
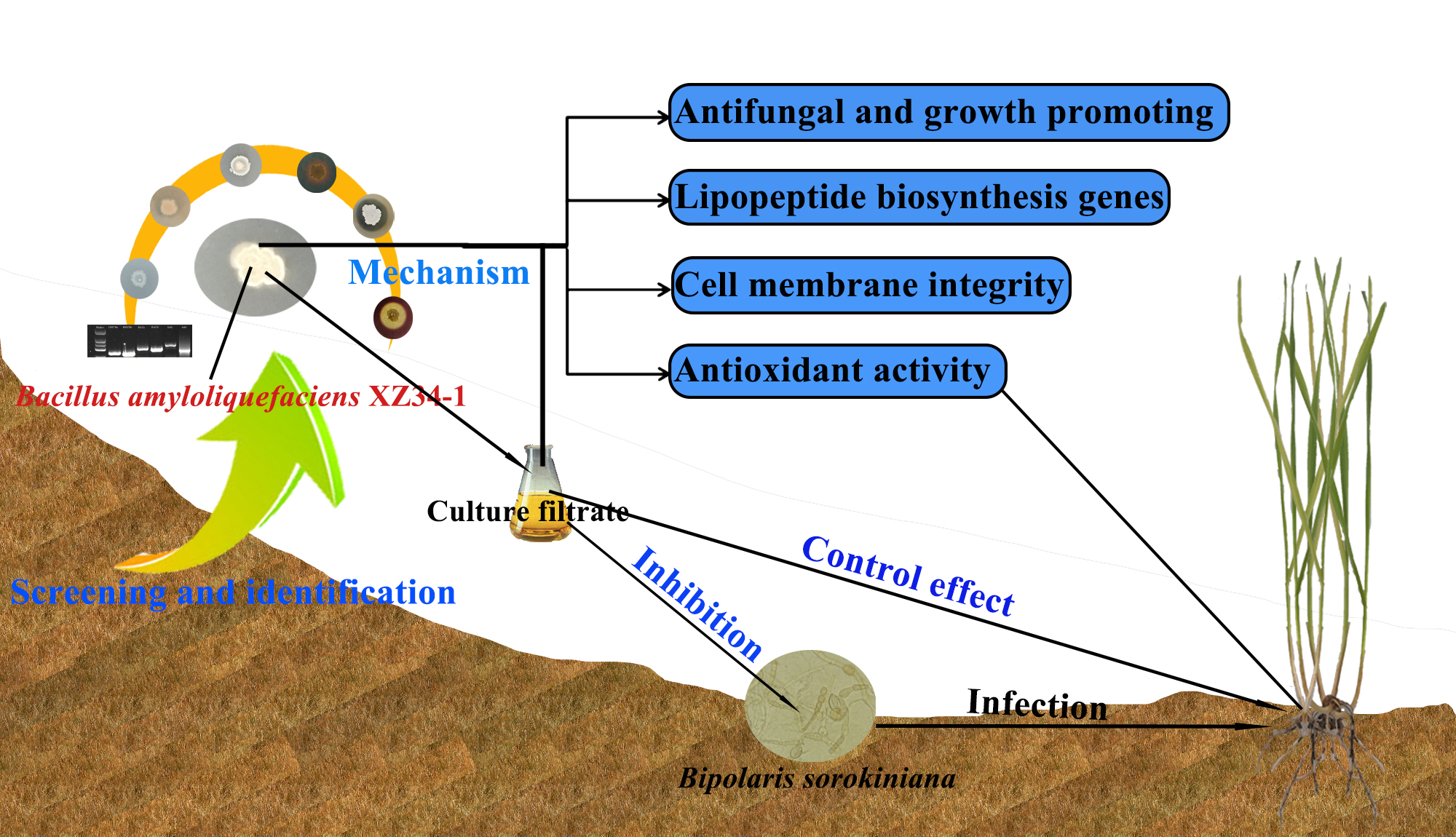
Antagonistic Strain Bacillus amyloliquefaciens XZ34-1 for Controlling Bipolaris sorokiniana and Promoting Growth in Wheat

 ,
, 
Abstract
:
1. Introduction
2. Results
2.1. Screening of Antagonistic Bacteria
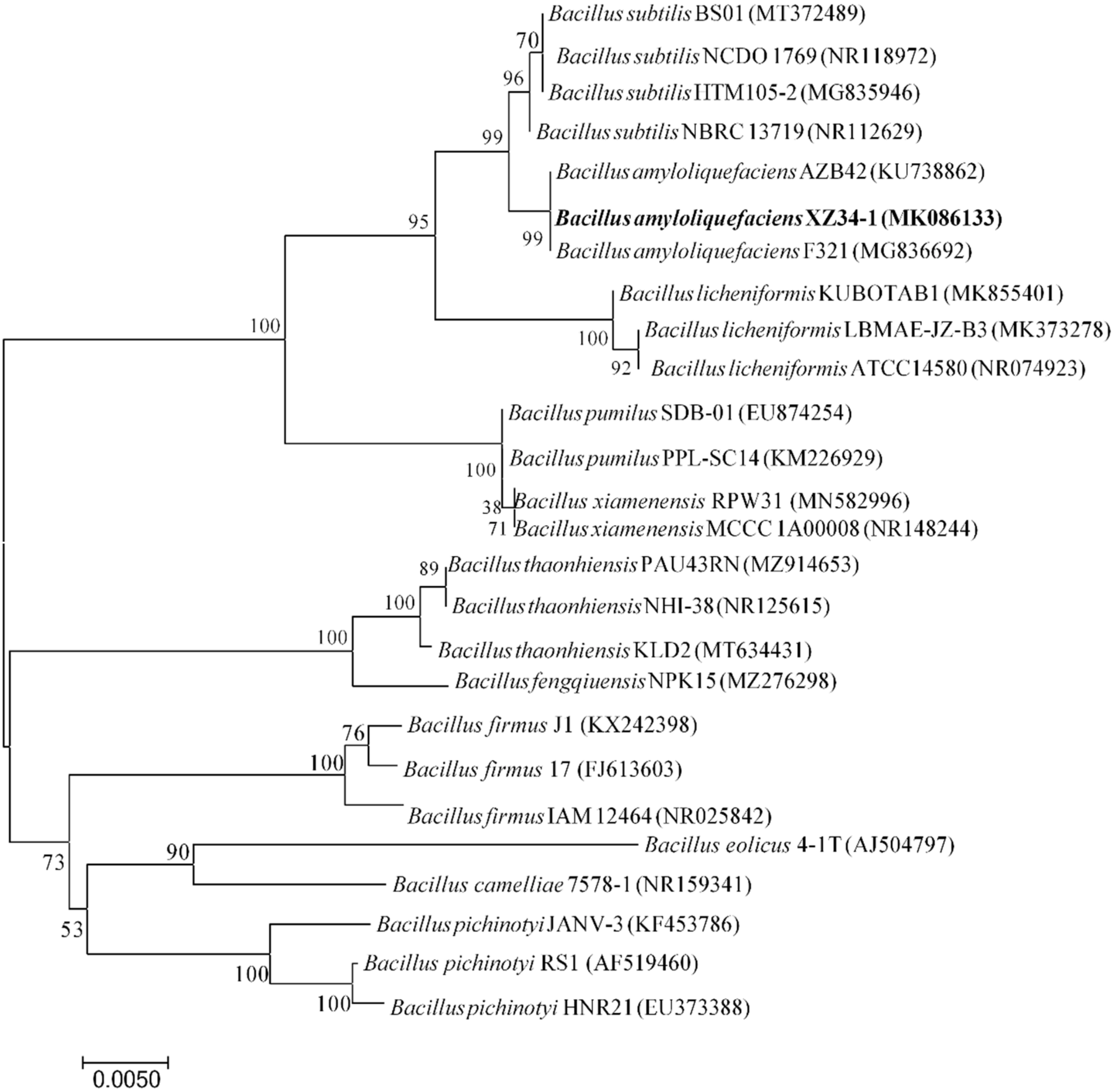
2.2. Identification of Antagonistic Strain XZ34-1
2.3. Inhibitory Effect of the Antagonistic Strain XZ34-1 CF on B. sorokiniana
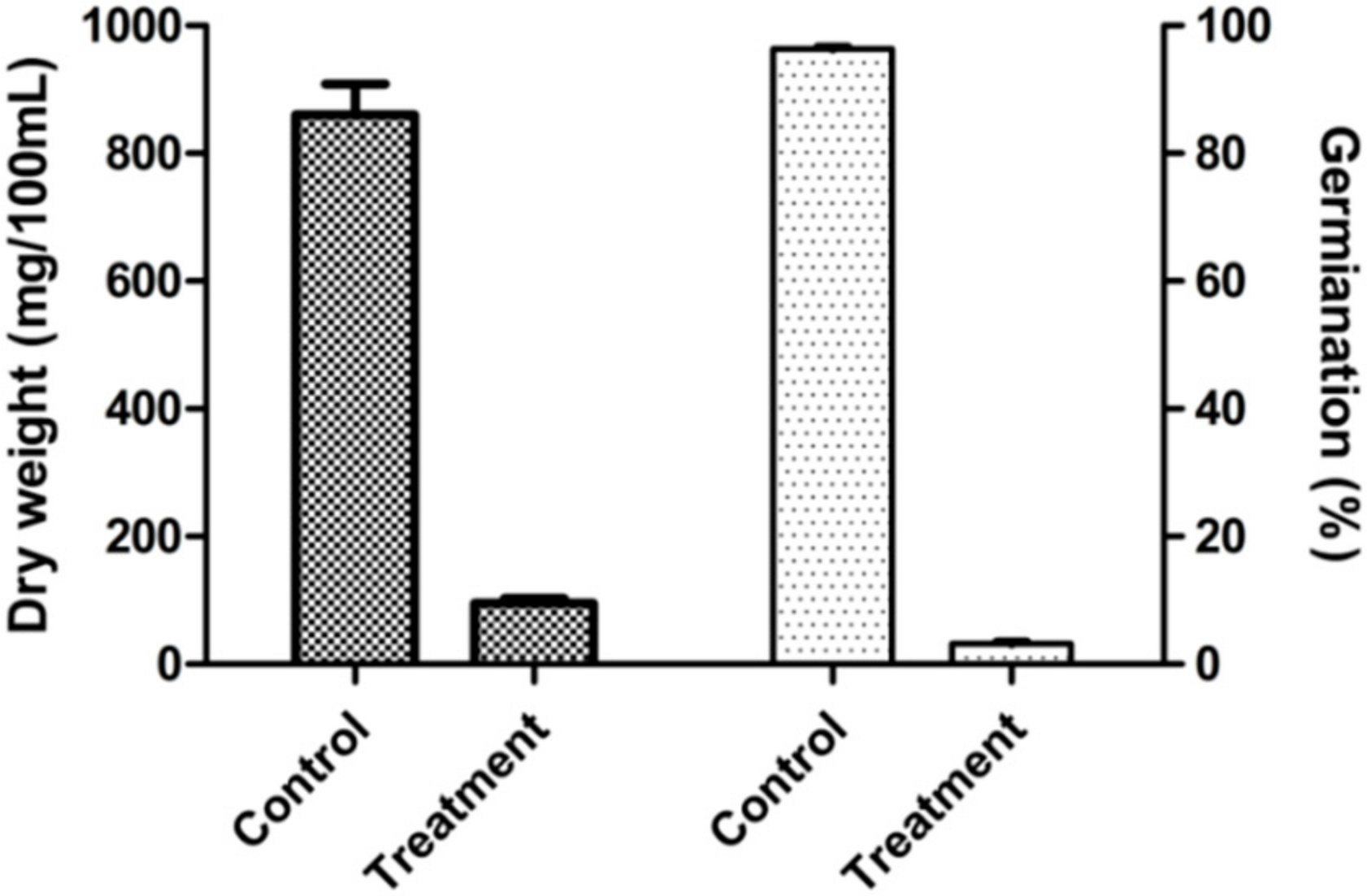
2.4. Biocontrol Efficacy of XZ34-1 CF on Common Root Rot Caused by B. sorokiniana
2.5. Potential of Strain XZ34-1 in Fungal Antagonism and Growth Promotion
2.6. Detection of Lipopeptide Biosynthesis Genes of Strain 34-1
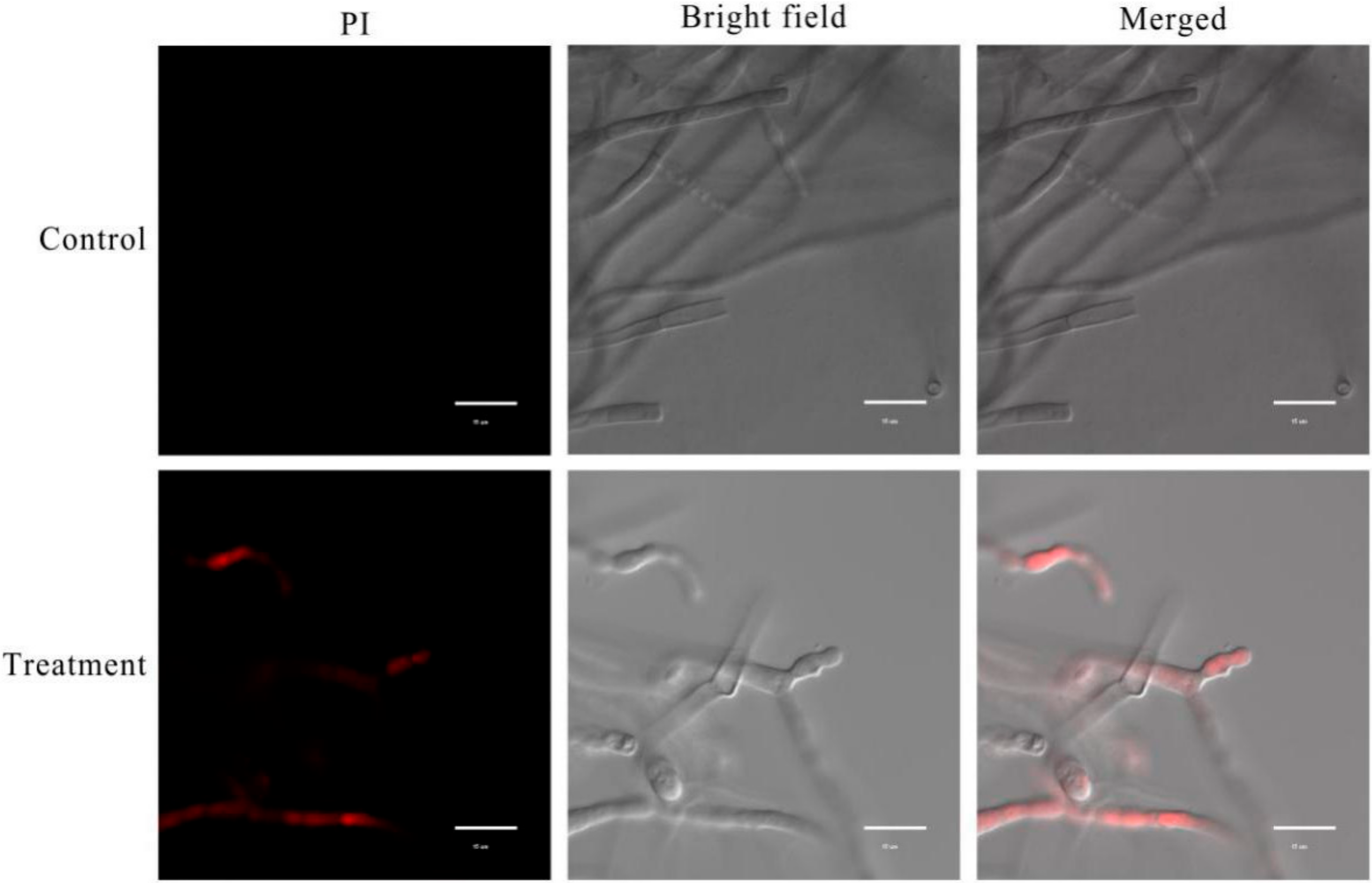
2.7. Effect of XZ34-1 CF on Cell Membrane Integrity in B. sorokiniana
2.8. Effects of XZ34-1 CF on Antioxidant Activity of Wheat Seedlings
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation and Screening of Antagonistic Bacteria
4.3. Identification of Strain XZ34-1
4.3.1. Morphological, Physiological and Biochemical Tests
4.3.2. Molecular Identification of Antagonistic Strain XZ34-1
4.4. Inhibitory Effect of the Antagonistic Strain XZ34-1 CF on B. sorokiniana
4.5. Pot Control Tests
4.6. Detection of Antifungal and Plant Growth-Promoting Traits
4.7. PCR Detection of Lipopeptide Biosynthesis Genes
4.8. Effect of CF on Cell Membrane Integrity in B. sorokiniana
4.9. Detection of Antioxidant Activity of Wheat Root
4.9.1. SOD Activity
4.9.2. POD Activity
4.9.3. PAL Activity
4.9.4. MDA Concentration
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Su, J.; Zhao, J.; Zhao, S.; Li, M.; Pang, S.; Chen, S.; Chen, F.; Zhen, W.; Kang, Z.; Wang, X. Genetics of Resistance to Common Root Rot (Spot Blotch), Fusarium Crown Rot, and Sharp Eyespot in Wheat. Front. Genet. 2021, 12, 699342. [Google Scholar] [CrossRef]
- Al-Sadi, A.M. Bipolaris sorokiniana-Induced Black Point, Common Root Rot, and Spot Blotch Diseases of Wheat: A Review. Front. Cell. Infect. Microbiol. 2021, 11, 9. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Deadman, M.L. Influence of Seed-borne Cochliobolus sativus (Anamorph Bipolaris sorokiniana) on Crown Rot and Root Rot of Barley and Wheat. J. Phytopathol. 2010, 158, 683–690. [Google Scholar] [CrossRef]
- Qostal, S.; Kribel, S.; Chliyeh, M.; Serghat, S.; Karimaselmaoui, A.O.; Touhami, A.O.; Zaarati, H.; Benkirane, R.; Allal Douira, A. Study of the fungal complex responsible for root rot of wheat and barley in the north-west of Morocco. Plant Arch. 2019, 19, 2143–2157. [Google Scholar]
- Fernandez, M.R.; Ulrich, D.; Brandt, S.A.; Zentner, R.P.; Wang, H.; Thomas, A.G.; Olfert, O. Crop Management Effects on Root and Crown Rot of Wheat in West-Central Saskatchewan, Canada. Agron. J. 2011, 103, 756–765. [Google Scholar] [CrossRef]
- Poole, G.J.; Harries, M.; Huberli, D.; Miyan, S.; MacLeod, W.J.; Lawes, R.; McKay, A. Predicting Cereal Root Disease in Western Australia Using Soil DNA and Environmental Parameters. Phytopathology 2015, 105, 1069–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Yang, G.Q.; Wang, J.M.; Song, Y.L.; Liu, L.L.; Zhao, K.; Li, Y.H.; Han, Z.H. Spatial Distribution of Root and Crown Rot Fungi Associated with Winter Wheat in the North China Plain and Its Relationship with Climate Variables. Front. Microbiol. 2018, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.X.; Zhang, W.L.; Cai, X.C.; Zhu, T.; Xue, Y.R.; Liu, C.H. Bacillus velezensis CC09: A Potential ’Vaccine’ for Controlling Wheat Diseases. Mol. Plant Microbe Interact. 2018, 31, 623–632. [Google Scholar] [CrossRef]
- Campanella, V.; Mandala, C.; Angileri, V.; Miceli, C. Management of common root rot and Fusarium foot rot of wheat using Brassica carinata break crop green manure. Crop Prot. 2020, 130, 7. [Google Scholar] [CrossRef]
- Saremi, H.; Saremi, H. Isolation of the most common Fusarium species and the effect of soil solarisation on main pathogenic species in different climatic zones of Iran. Eur. J. Plant Pathol. 2013, 137, 585–596. [Google Scholar] [CrossRef]
- Govaerts, B.; Mezzalama, M.; Sayre, K.; Crossa, J.; Lichter, K.; Troch, V.; Vanherck, K.; De Corte, P.; Deckers, J. Long-term consequences of tillage, residue management, and crop rotation on selected soil micro-flora groups in the subtropical highlands. Appl. Soil Ecol. 2008, 38, 197–210. [Google Scholar] [CrossRef]
- Shanmugam, V.; Kanoujia, N. Biological management of vascular wilt of tomato caused by Fusarium oxysporum f. sp lycospersici by plant growth-promoting rhizobacterial mixture. Biol. Control 2011, 57, 85–93. [Google Scholar] [CrossRef]
- Pereg, L.; McMillan, M. Scoping the potential uses of beneficial microorganisms for increasing productivity in cotton cropping systems. Soil Biol. Biochem. 2015, 80, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Ullah, H.; Yasmin, H.; Mumtaz, S.; Jabeen, Z.; Naz, R.; Nosheen, A.; Hassan, M.N. Multitrait Pseudomonas spp. Isolated from Monocropped Wheat (Triticum aestivum) Suppress Fusarium Root and Crown Rot. Phythopathology 2020, 110, 582–592. [Google Scholar] [CrossRef]
- Allali, K.; Goudjal, Y.; Zamoum, M.; Bouznada, K.; Sabaou, N.; Zitouni, A. Nocardiopsis dassonvillei strain MB22 from the Algerian Sahara promotes wheat seedlings growth and potentially controls the common root rot pathogen Bipolaris sorokiniana. J. Plant Pathol. 2019, 101, 1115–1125. [Google Scholar] [CrossRef]
- Yue, H.M.; Wang, M.; Gong, W.F.; Zhang, L.Q. The screening and identification of the biological control fungi Chaetomium spp. against wheat common root rot. FEMS Microbiol. Lett. 2018, 365, 6. [Google Scholar] [CrossRef] [PubMed]
- Ozlem, K.; Gary, Y.Y. Comparison of strains of Lysobacter enzymogenes and PGPR for induction of resistance against Bipolaris sorokiniana in tall fescue. Biol. Control 2004, 30, 446–455. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Cheng, X.K.; Ji, X.X.; Ge, Y.Z.; Li, J.J.; Qi, W.Z.; Qiao, K. Characterization of Antagonistic Bacillus methylotrophicus Isolated from Rhizosphere and Its Biocontrol Effects on Maize Stalk Rot. Phythopathology 2019, 109, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokry, M.E.; Ibrahim, S.D.; Wassief, A.Y. Biological control of pathogens associated with kernel black point disease of wheat. Crop Prot. 2017, 91, 13–19. [Google Scholar] [CrossRef]
- Wang, B.; Yuan, J.; Zhang, J.; Shen, Z.; Zhang, M.; Li, R.; Ruan, Y.; Shen, Q. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fert. Soils 2013, 49, 435–446. [Google Scholar] [CrossRef]
- Gong, A.D.; Li, H.P.; Yuan, Q.S.; Song, X.S.; Yao, W.; He, W.J.; Zhang, J.B.; Liao, Y.C. Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhang, Y.; Fu, X.; Li, Y.; Wang, Q. Isolation and characterization of Bacillus amyloliquefaciens PG12 for the biological control of apple ring rot. Postharvest Biol. Technol. 2016, 115, 113–121. [Google Scholar] [CrossRef]
- Yu, Y.Y.; Xu, J.D.; Huang, T.X.; Zhong, J.; Yu, H.; Qiu, J.P.; Guo, J.H. Combination of beneficial bacteria improves blueberry production and soil quality. Food Sci. Nutr. 2020, 8, 5776–5784. [Google Scholar] [CrossRef]
- Abadi, V.; Sepehri, M.; Rahmani, H.A.; Dolatabad, H.K.; Shamshiripour, M.; Khatabi, B. Diversity and abundance of culturable nitrogen-fixing bacteria in the phyllosphere of maize. J. Appl. Microbiol. 2021, 131, 898–912. [Google Scholar] [CrossRef] [PubMed]
- Niazi, A.; Manzoor, S.; Asari, S.; Bejai, S.; Meijer, J.; Bongcam, R.E. Genome analysis of Bacillus amyloliquefaciens Subspp. plantarum UCMB5113: A rhizobacterium that improves plant growth and stress management. PLoS ONE 2014, 9, e104651. [Google Scholar] [CrossRef]
- Farzand, A.; Moosa, A.; Zubair, M.; Khan, A.R.; Hanif, A.; Tahir, H.A.S.; Gao, X.W. Marker assisted detection and LC-MS analysis of antimicrobial compounds in different Bacillus strains and their antifungal effect on Sclerotinia sclerotiorum. Biol. Control 2019, 133, 91–102. [Google Scholar] [CrossRef]
- Zhang, L.; Khabbaz, S.E.; Wang, A.; Li, H.; Abbasi, P.A. Detection and characterization of broad-spectrum antipathogen activity of novel rhizobacterial isolates and suppression of Fusarium crown and root rot disease of tomato. J. Appl. Microbiol. 2015, 118, 685–703. [Google Scholar] [CrossRef] [PubMed]
- Mora, I.; Cabrefiga, J.; Montesinos, E. Antimicrobial peptide genes in Bacillus strains from plant environments. Int. Microbiol. 2011, 14, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compaore, C.S.; Nielsen, D.S.; Ouoba, L.I.I.; Berner, T.S.; Nielsen, K.F.; Sawadogo-Lingani, H.; Diawara, B.; Ouedraogo, G.A.; Jakobsen, M.; Thorsen, L. Co-production of surfactin and a novel bacteriocin by Bacillus subtilis subsp subtilis H4 isolated from Bikalga, an African alkaline Hibiscus sabdariffa seed fermented condiment. Int. J. Food Microbiol. 2013, 162, 297–307. [Google Scholar] [CrossRef]
- Athukorala, S.N.P.; Fernando, W.G.D.; Rashid, K.Y. Identification of antifungal antibiotics of Bacillus species isolated from different microhabitats using polymerase chain reaction and MALDI-TOF mass spectrometry. Can. J. Microbiol. 2009, 55, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Khedher, S.B.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control 2021, 152, 10. [Google Scholar] [CrossRef]
- Chung, S.; Kong, H.; Buyer, J.S.; Lakshman, D.K.; Lydon, J.; Kim, S.D.; Roberts, D.P. Isolation and partial characterization of Bacillus subtilis ME488 for suppression of soilborne pathogens of cucumber and pepper. Appl. Microbiol. Biotechnol. 2008, 80, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Villa-Rodriguez, E.; Parra-Cota, F.; Castro-Longoria, E.; Lopez-Cervantes, J.; Santos-Villalobos, S.D. Bacillus subtilis TE3: A promising biological control agent against Bipolaris sorokiniana, the causal agent of spot blotch in wheat (Triticum turgidum L. subspp. durum). Biol. Control 2019, 132, 135–143. [Google Scholar] [CrossRef]
- Zakry, F.A.A.; Shamsuddin, Z.H.; Rahim, K.A.; Zakaria, Z.Z.; Rahim, A.A. Inoculation of Bacillus sphaericus UPMB-10 to Young Oil Palm and Measurement of Its Uptake of Fixed Nitrogen Using the N-15 Isotope Dilution Technique. Microbes Environ. 2012, 27, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Tahir, M.; Khalid, U.; Ijaz, M.; Shah, G.M.; Naeem, M.A.; Shahid, M.; Mahmood, K.; Ahmad, N.; Kareem, F. Combined application of bio-organic phosphate and phosphorus solubilizing bacteria (Bacillus strain MWT 14) improve the performance of bread wheat with low fertilizer input under an arid climate. Braz. J. Microbiol. 2018, 49, 15–24. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.R.; Klein, M.N.; Ferraz, L.P.; da Silva, A.C.; Kupper, K.C. Saccharomyces cerevisiae: A novel and efficient biological control agent for Colletotrichum acutatum during pre-harvest. Microbiol. Res. 2015, 175, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Berini, F.; Casartelli, M.; Montali, A.; Reguzzoni, M.; Tettamanti, G.; Marinelli, F. Metagenome-Sourced Microbial Chitinases as Potential Insecticide Proteins. Front. Microbiol. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Hjort, K.; Presti, I.; Elvang, A.; Marinelli, F.; Sjoling, S. Bacterial chitinase with phytopathogen control capacity from suppressive soil revealed by functional metagenomics. Appl. Microbiol. Biotechnol. 2014, 98, 2819–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, D.; Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.W.; Arrebola, E.; Cazorla, F.M.; Kuipers, O.P.; Paquot, M.; Pérez-García, A. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol. Plant Microbe Interact. 2007, 20, 430–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongena, M.; Jourdan, E.; Adam, A.; Michel, P.; Alain, B.; Bernard, J.; Arpigny, J.L.; Philippe, T. Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Environ. Microbiol. 2007, 9, 1084–1090. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, J.; Zhang, C.; Wang, L.; Gao, W.; Jiang, J. Bacillus megaterium WL-3 Lipopeptides Collaborate against Phytophthora infestans to Control Potato Late Blight and Promote Potato Plant Growth. Front. Microbiol. 2020, 11, 1602. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, C.; Liang, J.; Wu, L.; Gao, W.; Jiang, J. Iturin A Extracted from Bacillus subtilis WL-2 Affects Phytophthora infestans via Cell Structure Disruption, Oxidative Stress, and Energy Supply Dysfunction. Front. Microbiol. 2020, 11, 536083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, C. Fengycins, cyclic lipopeptides from marine Bacillus subtilis strains, kill the plant-pathogenic fungus Magnaporthe grisea by inducing reactive oxygen species production and chromatin condensation. Appl. Environ. Microbiol. 2018, 84, e00445–e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Lee, Y.; Cheon, W.; Park, J.; Kwon, H.T.; Balaraju, K.; Kim, J.; Yoon, Y.J.; Jeon, Y. Characterization of Bacillus velezensis AK-0 as a biocontrol agent against apple bitter rot caused by Colletotrichum gloeosporioides. Sci. Rep. 2021, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.M.; Yoon, M.Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Shin, T.S.; Park, H.W.; Yu, N.H.; Kim, Y.H.; Kim, J.C. Diffusible and Volatile Antifungal Compounds Produced by an Antagonistic Bacillus velezensis G341 against Various Phytopathogenic Fungi. Plant Pathol. J. 2017, 33, 488–498. [Google Scholar] [CrossRef] [Green Version]
- Jamali, H.; Sharma, A.; Roohi; Srivastava, A.K. Biocontrol potential of Bacillus subtilis RH5 against sheath blight of rice caused by Rhizoctonia solani. J. Basic Microbiol. 2020, 60, 268–280. [Google Scholar] [CrossRef]
- Van Wees, S.C.M.; Van der Ent, S.; Pieterse, C.M.J. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Kumar, V.; Yuan, H.; Kanwar, M.K.; Bhardwaj, R.; Thukral, A.K.; Zheng, B. Jasmonic Acid Seed Treatment Stimulates Insecticide Detoxification in Brassica juncea L. Front. Plant Sci. 2018, 9, 1609. [Google Scholar] [CrossRef]
- Fatma, F.; Verma, S.; Kamal, A.; Srivastava, A. Phytotoxicity of pesticides mancozeb and chlorpyrifos: Correlation with the antioxidative defence system in Allium cepa. Physiol. Mol. Biol. Plants 2018, 24, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Zhou, X.N.; Cai, Z.B.; Guo, L.; Chen, X.L.; Chen, X.; Liu, J.Y.; Feng, M.F.; Qiu, Y.W.; Zhang, Y.; et al. A Biocontrol Strain of Pseudomonas aeruginosa CQ-40 Promote Growth and Control Botrytis cinerea in Tomato. Pathogens 2021, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Dubey, R.C.; Maheshwari, D.K. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol. Res. 2012, 167, 493–499. [Google Scholar] [CrossRef]
- Guo, D.S.; Yuan, C.H.; Luo, Y.Y.; Chen, Y.H.; Lu, M.H.; Chen, G.C.; Ren, G.W.; Cui, C.B.; Zhang, J.T.; An, D.R. Biocontrol of tobacco black shank disease (Phytophthora nicotianae) by Bacillus velezensis Ba168. Pestic. Biochem. Physiol. 2020, 165, 10. [Google Scholar] [CrossRef]
- Guerrero, R. Bergey’s manuals and the classification of prokaryotes. Int. Microbiol. 2001, 4, 103–109. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Wu, Y.D.; Chong, X.Y.; Xin, Q.H.; Wang, D.X.; Bian, K. Seed-borne endophytic Bacillus velezensis LHSB1 mediate the biocontrol of peanut stem rot caused by Sclerotium rolfsii. J. Appl. Microbiol. 2020, 128, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, Y.; Ha, A.; Kim, J.I.; Park, A.R.; Yu, N.H.; Son, H.; Choi, G.J.; Park, H.W.; Lee, C.W.; et al. Chemosensitization of Fusarium graminearum to Chemical Fungicides Using Cyclic Lipopeptides Produced by Bacillus amyloliquefaciens Strain JCK-12. Front. Plant Sci. 2017, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Vaishnav, A.; Jain, S.; Kasotia, A.; Kumari, S.; Gaur, R.K.; Choudhary, D.K. Effect of nitric oxide signaling in bacterial-treated soybean plant under salt stress. Arch. Microbiol. 2013, 195, 571–577. [Google Scholar] [CrossRef]
- He, S.Z.; Han, Y.F.; Wang, Y.P.; Zhai, H.; Liu, Q.C. In vitro selection and identification of sweetpotato (Ipomoea batatas (L.) Lam.) plants tolerant to NaCl. Plant Cell Tissue Organ Cult. 2009, 96, 69–74. [Google Scholar] [CrossRef]
- Choudhary, D.K. Plant growth-promotion (PGP) activities and molecular characterization of rhizobacterial strains isolated from soybean (Glycine max L. Merril) plants against charcoal rot pathogen, Macrophomina phaseolina. Biotechnol. Lett. 2011, 33, 2287–2295. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Choudhary, D.K. Induced defense-related proteins in soybean (Glycine max L. Merrill) plants by Carnobacterium sp SJ-5 upon challenge inoculation of Fusarium oxysporum. Planta 2014, 239, 1027–1040. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Inhibition Rate (%) | ||||
|---|---|---|---|---|---|
| Bipolaris sorokiniana | Fusarium graminearum | Rhizoctonia zeae | Aspergillus niger | Aspergillus flavus | |
| XZ34-1 | 53.52 ± 0.86 a | 43.06 ± 1.73 a | 46.70 ± 1.37 a | 35.77 ± 1.22 a | 34.35 ± 1.49 a |
| XZ33-1 | 32.66 ± 1.77 b | 30.41 ± 0.75 b | 33.93 ± 0.59 b | 19.98 ± 2.84 b | 26.94 ± 1.42 b |
| XZ22-1 | 22.02 ± 2.25 c | 12.41 ± 1.77 d | 26.70 ± 2.58 c | -- | -- |
| XZ19-2 | 15.86 ± 3.04 d | 19.94 ± 1.17 c | 26.25 ± 1.69 c | 9.22 ± 1.62 c | 8.73 ± 1.73 c |
| XZ11-2 | 15.17 ± 2.29 d | 13.59 ± 2.42 d | 12.64 ± 1.87 d | -- | -- |
| Morphological and Biochemical Characteristics | XZ34-1 | |
|---|---|---|
| Cell morphology | Gram’s reaction | Gram positive |
| Endospores | + | |
| Colony morphology | Morphology | Rounded |
| Pigment | Creamy white | |
| Surface | Rough | |
| Margin | Smooth | |
| Opacity | Opaque | |
| Biochemical tests | Nitrate reduction | − |
| Citrate utilization | − | |
| V-P test | + | |
| Methyl Red | − | |
| Glucose decomposition | Acids, not gas | |
| Treatments | Disease Incidence Rate (%) | Disease Index | Control Efficacy (%) |
|---|---|---|---|
| Sterile water control | 98.05 ± 2.76 a | 65.73 ± 11.29 a | − |
| Carbendazim treatment | 41.24 ± 10.87 c | 9.16 ± 2.20 b | 85.67 ± 4.67 |
| CF treatment | 54.13 ± 9.16 b | 14.17 ± 3.01 b | 78.24 ± 4.66 |
| Product | Genes | Primers | Primer Sequence (5′-3′) | Expected/Detected Size (bp) | References |
|---|---|---|---|---|---|
| Bacyllomicin | BMYBa | BMYBa-R | CGAAACGACGGTATGAAT | 371/yes | Farzand, A. et al. [27] |
| BMYBa-R | TCTGCCGTTCCTTATCTC | ||||
| BMYBb | BMYBb-F | GAATCCCGTTGTTCTCCAAA | 370/yes | Zhang, L. et al. [28] | |
| BMYBb-R | GCGGGTATTGAATGCTTGTT | ||||
| Bacilysin | BACa | BACa-F | ATCTTTATGGCGGCAGTC | 595/yes | Farzand, A. et al. [27] |
| BACa-R | ATACGGCTTACAGGCGAG | ||||
| BACb | BACb-F | CAGCTCATGGGAATGCTTTT | 498/yes | Mora, I. et al. [29] | |
| BACb-R | CTCGGTCCTGAAGGGACAAG | ||||
| Bacillaene | BAE | BAE-F | ATGTCAGCTCAGTTTCCGCA | 688/yes | Compaore, C. S. et al. [30] |
| BAE-R | GATCGCCGTCTTCAATTGCC | ||||
| Iturin | ITU | ITU-F | GGCTGCTGCAGATGCTTTAT | 423/yes | Mora, I. et al. [29] |
| ITU-R | TCGCAGATAATCGCAGTGAG | ||||
| Fengycin | fenD | FEND-F | TTTGGCAGCAGGAGAAGTTT | 964/no | Athukorala, S. N. P. et al. [31] |
| FEND-R | GCTGTCCGTTCTGCTTTTTC | ||||
| Surfactin | srfAA | srfAA-F | TCGGGACAGGAAGACATCAT | 201/no | Khedher, S.B. et al. [32] |
| srfAA-R | CCACTCAAACGGATAATCCTG | ||||
| sfP | SFP-F | ATGAAGATTTACGGAATTTA | 675/no | Chung, S. et al. [33] | |
| SFP-R | TTATAAAAGCTCTTCGTACG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, Y.; Shan, Y.; Liu, S.; Yang, Y.; Liu, Y.; Yin, Y.; Hou, Z.; Luan, P.; Li, R. Antagonistic Strain Bacillus amyloliquefaciens XZ34-1 for Controlling Bipolaris sorokiniana and Promoting Growth in Wheat. Pathogens 2021, 10, 1526. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111526
Yi Y, Shan Y, Liu S, Yang Y, Liu Y, Yin Y, Hou Z, Luan P, Li R. Antagonistic Strain Bacillus amyloliquefaciens XZ34-1 for Controlling Bipolaris sorokiniana and Promoting Growth in Wheat. Pathogens. 2021; 10(11):1526. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111526
Chicago/Turabian StyleYi, Yanjie, Youtian Shan, Shifei Liu, Yanhui Yang, Yang Liu, Yanan Yin, Zhipeng Hou, Pengyu Luan, and Ruifang Li. 2021. "Antagonistic Strain Bacillus amyloliquefaciens XZ34-1 for Controlling Bipolaris sorokiniana and Promoting Growth in Wheat" Pathogens 10, no. 11: 1526. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111526