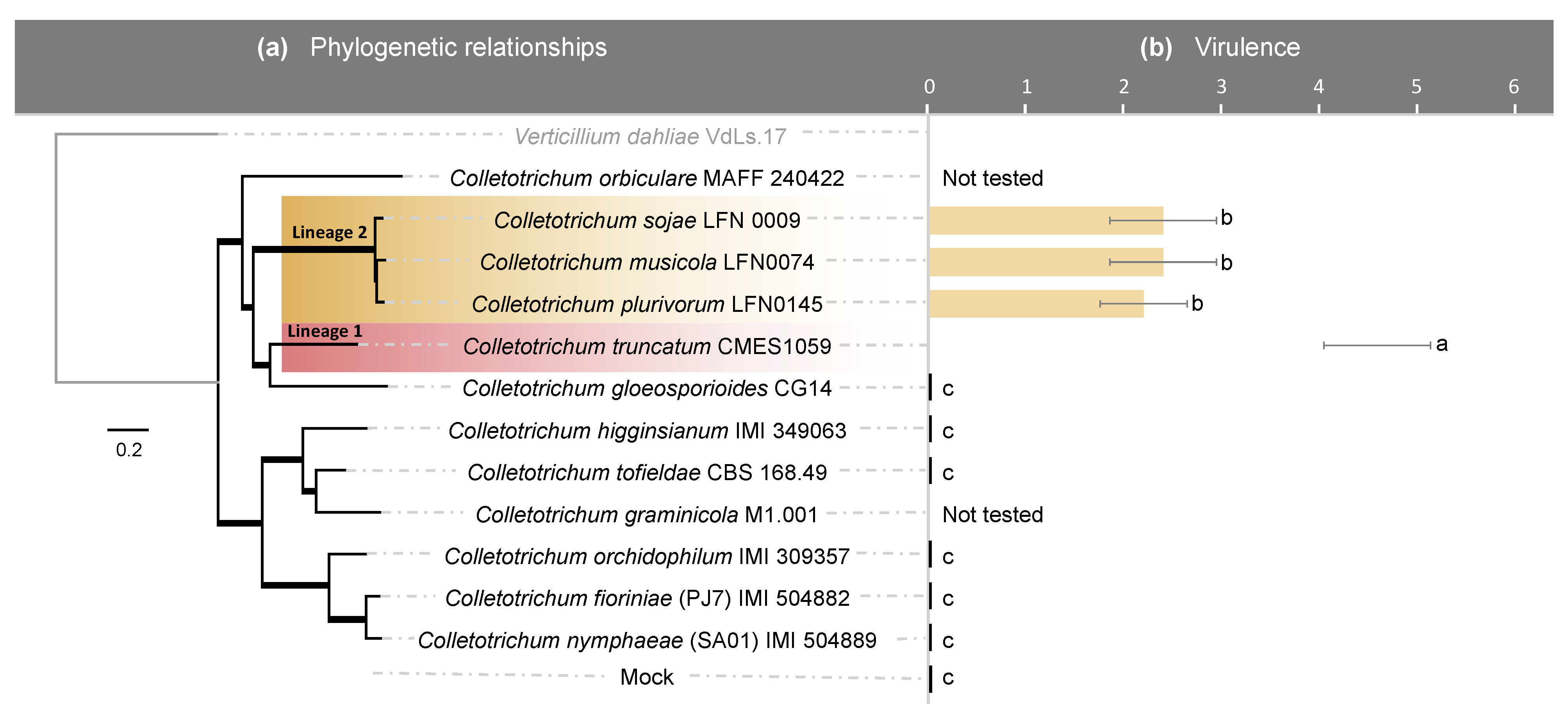
2.1. Among the Selected Colletotrichum Species, Only C. truncatum and Members of the C. orchidearum s.c. Are Pathogenic to Soybean
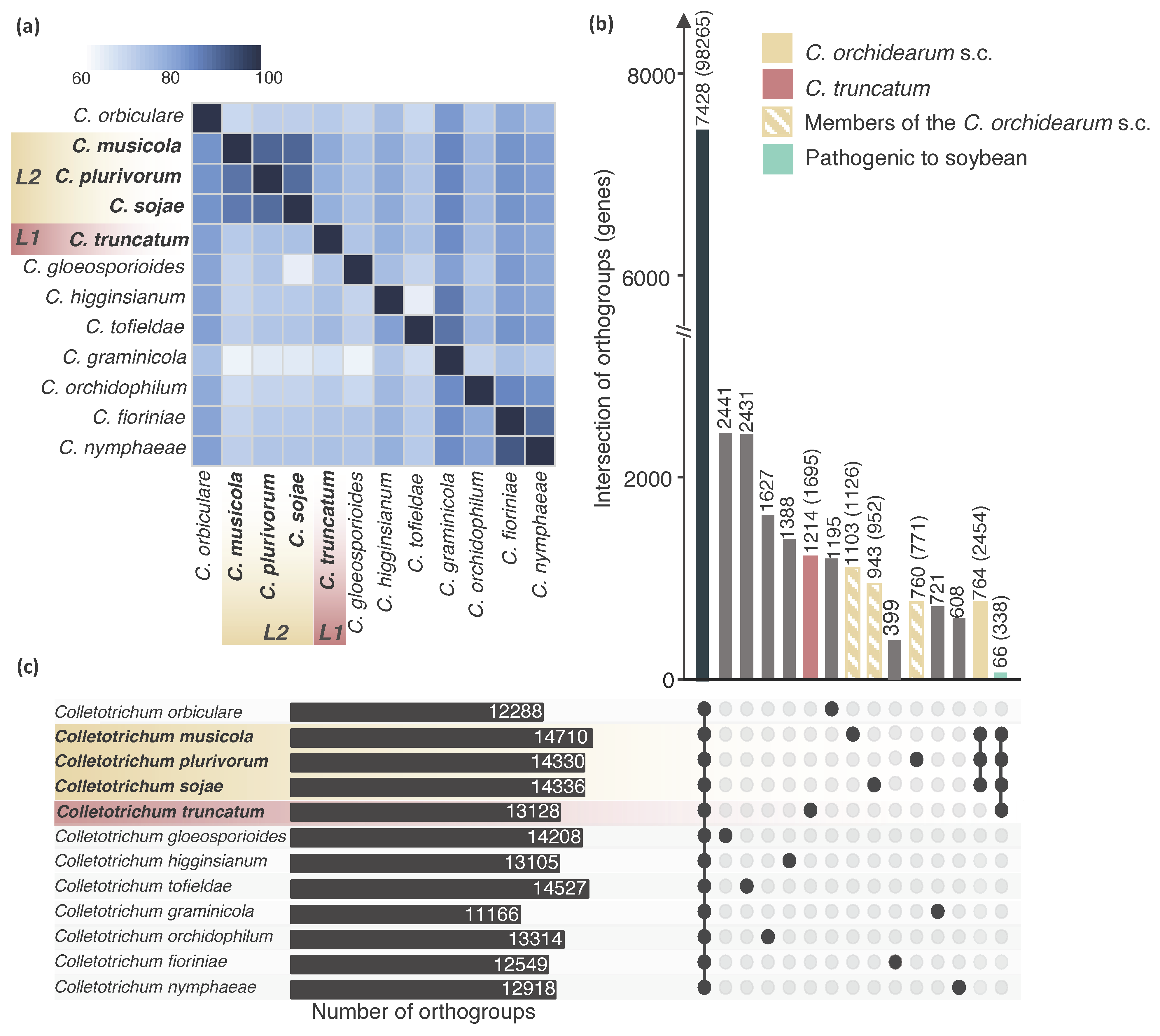
2.2. The Majority of Candidate Effectors of Colletotrichum Species Are Conserved
2.3. C. truncatum LSECs Are Expressed and Have Evolutionary Evidence
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strains Used
5.2. Pathogenicity Assays
5.3. Identification of Specific Effector Protein Candidates (SECs) of Soybean Pathogenic Colletotrichum Species
5.4. Genome Assembly and Gene Evolution
5.5. Evidence of Expression of C. truncatum by RNAseq
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops That Feed the World 2. Soybean—Worldwide Production. Food Sec. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Boufleur, T.R.; Ciampi-Guillardi, M.; Tikami, I.; Rogério, F.; Thon, M.R.; Sukno, S.A.; Massola Júnior, N.S.; Baroncelli, R. Soybean Anthracnose Caused by Colletotrichum Species: Curr. Status and Future Prospects. Mol. Plant Pathol. 2021. [Google Scholar] [CrossRef]
- Dias, M.D.; Pinheiro, V.F.; Café-Filho, A.C. Impact of Anthracnose on the Yield of Soybean Subjected to Chemical Control in the North Region of Brazil. Summa Phytopathol. 2016, 42, 18–23. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Mol. Plant Pathol.: Top 10 Fungal Pathogens. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Talhinhas, P.; Baroncelli, R. Colletotrichum Species and Complexes: Geographic Distribution, Host Range and Conservation Status. Fungal Divers. 2021, 110, 109–198. [Google Scholar] [CrossRef]
- Nakata, K.; Takimoto, K. A List of Crop Diseases in Korea; Agricultural experiment station governmental central chosen research report; Korean Society of Plant Pathology: Seoul, Korea, 1934; pp. 1–146. [Google Scholar]
- Chen, S.; Wang, Y.; Schnabel, G.; Peng, C.A.; Lagishetty, S.; Smith, K.; Luo, C.; Yuan, H. Inherent Resistance to 14α-Demethylation Inhibitor Fungicides in Colletotrichum truncatum Is Likely Linked to CYP51A and/or CYP51B Gene Variants. Phytopathology 2018, 108, 1263–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogério, F.; Ciampi-Guillardi, M.; Barbieri, M.C.G.; Bragança, C.A.D.; Seixas, C.D.S.; Almeida, A.M.R.; Massola, N.S. Phylogeny and Variability of Colletotrichum truncatum Associated with Soybean Anthracnose in Brazil. J. Appl. Microbiol. 2017, 122, 402–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciampi-Guillardi, M.; Ramiro, J.; de Moraes, M.H.D.; Barbieri, M.C.G.; Massola, N.S. Multiplex QPCR Assay for Direct Detection and Quantification of Colletotrichum truncatum, Corynespora cassiicola, and Sclerotinia sclerotiorum in Soybean Seeds. Plant Dis. 2020, 104, 3002–3009. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum Species with Curved Conidia from Herbaceous Hosts. Fungal Divers. 2009, 39, 45–87. [Google Scholar]
- Dias, M.D.; Dias-Neto, J.J.; Santos, M.D.M.; Formento, A.N.; Bizerra, L.V.A.S.; Fonseca, M.E.N.; Boiteux, L.S.; Café-Filho, A.C. Curr. Status of Soybean Anthracnose Associated with Colletotrichum truncatum in Brazil and Argentina. Plants 2019, 8, 459. [Google Scholar] [CrossRef] [Green Version]
- Nataraj, V.; Maranna, S.; Kumawat, G.; Gupta, S.; Rajput, L.S.; Kumar, S.; Sharma, A.N.; Bhatia, V.S. Genetic Inheritance and Identification of Germplasm Sources for Anthracnose Resistance in Soybean [Glycine max (L.) Merr.]. Genet. Resour. Crop Evol. 2020, 67, 1449–1456. [Google Scholar] [CrossRef]
- Poti, T.; Mahawan, K.; Cheewangkoon, R.; Arunothayanan, H.; Akimitsu, K.; Nalumpang, S. Detection and Mol. Characterization of Carbendazim-Resistant Colletotrichum truncatum Isolates Causing Anthracnose of Soybean in Thailand. J. Phytopathol. 2020, 168, 267–278. [Google Scholar] [CrossRef]
- Rogério, F.; Gladieux, P.; Massola, N.S.; Ciampi-Guillardi, M. Multiple Introductions without Admixture of Colletotrichum truncatum Associated with Soybean Anthracnose in Brazil. Phytopathology 2019, 109, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.-C.; Haudenshield, J.S.; Hartman, G.L. Multiplex Real-Time PCR Detection and Differentiation of Colletotrichum Species Infecting Soybean. Plant Dis. 2015, 99, 1559–1568. [Google Scholar] [CrossRef] [Green Version]
- Boufleur, T.R.; Castro, R.R.L.; Rogério, F.; Ciampi-Guillardi, M.; Baroncelli, R.; Massola Júnior, N.S. First Report of Colletotrichum musicola Causing Soybean Anthracnose in Brazil. Plant Dis. 2020, 104, 1858. [Google Scholar] [CrossRef]
- Barbieri, M.C.G.; Ciampi-Guillardi, M.; Moraes, S.R.G.; Bonaldo, S.M.; Rogério, F.; Linhares, R.R.; Massola, N.S. First Report of Colletotrichum cliviae Causing Anthracnose on Soybean in Brazil. Plant Dis. 2017, 101, 1677. [Google Scholar] [CrossRef]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum Species Complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.-C.; Wang, S.-Y.; Duan, X.-C.; Gao, X.; Zhu, X.-Y.; Laborda, P. First Report of Colletotrichum Brevisporum Causing Soybean Anthracnose in China. Plant Dis. 2020, 105, 707. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Stam, R.; Cano, L.M.; Song, J.; Sklenar, J.; Yoshida, K.; Bozkurt, T.O.; Oliva, R.; Liu, Z.; Tian, M.; et al. Effector Specialization in a Lineage of the Irish Potato Famine Pathogen. Science 2014, 343, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Raffaele, S.; Kamoun, S. The Two-Speed Genomes of Filamentous Pathogens: Waltz with Plants. Curr. Opin. Genet. Devel. 2015, 35, 57–65. [Google Scholar] [CrossRef]
- Win, J.; Chaparro-Garcia, A.; Belhaj, K.; Saunders, D.G.O.; Yoshida, K.; Dong, S.; Schornack, S.; Zipfel, C.; Robatzek, S.; Hogenhout, S.A.; et al. Effector Biol. of Plant-Associated Organisms: Concepts and Perspectives. Cold Spring Harbor Symp. Quant. Biol. 2012, 77, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Kanyuka, K.; Rudd, J.J. Cell Surface Immune Receptors: The Guardians of the Plant’s Extracellular Spaces. Curr. Opin. Plant Biol. 2019, 50, 1–8. [Google Scholar] [CrossRef]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-Recognition Receptors Are Required for NLR-Mediated Plant Immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D.G. Mutual Potentiation of Plant Immunity by Cell-Surface and Intracellular Receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef]
- Bjornson, M.; Zipfel, C. Plant Immunity: Crosstalk between Plant Immune Receptors. Curr. Biol. 2021, 31, R796–R798. [Google Scholar] [CrossRef]
- Zhan, J.; Thrall, P.H.; Burdon, J.J. Achieving Sustainable Plant Dis. Management through Evolutionary Principles. Trends Plant Sci. 2014, 19, 570–575. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Kamoun, S. Genome Evolution in Filamentous Plant Pathogens: Why Bigger Can Be Better. Nat. Rev. Microbiol. 2012, 10, 417–430. [Google Scholar] [CrossRef]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal Effectors and Plant Susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Plissonneau, C.; Benevenuto, J.; Mohd-Assaad, N.; Fouché, S.; Hartmann, F.E.; Croll, D. Using Population and Comparative Genomics to Understand the Genetic Basis of Effector-Driven Fungal Pathogen Evolution. Front. Plant Sci. 2017, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle Transitions in Plant Pathogenic Colletotrichum Fungi Deciphered by Genome and Transcriptome Analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Gan, P.; Ikeda, K.; Irieda, H.; Narusaka, M.; O’Connell, R.J.; Narusaka, Y.; Takano, Y.; Kubo, Y.; Shirasu, K. Comparative Genomic and Transcriptomic Analyses Reveal the Hemibiotrophic Stage Shift of Colletotrichum Fungi. New Phytol. 2013, 197, 1236–1249. [Google Scholar] [CrossRef] [PubMed]
- Alkan, N.; Friedlander, G.; Ment, D.; Prusky, D.; Fluhr, R. Simultaneous Transcriptome Analysis of Colletotrichum gloeosporioides and Tomato Fruit Pathosystem Reveals Novel Fungal Pathogenicity and Fruit Defense Strategies. New Phytol. 2015, 205, 801–815. [Google Scholar] [CrossRef]
- Huo, J.; Wang, Y.; Hao, Y.; Yao, Y.; Wang, Y.; Zhang, K.; Tan, X.; Li, Z.; Wang, W. Genome Sequence Resource for Colletotrichum scovillei, the Cause of Anthracnose Disease of Chili. MPMI 2021, 34, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Eaton, M.J.; Edwards, S.; Inocencio, H.A.; Machado, F.J.; Nuckles, E.M.; Farman, M.; Gauthier, N.A.; Vaillancourt, L.J. Diversity and Cross-Infection Potential of Colletotrichum Causing Fruit Rots in Mixed-Fruit Orchards in Kentucky. Plant Dis. 2021, 105, 1115–1128. [Google Scholar] [CrossRef]
- Liu, X.; Li, B.; Yang, Y.; Cai, J.; Shi, T.; Zheng, X.; Huang, G. Pathogenic Adaptations Revealed by Comparative Genome Analyses of Two Colletotrichum Spp., the Causal Agent of Anthracnose in Rubber Tree. Front. Microbiol. 2020, 11, 1484. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, R.; Sreenivasaprasad, S.; Sukno, S.A.; Thon, M.R.; Holub, E. Draft Genome Sequence of Colletotrichum scutatum sensu lato (Colletotrichum fioriniae). Genome Announc. 2014, 2, e00112-14. [Google Scholar] [CrossRef] [Green Version]
- Baroncelli, R.; Amby, D.B.; Zapparata, A.; Sarrocco, S.; Vannacci, G.; Le Floch, G.; Harrison, R.J.; Holub, E.; Sukno, S.A.; Sreenivasaprasad, S.; et al. Gene Family Expansions and Contractions Are Associated with Host Range in Plant Pathogens of the Genus Colletotrichum. BMC Genom. 2016, 17, 555. [Google Scholar] [CrossRef] [Green Version]
- Gan, P.; Narusaka, M.; Kumakura, N.; Tsushima, A.; Takano, Y.; Narusaka, Y.; Shirasu, K. Genus-Wide Comparative Genome Analyses of Colletotrichum Species Reveal Specific Gene Family Losses and Gains during Adaptation to Specific Infection Lifestyles. Genome Biol. Evol. 2016, 8, 1467–1481. [Google Scholar] [CrossRef] [Green Version]
- Hacquard, S.; Kracher, B.; Hiruma, K.; Münch, P.C.; Garrido-Oter, R.; Thon, M.R.; Weimann, A.; Damm, U.; Dallery, J.-F.; Hainaut, M.; et al. Survival Trade-Offs in Plant Roots during Colonization by Closely Related Beneficial and Pathogenic Fungi. Nat. Commun. 2016, 7, 11362. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, R.; Prasanth, C.N.; Malathi, P.; Sundar, A.R. Draft Genome Sequence of Colletotrichum Falcatum-a Prelude on Screening of Red Rot Pathogen in Sugarcane. J. Genom. 2016, 4, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-H.; Chon, J.-K.; Ahn, J.-H.; Choi, I.-Y.; Lee, Y.-H.; Kim, K.S. Whole Genome Sequence and Genome Annotation of Colletotrichum Acutatum, Causal Agent of Anthracnose in Pepper Plants in South Korea. Genom. Data 2016, 8, 45–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallery, J.-F.; Lapalu, N.; Zampounis, A.; Pigné, S.; Luyten, I.; Amselem, J.; Wittenberg, A.H.J.; Zhou, S.; de Queiroz, M.V.; Robin, G.P.; et al. Gapless Genome Assembly of Colletotrichum higginsianum Reveals Chromosome Structure and Association of Transposable Elements with Secondary Metabolite Gene Clusters. BMC Genom. 2017, 18, 667. [Google Scholar] [CrossRef]
- Baroncelli, R.; Sukno, S.A.; Sarrocco, S.; Cafà, G.; Le Floch, G.; Thon, M.R. Whole-Genome Sequence of the Orchid Anthracnose Pathogen Colletotrichum orchidophilum. MPMI 2018, 31, 979–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, P.; Narusaka, M.; Tsushima, A.; Narusaka, Y.; Takano, Y.; Shirasu, K. Draft Genome Assembly of Colletotrichum chlorophyti, a Pathogen of Ehbaceous Plants. Genome Announc. 2017, 5, e01733-16. [Google Scholar] [CrossRef] [Green Version]
- Gan, P.; Tsushima, A.; Narusaka, M.; Narusaka, Y.; Takano, Y.; Kubo, Y.; Shirasu, K. Genome Sequence Resources for Four Phytopathogenic Fungi from the Colletotrichum orbiculare Species Complex. MPMI 2019, 32, 1088–1090. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Fujikawa, T.; Nakamori, D.; Iwai, H. Draft Genome Sequence of Colletotrichum sansevieriae Sa-1–2, the Anthracnose Pathogen of Sansevieria trifasciata. Data Brief 2018, 18, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Bhadauria, V.; MacLachlan, R.; Pozniak, C.; Cohen-Skalie, A.; Li, L.; Halliday, J.; Banniza, S. Genetic Map-Guided Genome Assembly Reveals a Virulence-Governing Minichromosome in the Lentil Anthracnose Pathogen Colletotrichum lentis. New Phytol. 2019, 221, 431–445. [Google Scholar] [CrossRef] [Green Version]
- da Silva Junior, W.J.; Falcão, R.M.; de Sousa-Paula, L.C.; Sbaraini, N.; dos Santos Vieira, W.A.; Lima, W.G.; Junior, S.D.S.L.P.; Staats, C.C.; Schrank, A.; Benko-Iseppon, A.M.; et al. Draft Genome Assembly of Colletotrichum musae, the Pathogen of Banana Fruit. Data Brief 2018, 17, 256–260. [Google Scholar] [CrossRef]
- Lelwala, R.V.; Korhonen, P.K.; Young, N.D.; Scott, J.B.; Ades, P.K.; Gasser, R.B.; Taylor, P.W.J. Comparative Genome Analysis Indicates High Evolutionary Potential of Pathogenicity Genes in Colletotrichum tanaceti. PLoS ONE 2019, 14, e0212248. [Google Scholar] [CrossRef]
- Gan, P.; Tsushima, A.; Hiroyama, R.; Narusaka, M.; Takano, Y.; Narusaka, Y.; Kawaradani, M.; Damm, U.; Shirasu, K. Colletotrichum shisoi Sp. Nov., an Anthracnose Pathogen of Perilla frutescens in Japan: Mol. Phylogenetic, Morphological and Genomic Evidence. Sci. Rep. 2019, 9, 13349. [Google Scholar] [CrossRef]
- Gan, P.; Hiroyama, R.; Tsushima, A.; Masuda, S.; Shibata, A.; Ueno, A.; Kumakura, N.; Narusaka, M.; Hoat, T.X.; Narusaka, Y.; et al. Telomeres and a Repeat-rich Chromosome Encode Effector Gene Clusters in Plant Pathogenic Colletotrichum Fungi. Environ. Microbiol. 2021, 23, 6004–6018. [Google Scholar] [CrossRef]
- Rogério, F.; Boufleur, T.R.; Ciampi-Guillardi, M.; Sukno, S.A.; Thon, M.R.; Massola Júnior, N.S.; Baroncelli, R. Genome Sequence Resources of Colletotrichum truncatum, C. Plurivorum, C. musicola, and C. sojae: Four Species Pathogenic to Soybean (Glycine max). Phytopathology 2020, 110, 1497–1499. [Google Scholar] [CrossRef]
- Meng, Y.; Ren, Y.; Wang, W.; Gleason, M.L.; Zhang, R.; Sun, G. A Genome Sequence Resource for the Geographically Widespread Anthracnose Pathogen Colletotrichum asianum. Plant Dis. 2020, 104, 2044–2047. [Google Scholar] [CrossRef] [Green Version]
- Baroncelli, R.; Pensec, F.; Da Lio, D.; Boufleur, T.R.; Vicente, I.; Sarrocco, S.; Picot, A.; Baraldi, E.; Sukno, S.; Thon, M.R.; et al. Complete Genome Sequence of the Plant Pathogenic Fungus Colletotrichum lupini. MPMI 2021. [Google Scholar] [CrossRef]
- Ma, L.-J.; van der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.-J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.; et al. Comparative Genomics Reveals Mobile Pathogenicity Chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Poppe, S.; Dorsheimer, L.; Happel, P.; Stukenbrock, E.H. Rapidly Evolving Genes Are Key Players in Host Specialization and Virulence of the Fungal Wheat Pathogen Zymoseptoria tritici (Mycosphaerella graminicola). PLoS Pathog. 2015, 11, e1005055. [Google Scholar] [CrossRef]
- Petre, B.; Lorrain, C.; Stukenbrock, E.H.; Duplessis, S. Host-Specialized Transcriptome of Plant-Associated Organisms. Curr. Opin. Plant Biol. 2020, 56, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Palma-Guerrero, J.; Ma, X.; Torriani, S.F.F.; Zala, M.; Francisco, C.S.; Hartmann, F.E.; Croll, D.; McDonald, B.A. Comparative Transcriptome Analyses in Zymoseptoria tritici Reveal Significant Differences in Gene Expression among Strains during Plant Infection. MPMI 2017, 30, 231–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, J.; Huang, H.; Meusnier, I.; Adreit, H.; Ducasse, A.; Bonnot, F.; Pan, L.; He, X.; Kroj, T.; Fournier, E.; et al. Pathogen Effectors and Plant Immunity Determine Specialization of the Blast Fungus to Rice Subspecies. eLife 2016, 5, e19377. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vallet, A.; Fouché, S.; Fudal, I.; Hartmann, F.E.; Soyer, J.L.; Tellier, A.; Croll, D. The Genome Biol. of Effector Gene Evolution in Filamentous Plant Pathogens. Annu. Rev. Phytopathol. 2018, 56, 21–40. [Google Scholar] [CrossRef]
- Kleemann, J.; Rincon-Rivera, L.J.; Takahara, H.; Neumann, U.; Ver Loren van Themaat, E.; van der Does, H.C.; Hacquard, S.; Stüber, K.; Will, I.; Schmalenbach, W.; et al. Correction: Sequential Delivery of Host-Induced Virulence Effectors by Appressoria and Intracellular Hyphae of the Phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar] [CrossRef]
- Robin, G.P.; Kleemann, J.; Neumann, U.; Cabre, L.; Dallery, J.-F.; Lapalu, N.; O’Connell, R.J. Subcellular Localization Screening of Colletotrichum higginsianum Effector Candidates Identifies Fungal Proteins Targeted to Plant Peroxisomes, Golgi Bodies, and Microtubules. Front. Plant Sci. 2018, 9, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irieda, H.; Maeda, H.; Akiyama, K.; Hagiwara, A.; Saitoh, H.; Uemura, A.; Terauchi, R.; Takano, Y. Colletotrichum orbiculare Secretes Virulence Effectors to a Biotrophic Interface at the Primary Hyphal Neck via Exocytosis Coupled with SEC22-Mediated Traffic. Plant Cell 2014, 26, 2265–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irieda, H.; Takano, Y. Identification and Characterization of Virulence-Related Effectors in the Cucumber Anthracnose Fungus Colletotrichum orbiculare. Physiol. Mol. Plant Pathol. 2016, 95, 87–92. [Google Scholar] [CrossRef]
- Bhadauria, V.; MacLachlan, R.; Pozniak, C.; Banniza, S. Candidate Effectors Contribute to Race Differentiation and Virulence of the Lentil Anthracnose Pathogen Colletotrichum lentis. BMC Genom. 2015, 16, 628. [Google Scholar] [CrossRef] [Green Version]
- Bhadauria, V.; Banniza, S.; Vandenberg, A.; Selvaraj, G.; Wei, Y. Overexpression of a Novel Biotrophy-Specific Colletotrichum truncatum Effector, CtNUDIX, in Hemibiotrophic Fungal Phytopathogens Causes Incompatibility with Their Host Plants. Eukar. Cell 2013, 12, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Bhadauria, V.; Vijayan, P.; Wei, Y.; Banniza, S. Transcriptome Analysis Reveals a Complex Interplay between Resistance and Effector Genes during the Compatible Lentil- Colletotrichum lentis Interaction. Sci. Reports 2017, 7, 42338. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Martín, J.M.; Pacheco-Arjona, J.R.; Bello-Rico, V.; Vargas, W.A.; Monod, M.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. A Highly Conserved Metalloprotease Effector Enhances Virulence in the Maize Anthracnose Fungus Colletotrichum graminicola. Mol. Plant Pathol. 2016, 17, 1048–1062. [Google Scholar] [CrossRef] [Green Version]
- Vargas, W.A.; Sanz-Martín, J.M.; Rech, G.E.; Armijos-Jaramillo, V.D.; Rivera, L.P.; Echeverria, M.M.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. A Fungal Effector with Host Nuclear Localization and DNA-Binding Properties Is Required for Maize Anthracnose Development. MPMI 2015, 29, 83–95. [Google Scholar] [CrossRef]
- Eisermann, I.; Weihmann, F.; Krijger, J.-J.; Kröling, C.; Hause, G.; Menzel, M.; Pienkny, S.; Kiesow, A.; Deising, H.B.; Wirsel, S.G.R. Two Genes in a Pathogenicity Gene Cluster Encoding Secreted Proteins Are Required for Appressorial Penetration and Infection of the Maize Anthracnose Fungus Colletotrichum graminicola. Environ. Microbiol. 2019, 21, 4773–4791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Queiroz, C.B.; Correia, H.L.N.; Santana, M.F.; Batista, D.S.; Vidigal, P.M.P.; Brommonschenkel, S.H.; de Queiroz, M.V. The Repertoire of Effector Candidates in Colletotrichum lindemuthianum Reveals Important Information about Colletotrichum Genus Lifestyle. Appl. Microbiol. Biotechnol. 2019, 103, 2295–2309. [Google Scholar] [CrossRef]
- Prasanth, C.N.; Viswanathan, R.; Malathi, P.; Sundar, A.R. Comparative Transcriptome Analysis of Candidate Secretory Effector Proteins from Colletotrichum falcatum Infecting Sugarcane. Agri Gene 2019, 13, 100089. [Google Scholar] [CrossRef]
- Zampounis, A.; Pigné, S.; Dallery, J.-F.; Wittenberg, A.H.J.; Zhou, S.; Schwartz, D.C.; Thon, M.R.; O’Connell, R.J. Genome Sequence and Annotation of Colletotrichum higginsianum, a Causal Agent of Crucifer Anthracnose Disease. Genome Announc. 2016, 4, e00821-16. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Mol. Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depotter, J.R.L.; Doehlemann, G. Target the Core: Durable Plant Resistance against Filamentous Plant Pathogens through Effector Recognition. Pest. Manag. Sci. 2020, 76, 426–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Edwards, M.C. Genome-Wide Analysis of Small Secreted Cysteine-Rich Proteins Identifies Candidate Effector Proteins Potentially Involved in Fusarium graminearum −wheat Interactions. Phytopathology 2016, 106, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Mesarich, C.H.; Bowen, J.K.; Hamiaux, C.; Templeton, M.D. Repeat-Containing Protein Effectors of Plant-Associated Organisms. Front. Plant Sci. 2015, 6, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperschneider, J.; Catanzariti, A.-M.; DeBoer, K.; Petre, B.; Gardiner, D.M.; Singh, K.B.; Dodds, P.N.; Taylor, J.M. LOCALIZER: Subcellular Localization Prediction of Both Plant and Effector Proteins in the Plant Cell. Sci. Rep. 2017, 7, 44598. [Google Scholar] [CrossRef] [Green Version]
- Sperschneider, J.; Dodds, P.N.; Singh, K.B.; Taylor, J.M. ApoplastP: Prediction of Effectors and Plant Proteins in the Apoplast Using Machine Learning. New Phytol. 2018, 217, 1764–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogenhout, S.A.; Van der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging Concepts in Effector Biology of Plant-Associated Organisms. MPMI 2009, 22, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, I.; van den Burg, H.A.; Okmen, B.; Beenen, H.G.; van Liere, S.; Kema, G.H.J.; de Wit, P.J.G.M. Tomato Cf Resistance Proteins Mediate Recognition of Cognate Homologous Effectors from Fungi Pathogenic on Dicots and Monocots. PNAS 2010, 107, 7610–7615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stergiopoulos, I.; Kourmpetis, Y.A.I.; Slot, J.C.; Bakker, F.T.; De Wit, P.J.G.M.; Rokas, A. In Silico Characterization and Molecular Evolutionary Analysis of a Novel Superfamily of Fungal Effector Proteins. Mol. Biol. Evol. 2012, 29, 3371–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokchorchik, M.; Won, K.; Lee, Y.; Segonzac, C.; Sohn, K.H. Whole Genome Enabled Phylogenetic and Secretome Analyses of Two Venturia nashicola Isolates. Plant Pathol. J. 2020, 36, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Fourie, A.; de Jonge, R.; van der Nest, M.A.; Duong, T.A.; Wingfield, M.J.; Wingfield, B.D.; Barnes, I. Genome Comparisons Suggest an Association between Ceratocystis Host Adaptations and Effector Clusters in Unique Transposable Element Families. Fungal Genet. Biol. 2020, 143, 103433. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.D.; Fonseca, M.E.N.; Dias-Neto, J.J.; Santos, M.D.M.; Pandolfo, G.M.; Boiteux, L.S.; Café-Filho, A.C. Biology, Pathogenicity, and Haplotype Analyses of Colletotrichum cliviae: A Novel Soybean Anthracnose Agent in Warm Tropical Areas. Trop. Plant Pathol. 2018, 43, 439–451. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating Species Boundaries in Colletotrichum. Fungal Divers. 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Hymowitz, T. On the Domestication of the Soybean. Econ. Bot. 1970, 24, 408–421. [Google Scholar] [CrossRef]
- Sharma, S.K.; Gupta, G.K.; Ramteke, R. Colletotrichum truncatum [(Schw.) Andrus & W.D. Moore], the Causal Agent of Anthracnose of Soybean [Glycine max (L.) Merrill.]–a Review. Soybean Res. 2011, 9, 31–52. [Google Scholar]
- Fouché, S.; Plissonneau, C.; Croll, D. The Birth and Death of Effectors in Rapidly Evolving Filamentous Pathogen Genomes. Curr. Opin. Microbiol. 2018, 46, 34–42. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.S.; Waller, J.M.; Abang, M.M.; Zhang, J.Z.; Yang, Y.H.; Phoulivong, S.; Liu, Z.Y.; et al. A Polyphasic Approach for Studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Jayawardena, R. Notes on Currently Accepted Species of Colletotrichum. Mycosphere 2016, 7, 1192–1260. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving Fundamental Biases in Whole Genome Comparisons Dramatically Improves Orthogroup Inference Accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubrulle, G.; Picot, A.; Madec, S.; Corre, E.; Pawtowski, A.; Baroncelli, R.; Zivy, M.; Balliau, T.; Le Floch, G.; Pensec, F. Deciphering the Infectious Process of Colletotrichum lupini in Lupin through Transcriptomic and Proteomic Analysis. Microorganisms 2020, 8, 1621. [Google Scholar] [CrossRef]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R Package for the Visualization of Intersecting Sets and Their Properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes11Edited by F. Cohen. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Pierleoni, A.; Martelli, P.; Casadio, R. PredGPI: A GPI-Anchor Predictor. BMC Bioinform. 2008, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Jorda, J.; Kajava, A.V. T-REKS: Identification of Tandem REpeats in Sequences with a K-MeanS Based Algorithm. Bioinformatics 2009, 25, 2632–2638. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Dmitry, A.; Gurevich, A.A.; Dvorkin, M.; Kulikoc, A.S.; Lesin, V.M.; Nikolenki, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
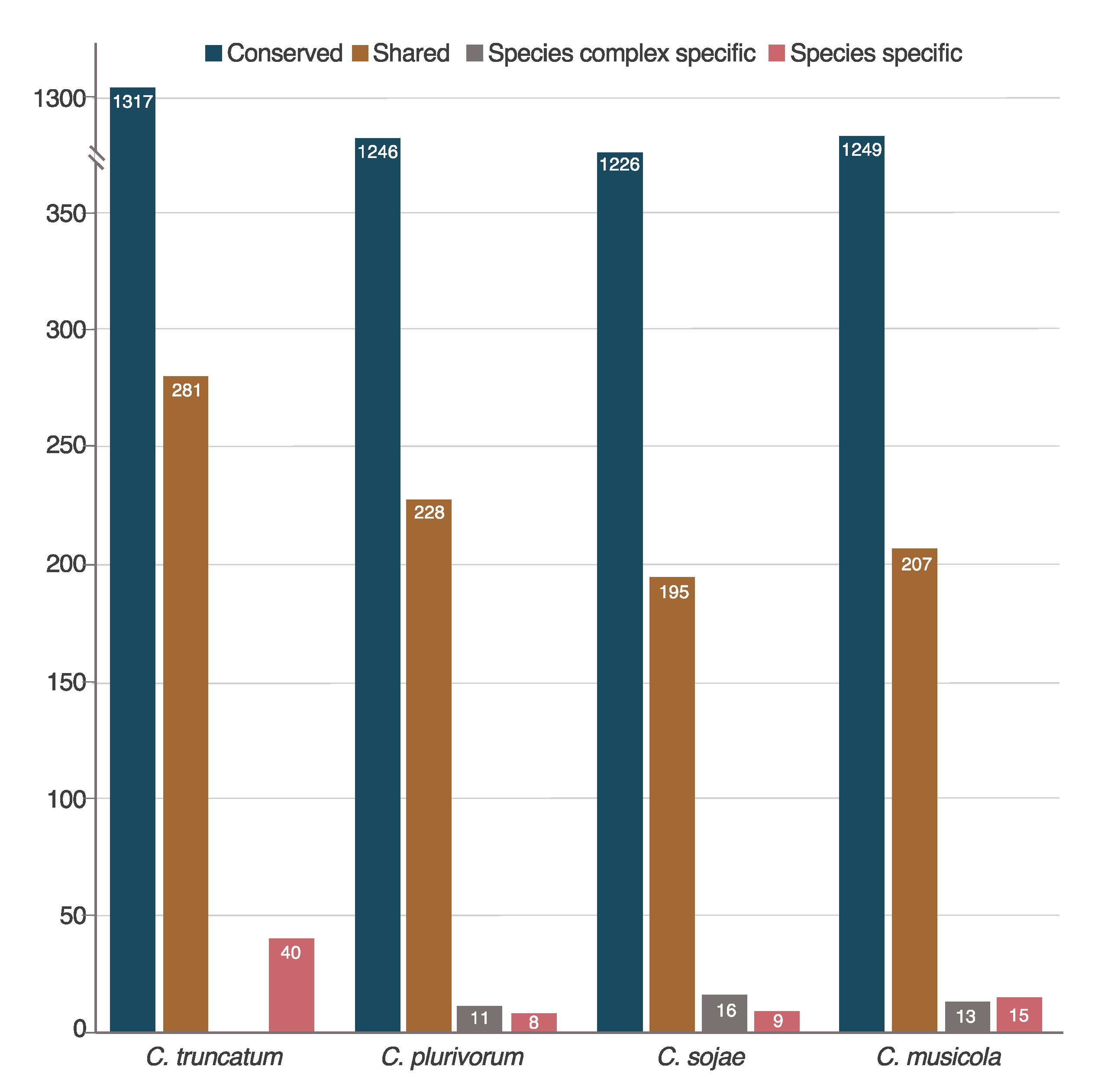{kind=link}
{kind=link}
| Strain | Species | Species Complex | Host | Origin | * |
|---|---|---|---|---|---|
| MAFF 240422 | C. orbiculare | C. orbiculare | Cucumis sativus | Japan | [47] |
| LFN0074 | C. musicola | C. orchidearum | Glycine max | Brazil | [54] |
| LFN0145 | C. plurivorum | C. orchidearum | Glycine max | Brazil | [54] |
| LFN0009 | C. sojae | C. orchidearum | Glycine max | Brazil | [54] |
| CMES1059 | C. truncatum | C. truncatum | Glycine max | Brazil | [54] |
| Cg-14 | C. gloeosporioides s.s. | C. gloeosporioides | Persea americana | Israel | [34] |
| IMI 349063 | C. higginsianum | C. destructivum | Brassica rapa | Trinidad & Tobago | [75] |
| CBS 168.49 | C. tofieldiae | C. spaethianum | Lupinus polyphyllus | Germany | [41] |
| M1.001 | C. graminicola | C. graminicola | Zea mays | USA | [32] |
| IMI 309357 | C. orchidophilum | none | Phalaenopsis sp. | United Kingdom | [45] |
| IMI 504882 | C. fioriniae | C. acutatum | Fragaria x ananassa | New Zealand | [29] |
| IMI 504889 | C. nymphaeae | C. acutatum | Fragaria x ananassa | Denmark | [39] |
| Species | Proteome | Signal Peptide | Absence of TM/GPI Anchor | % of Secreted Proteins |
|---|---|---|---|---|
| C. truncatum | 15,901 | 2116 | 1638 | 10 |
| C. plurivorum | 15,153 | 1989 | 1495 | 10 |
| C. sojae | 16,124 | 1931 | 1447 | 9 |
| C. musicola | 16,826 | 1871 | 1485 | 9 |
| C. orchidearum s.c. SECs | |||||||
|---|---|---|---|---|---|---|---|
| Species | LSECs | RCP | SL (NLS) | SL (Other) | Apoplast | CR | EffectorP |
| C. musicola | 13 | 5 | 0 | 0 | 9 | 7 | 6 |
| C. plurivorum | 11 | 5 | 1 | 0 | 7 | 6 | 5 |
| C. sojae | 16 | 3 | 2 | 2 | 11 | 8 | 5 |
| Species-LSECs | |||||||
| C. truncatum | 40 | 7 | 5 | 2 | 16 | 21 | 16 |
| C. musicola | 15 | 2 | 0 | 2 | 4 | 8 | 5 |
| C. plurivorum | 8 | 0 | 0 | 4 | 1 | 4 | 2 |
| C. sojae | 9 | 2 | 0 | 1 | 2 | 6 | 3 |
| Strain | Species | Species Complex | Host | Origin | Accession N° |
|---|---|---|---|---|---|
| MT1-01 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095338 |
| MT2-05 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095339 |
| MT3-01 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095348 |
| MT3-21 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095349 |
| MT4-05 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095350 |
| MT4-13 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095351 |
| MT5-12 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095352 |
| MT5-26 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095353 |
| MT5-32 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095354 |
| GO2-03 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095355 |
| GO2-06 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095340 |
| GO2-12 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095341 |
| GO4-07 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095342 |
| GO4-08 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095343 |
| GO4-17 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095344 |
| GO5-11 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095345 |
| GO5-14 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095346 |
| GO5-25 | C. truncatum | C. truncatum | Glycine max | Brazil | SRX7095347 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boufleur, T.R.; Massola Júnior, N.S.; Tikami, Í.; Sukno, S.A.; Thon, M.R.; Baroncelli, R. Identification and Comparison of Colletotrichum Secreted Effector Candidates Reveal Two Independent Lineages Pathogenic to Soybean. Pathogens 2021, 10, 1520. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111520
Boufleur TR, Massola Júnior NS, Tikami Í, Sukno SA, Thon MR, Baroncelli R. Identification and Comparison of Colletotrichum Secreted Effector Candidates Reveal Two Independent Lineages Pathogenic to Soybean. Pathogens. 2021; 10(11):1520. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111520
Chicago/Turabian StyleBoufleur, Thaís R., Nelson S. Massola Júnior, Ísis Tikami, Serenella A. Sukno, Michael R. Thon, and Riccardo Baroncelli. 2021. "Identification and Comparison of Colletotrichum Secreted Effector Candidates Reveal Two Independent Lineages Pathogenic to Soybean" Pathogens 10, no. 11: 1520. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111520