
Fungal Species Causing Maize Leaf Blight in Different Agro-Ecologies in India

 , and
, and
Abstract
:1. Introduction
2. Results
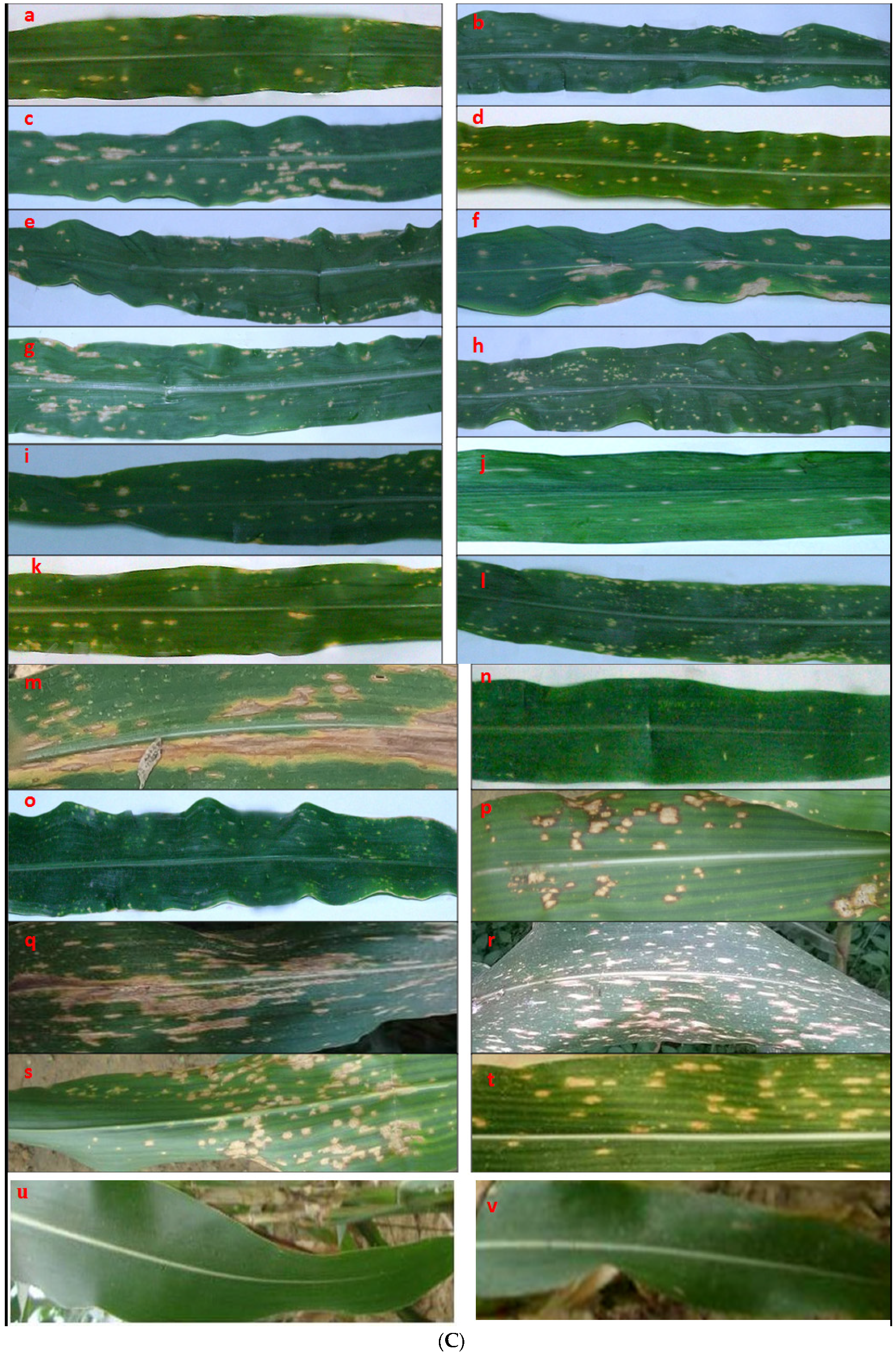
2.1. Survey, Collection, and Maintenance of Fungal Isolates
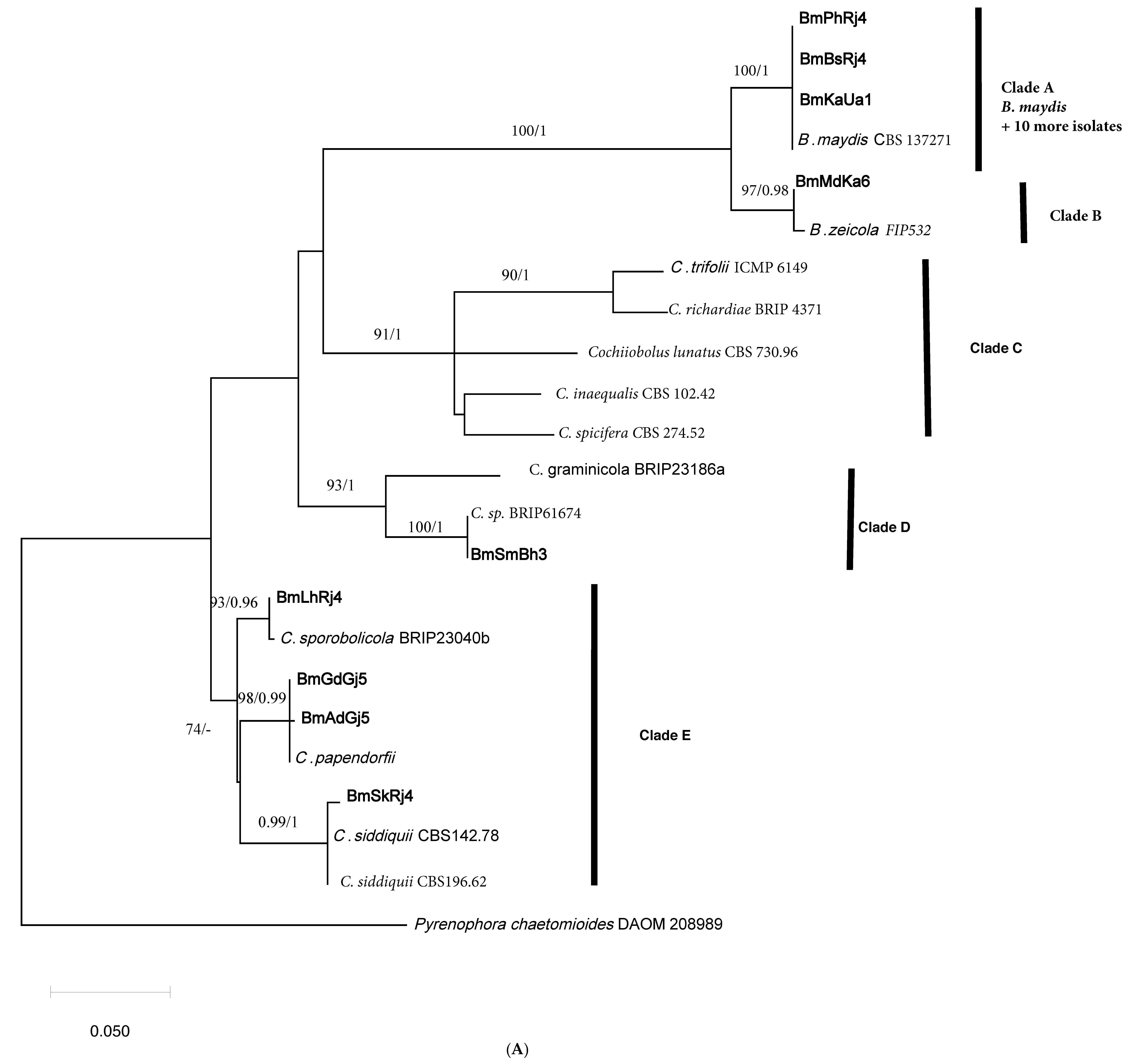
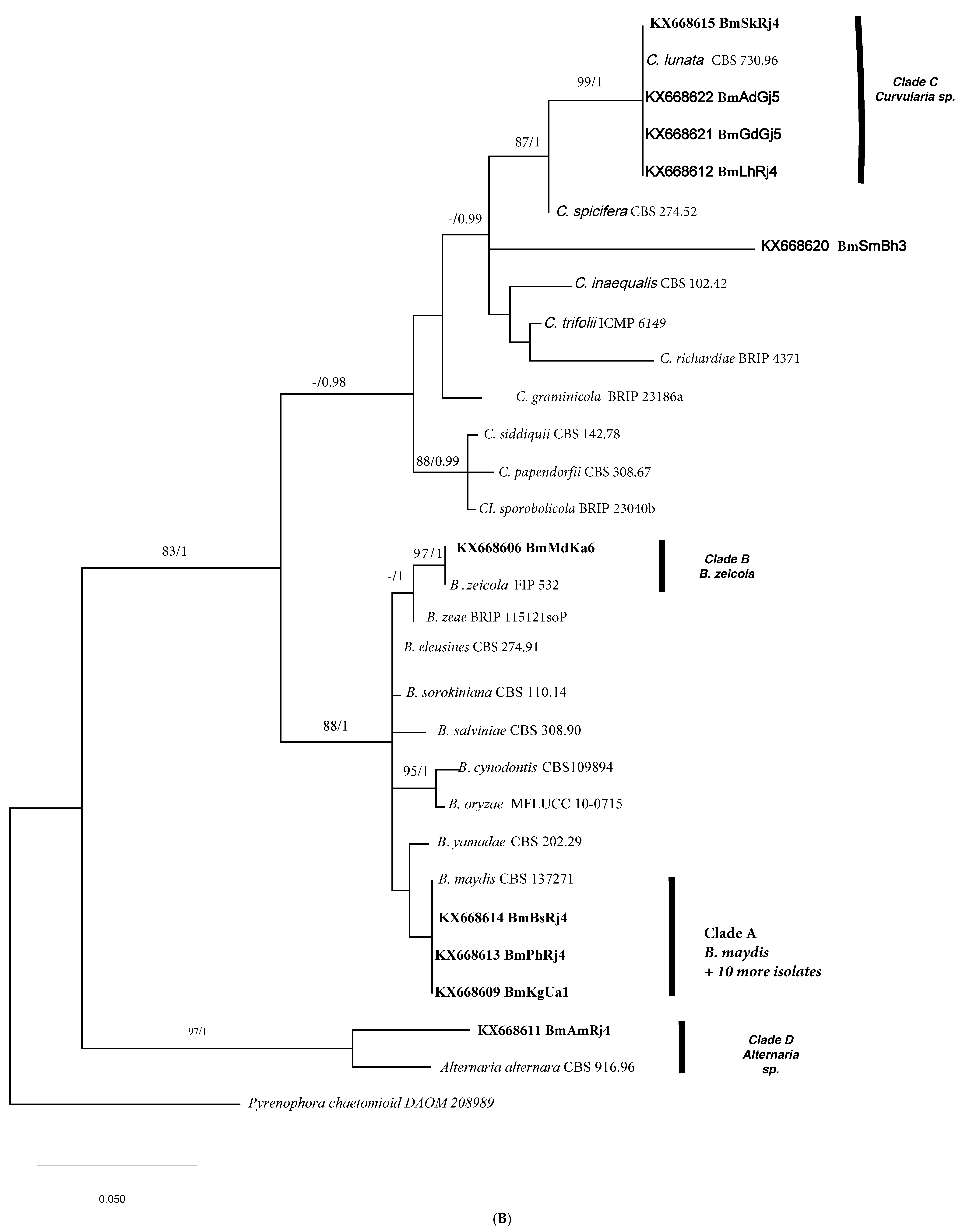
2.2. Phylogenetic Analyses
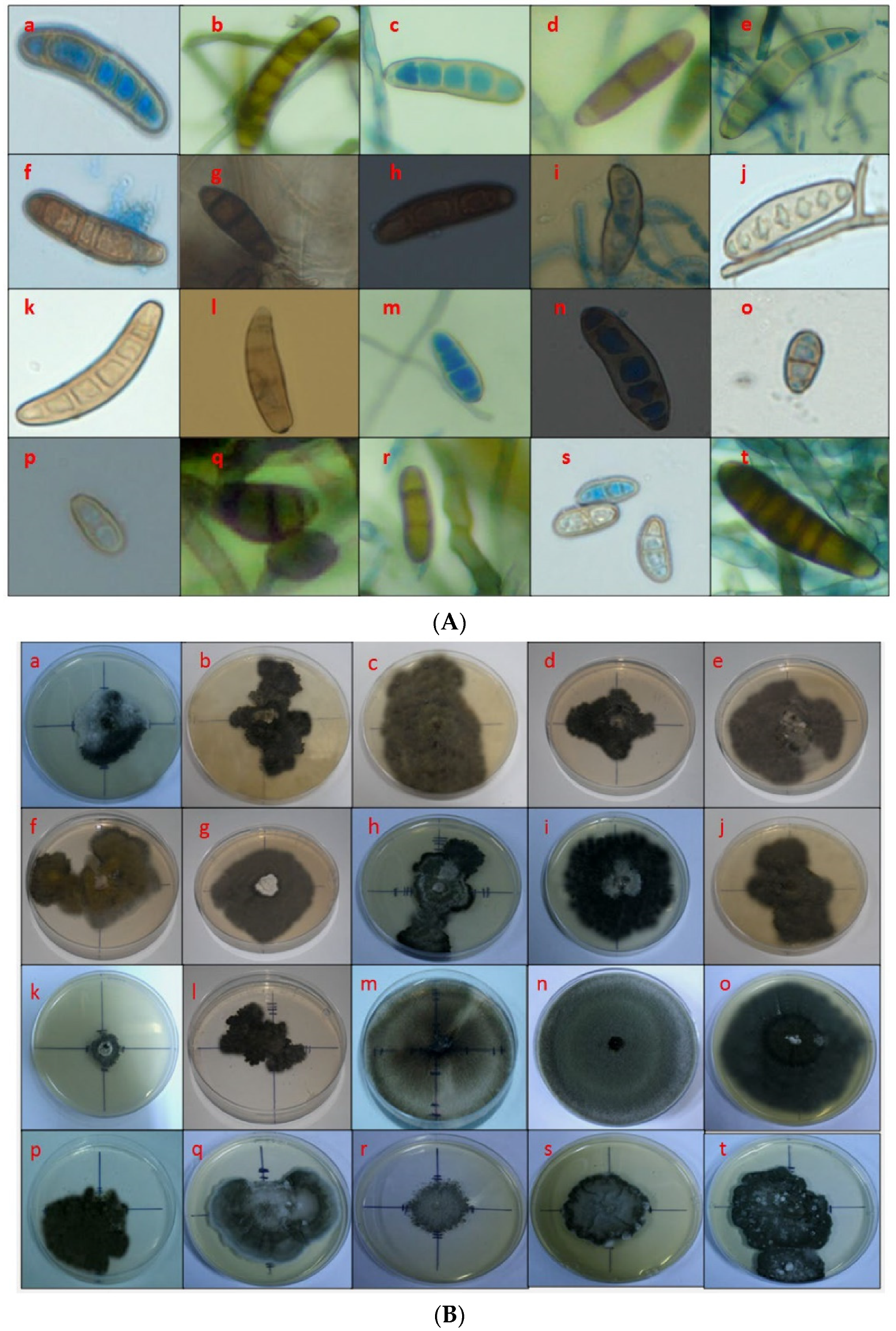
2.3. Morphological Characterization of Fungal sp. Causing Maize Leaf Blight
2.4. Pathogenic Variability of Fungal Isolates on Zea mays c.v. DHM 117
3. Discussion
4. Materials and Methods
4.1. Collection and Maintenance of Isolates
4.2. Fungal DNA Extraction, Amplification, Sequencing and Phylogenetic Analysis
4.3. Phylogenetic Analysis
4.4. Cultural and Morphological Variation
4.5. Pathogenic Variability
- Very mild infection, as 1 to 2 or more scattered lesions on lower leaves of the host.
- Moderate infection showing few lesions on lower leaves only of the host.
- Moderate infection, with abundant lesions on lower leaves, spreading up to middle leaves and extending to upper leaves of the host.
- Severe infection showing abundant lesions on lower and middle leaves, extending to upper leaves of the host.
- Intense severity with abundant lesions on almost all the leaves showing premature drying or necrosis of infected leaf tissue.
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yadav, O.P.; Hossain, F.; Karjagi, C.G.; Kumar, B.; Zaidi, P.H.; Jat, S.L.; Chawla, J.S.; Kaul, J.; Hooda, K.S.; Kumar, P.; et al. Genetic improvement of maize in India: Retrospect and prospects. Agric. Res. 2015, 4, 325–338. [Google Scholar] [CrossRef]
- ICAR-IIMR Annual Report. ICAR-Indian Institute of Maize Research, Punjab Agricultural University Campus, Ludhiana–141004; ICAR-IIMR: Telangana, Indian. Available online: https://iimr.icar.gov.in/publications-category/annual-reports/ (accessed on 9 November 2021).
- Kaur, H.; Hooda, K.S.; Khokhar, M.K. Maydis leaf blight of maize: Historical perspective, impact and present status. Maize J. 2014, 3, 1–8. [Google Scholar]
- Manamgoda, D.S.; Rossman, A.Y.; Castlebury, L.A.; Crous, P.W.; Madrid, H.; Chukeatirote, E.; Hyde, K.D. The genus Bipolaris. Stud. Mycol. 2014, 79, 221–288. [Google Scholar] [CrossRef] [Green Version]
- Rossman, A.Y.; Manamgoda, D.S.; Hyde, K.D. Proposal to conserve the name Bipolaris against Cochliobolus (Ascomycota: Pleosporales: Pleosporaceae). Taxon 2013, 62, 1331–1332. [Google Scholar] [CrossRef]
- Manamgoda, D.; Cai, L.; Bahkali, A.; Chukeatirote, E.; Hyde, K.D. Cochliobolus: An overview and current status of species. Fung. Diver. 2011, 51, 3–42. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, Systematic Mycology and Microbiology Laboratory; ARS, USDA: Beltsville, MD, USA, 2013. Available online: http://nt.arsgrin.gov/fungaldatabases (accessed on 9 November 2021).
- Zhang, X.F.; Cui, L.N.; Li, X.; Zou, C.J.; Yang, X.R. Biological characters and the resistance study of corn leaf spot caused by Bipolaris zeicola in southwest China. J. Maize Sci. 2013, 21, 128–133. [Google Scholar]
- Berbee, M.L.; Carmean, D.A.; Winka, K. Ribosomal DNA and resolution of branching order among the ascomycota: How many nucleotides are enough? Mol. Phylogenet. Evol. 2000, 17, 337–344. [Google Scholar] [CrossRef]
- Hernandez-Restrepo, M.; Madrid, H.; Tan, Y.P.; Da Cunha, K.C.; Gene, J.; Guarro, J.; Crous, P.W. Multi-locus phylogeny and taxonomy of Exserohilum. Persoonia 2018, 41, 71–108. [Google Scholar] [CrossRef] [PubMed]
- Boedijn, K.B. Ueber einige phragmosporen Dematiazeen. Ann. Jard. Bot. Buitenzorg 1933, 13, 123. [Google Scholar]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Manamgoda, D.S.; Cai, L.; McKenzie, E.H.C.; Crous, P.W.; Madrid, H.; Chukeatirote, E.; Shivas, R.G.; Tan, Y.P.; Hyde, K.D. A phylogenetic and taxonomic re-evaluation of the Bipolaris–Cochliobolus–Curvularia complex. Fung. Diver. 2012, 56, 131–144. [Google Scholar] [CrossRef]
- Zhang, L.X.; Dong, M.; Yang, L.M.; Wang, J.H.; Tan, G.J. Identification of physiological races of Bipolaris maydis and their sensitivities to diniconazole in Anhui Province. Acta Phytopathol. Sin. 2011, 41, 441–444. [Google Scholar]
- Ullstrup, A.J. The impacts of the southern corn leaf blight epidemics of 1970–1971. Annu. Rev. Phytopathol. 1972, 10, 37–50. [Google Scholar] [CrossRef]
- Misra, A.P. Variability, physiologic specialization and genetics of pathogenicity in graminicolous Helminthosporia affecting cereal crops. Indian Phytopath. 1979, 32, 1–22. [Google Scholar]
- Carson, M.L. Aggressiveness and perennation of isolates of Cochliobolus heterostrophus from North Carolina. Plant Dis. 1998, 82, 1043–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lev, S.; Sharon, A.; Hadar, R.; Ma, H.; Horwitz, B.A. A mitogen-activated protein kinase of the corn leaf pathogen Cochliobolus heterostrophus is involved in conidiation, appressorium formation, and pathogenicity: Diverse roles for mitogen activated protein kinase homologs in foliar pathogens. Proc. Nat. Acad. Sci. USA 1999, 96, 13542–13547. [Google Scholar] [CrossRef] [Green Version]
- Fatima, R.; Mathur, S.B.; Paul, N. Importance of Drechslera maydis on seed crops other than maize. Seed Sci. Technol. 1974, 2, 371–383. [Google Scholar]
- Sun, X.; Qi, X.; Wang, W.; Liu, X.; Zhao, H.; Wu, C.; Chang, X.; Zhang, M.; Chen, H.; Gong, G. Etiology and symptoms of maize leaf spot caused by Bipolaris spp. in Sichuan, China. Pathogens 2020, 9, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogoi, R.; Singh, S.; Singh, P.K.; Kulanthaivel, S.; Rai, S.N. Genetic variability in the isolates of Bipolaris maydis causing maydis leaf blight of maize. Afr. J. Agric. Res. 2014, 9, 1906–1913. [Google Scholar] [CrossRef]
- Mitra, M. India: New diseases reported during the year 1931. Int. Bull. Pl. Prot. 1931, 7, 103–104. [Google Scholar]
- Sharma, R.C.; Lilaramani, J.; Payak, M.M. Outbreak of a new pathotype of Helminthosporium maydis on maize in India. Indian Phytopath. 1978, 31, 112–113. [Google Scholar]
- Wei, J.K.; Liu, K.M.; Chen, J.P.; Luo, P.C.; Stadelmann, O.Y.L. Pathological and physiological identification of race C of Bipolaris maydis in China. Phytopathology 1988, 78, 550–554. [Google Scholar] [CrossRef]
- Mandokhot, A.M.; Chaudhary, B.K.C. A new leaf spot of maize incited by Curvularia clavata. Neth J. Plant Pathol. 1972, 78, 65–68. [Google Scholar] [CrossRef]
- Gopi, R. Pathogenic and Molecular Variability in Bipolaris maydis Incitant of Maydis Leaf Blight in Maize. Ph.D. Thesis, P.G. School, Indian Agricultural Research Institute, New Delhi, India, 2008. [Google Scholar]
- Yadav, R.L.; Bunker, R.N.; Sharma, S.S. Morphological and pathogenic variability among isolates of Bipolaris maydis causing maydis leaf blight of maize. J. Mycol. Plant Pathol. 2013, 43, 401–408. [Google Scholar]
- Pal, I.; Singh, V.; Gogoi, R.; Hooda, K.S.; Bedi, N. Characterization of Bipolaris maydis isolates of different maize cropping zones of India. Indian Phytopath. 2015, 68, 63–66. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Dong, Y.; Jayawardena, R.; Jeewon, R.; Phukhamsakda, C.; Bundhun, D.; Hyde, K.D.; Sheng, J. A polyphasic approach to delineate species in Bipolaris. Fungal Divers. 2020, 102, 225–256. [Google Scholar] [CrossRef]
- Lu, C.H.; Wu, Y.X.; Ho, H.H.; Mao, Z.C.; He, Y.Q. Identification of races and mating types of Cochliobolus carbonum from corn in the Yunnan Province in China. J. Phytopathol. 2014, 162, 313–321. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Q.; Sun, G.Y.; Mao, Q.; Gleason, M.L. First report of race 3 of Bipolaris zeicola on corn in China. Plant Dis. 2007, 91, 1360. [Google Scholar] [CrossRef]
- Kumar, M.; Agarwal, V.K. Location of seed borne fungi associated with discoloured maize seeds. Indian Phytopathol. 1998, 51, 247–250. [Google Scholar]
- Viswanathan, R.; Narayanasamy, P. Nature of rice phylloplane microorganisms and their sensitivity to fungicides. Acta Botan. Indic. 1991, 19, 236–238. [Google Scholar]
- Xiao, J.Z.; Tsuge, T.; Doke, N.; Nakatsuka, S.; Tsuda, M.; Nishimura, S. Rice-specific toxins produced by Bipolaris zeicola, race 3; Evidence for role as pathogenicity factors for rice and maize plants. Physiol. Mol. Plant Pathol. 1991, 38, 67–82. [Google Scholar] [CrossRef]
- Aslam, H.M.U.; Naveed, K.; Hussain, S.I.; Shakeel, Q.; Ashraf, W.; Anwaar, H.A.; Raza, M.M.; Sarfraz, S.; Tariq, I. First report of brown leaf spot of rice caused by Bipolaris zeicola in Pakistan. Plant Dis. 2020, 105, 212. [Google Scholar] [CrossRef]
- Wang, M.; Wang, S.Q.; Ma, J.; Yu, C.J.; Gao, J.X.; Chen, J. Detection of Cochliobolus heterostrophus races in South China. J. Phytopathol. 2017, 165, 681–691. [Google Scholar] [CrossRef]
- Sharma, R.C.; Rai, S.N. Evaluation of maize inbred lines for resistance to maydis leaf blight. Indian Phytopath. 2005, 58, 339–340. [Google Scholar]
- Gan, L.; Dai, Y.L.; Ruan, H.C.; Shi, N.N.; Du, Y.X.; Chen, F.R.; Yang, X.J. Pathotype and its population structure of Cochliobolus heterostrophus in Fujian Province. Chin. Agric. Sci. Bull. 2018, 34, 147–151. [Google Scholar]
- Tsukiboshi, T.; Kimigafukuro, T.; Sato, T. Identification of races of Bipolaris zeicola, the causal fungus of Helminthosporium leaf spot on corn in Japan. Ann. Phytopathol. Soc. Jpn. 1987, 53, 647–649. [Google Scholar] [CrossRef]
- Manzar, N.; Kashyap, A.S.; Sharma, P.K.; Saxena, A.K. First report of leaf spot of maize caused by Curvularia geniculata in India. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Giridharan, P.; Verekar, S.A.; Gohil, A.R.; Mishra, P.D.; Khanna, A.; Deshmukh, S.K. Antiprolif-erative activity of hamigerone and radicinol isolated from Bipolaris papendorfii. BioMed Res. Int. 2014, 2014, 890904. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Crous, P.W. Multi-locus phylogeny of the ge-nus Curvularia and description of ten new species. Mycol Progress 2020, 19, 559–588. [Google Scholar] [CrossRef]
- Sivanesan, A. Graminicolous Species of Bipolaris, Curvularia, Drechslera, Exserohilum and tTheir Teleomorphs; Mycological Papers No. 158; C.A.B. International: Wallingford, UK, 1987; pp. 1–261. [Google Scholar]
- Cui, W.L.; Lu, X.Q.; Bian, J.Y.; Qi, X.L.; Li, D.W.; Huang, L. Curvularia spicifera and Curvularia muehlenbeckiae causing leaf blight on Cunninghamia lanceolata. Plant Pathol. 2020, 69, 1139–1147. [Google Scholar] [CrossRef]
- Khemmuk, W.; Shivas, R.G.; Henry, R.J.; Geering, A.D. Fungi associated with foliar diseases of wild and cultivated rice (Oryza spp.) in northern Queensland. Australasian Plant Pathol. 2016, 45, 297–308. [Google Scholar] [CrossRef]
- Li, P.P.; Cao, Z.Y.; Dong, J.G.; Zhang, L.H.; Jia, H.; Liu, N.; Li, S.H.; Hao, Z.M.; Gu, S.Q.; Wang, X.Y. First report of Bipolaris papendorfii causing corn leaf spot in China. Plant Dis. 2013, 97, 1506–1507. [Google Scholar] [CrossRef]
- Trainor, M.J.; Martinson, C.J. Epidemiology of Alternaria leaf blight of maize (Abstr.). Phytopathology 1981, 71, 262. [Google Scholar]
- Xu, X.; Zhang, L.; Yang, X.; Cao, H.; Li, J.; Cao, P.; Guo, L.; Wang, X.; Zhao, J.; Xiang, W. Alternaria spp. associated with leaf blight of maize in Heilongjiang Province, China. Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Niu, Y.C.; Deng, H. Bipolaris and Curvularia species associated with corn leaf spot in northern China. Plant Prot. 2016, 42, 39–46. [Google Scholar]
- Guo, N.; Ni, X.; Shi, J.; Ma, J.Y.; Xue, C.S.; Chen, J. The occurrence and pathogen identification of leaf spot on maize. Acta Phytopathol. Sin. 2017, 47, 1–8. [Google Scholar]
- Acharya, K.; Dutta, A.K.; Pradhan, P. Bipolaris sorokiniana (Sacc.) Shoem.: The most destructive wheat fungal pathogen in the warmer areas. Aust. J. Crop. Sci. 2011, 5, 1064–1071. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, Systematic Mycology and Microbiology Laboratory; ARS, USDA: Beltsville, MD, USA, 2020. Available online: https://nt.arsgrin.gov/fungaldatabases (accessed on 9 November 2021).
- Singh, V.; Lakshman, D.K.; Roberts, D.P.; Ismaiel, A.; Hooda, K.S.; Gogoi, R. Morphopathological and molecular morphometric characterization of Waitea circinata var. prodigus causing a novel sheath spot disease of maize in India. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [Green Version]
- White, T.F.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocol: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, F.S., White, T.T., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A modular system for evolutionary analysis, Version 3.70. Evolution 2021. Available online: https://www.mesquiteproject.org (accessed on 9 November 2021).
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payak, M.M.; Sharma, R.C. Disease rating scales in maize in India. In Techniques of Scoring for Resistance to Important Diseases of Maize; All India Coordinated Maize Improvement Project; IARI: New Delhi, India, 1983; pp. 1–4. [Google Scholar]
- Mullen, J.M. Plant Disease Diagnosis. In Plant Pathology Concepts and Laboratory Exercises, 2nd ed.; Trigiano, R.N., Windham, M.T., Windham, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Hooda, K.S.; Bagaria, P.K.; Khokhar, M.; Kaur, H.; Rakshit, S. Mass Screening Techniques for Resistance to Maize Diseases; ICAR-Indian Institute of Maize Research, PAU Campus: Ludhiana, India, 2018. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. $ | New Code | Location | AEZ * | State | Latitude | Longitude | Soil Type | No. of Samples Collected | Accession No. | |
|---|---|---|---|---|---|---|---|---|---|---|
| ITS | GAPDH | |||||||||
| 1 | BmPhRj4 | Pichola (Udaipur) | WDR | Rajasthan | 24°36′48.00″ N | 73°40′48.00″ E | Sandy/clay loam to dessert loam | 15 | KX668613 | OL519604 |
| 2 | BmBjUa1 | Bajaura | WHR | Uttaranchal | 31°50′54.2483″ N | 77°9′51.6013″ E | Sandy to clay loam | 23 | KX668605 | OL519605 |
| 3 | BmBsRj4 | Banswara (Udaipur) | WDR | Rajasthan | 23°32′48.3252″ N | 74°26′1.7880″ E | Sandy/clay loam to dessert loam | 11 | KX668614 | OL519606 |
| 4 | BmDhBh3 | Dholi | MGP | Bihar | 25°51′25.9951″ N | 85°46′85.5895″ E | Deep loamy/silt/clay loam | 27 | KX668619 | OL519607 |
| 5 | BmCgRj4 | Chittorgarh (Udaipur) | WDR | Rajasthan | 24°54′16.5716″ N | 74°42′29.558″ E | Sandy/clay loam to dessert loam | 18 | KX668617 | OL519608 |
| 6 | BmPnDl2 | Patel Nagar | TGP | Delhi | 28°39′8.7966″ N | 77°11′29.9389″ E | Alluvium | 15 | KX668610 | OL519609 |
| 7 | BmDnRj4 | Dungarpur (Udaipur) | WDR | Rajasthan | 23°50′16.5716″ N | 73°50′29.558″ E | Sandy/clay loam to dessert loam | 19 | KX668616 | OL519610 |
| 8 | BmKgUa1 | Kangra | WHR | Uttaranchal | 32°5′59.2944″ N | 76°16′8.7744″ E | Shallow to deep loam | 21 | KX668609 | OL519611 |
| 9 | BmKrHr2 | Karnal | TGP | Haryana | 29°41′8.4944″ N | 76°59′25.737″ E | Sandy clay | 20 | KX668608 | OL519612 |
| 10 | BmKtRj4 | Kota (Udaipur) | WDR | Rajasthan | 24°10′16.5716″ N | 75°52′29.558″ E | Sandy/clay loam to dessert loam | 23 | KX668618 | OL519614 |
| 11 | BmLdPj2 | Ludhiana | TGP | Punjab | 30°54′3.474″ N | 75°51′26.1929″ E | Deep loamy/sandy/clay loam | 34 | KX668623 | OL519613 |
| 12 | BmPtUa1 | Pantnagar | WHR | Uttaranchal | 29°1′15.74″ N | 79°29′23.06″ E | Sandy clay | 18 | KX668604 | OL519615 |
| 13 | BmMyKa6 | Mysore | SPHR | Karnataka | 12°31′25.4316″ N | 76°53′40.8624″ E | Light red sandy loam | 9 | OK576634 | OL519616 |
| 14 | BmMdKa6 | Mandya | SPHR | Karnataka | 12°31′25.4316″ N | 76°53′40.8624″ E | Light red sandy loam | 13 | KX668606 | OL502169 |
| 15 | BmGdGj5 | Godhra | GPHR | Gujarat | 22°46′24.9456″ N | 73°36′49.9824″ E | Sandy loam | 12 | KX668621 | OL502170 |
| 16 | BmAdGj5 | Anand | GPHR | Gujarat | 22°33′14.5044″ N | 72°56′56.1696″ E | Sandy loam | 12 | KX668622 | OL502171 |
| 17 | BmLhRj4 | Lakhawali (Udaipur) | WDR | Rajasthan | 24°34′16.5716″ N | 73°41′29.558″ E | Sandy/clay loam to dessert loam | 11 | KX668612 | OL502172 |
| 18 | BmSkRj4 | Sukher (Udaipur) | WDR | Rajasthan | 24°34′16.5716″ N | 73°41′29.558″ E | Sandy/clay loam to dessert loam | 16 | KX668615 | OL519603 |
| 19 | BmSmBh3 | Samastipur | MGP | Bihar | 25°51′46.6848″ N | 85°46′51.7044″ E | Deep loamy/silt/clay loam | 18 | KX668620 | OL519617 |
| 20 | BmAmRj4 | Amberi (Udaipur) | WDR | Rajasthan | 26°55′16.5716″ N | 73°50′29.558″ E | Sandy/clay loam to dessert loam | 15 | KX668611 | ** |
| S N $ | Isolate | Size of Conidia (µm) and Septations * | Radial Growth (in mm) ** | Disease Score *** | Incubation Period (in h) ∞ | Colony Texture # | Colour of Colony ** | Disease Index β (PDI) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Length (µm) | Width (µm) | No. of Septa | ||||||||
| 1 | BmPhRj4 | 9.8 (6.54–17.86) | 3.4 (2.25–4.63) | 2.7 (2–5) | 47.5 (++) | 1.5 | 72 (Type-I) | Rr/Nz/Irm | Light black | 30 (MV) |
| 2 | BmBjUa1 | 16.9 (10.68–20.84) | 4.1 (3.72–4.45) | 7.1 (5–8) | 58.5 (++) | 2.6 | 72 (Type-II) | Rr/Nz/Rm | Black | 52 (MoV) |
| 3 | BmBsRj4 | 12.7 (7.79–22.78) | 4.9 (4.36–5.26) | 4.7 (3–6) | 71.4 (+++) | 2.3 | 48 (Type-III) | Sap/Nz/Rm | Black | 46 (MV) |
| 4 | BmDhBh3 | 14.4 (11.91–16.66) | 4.5 (3.91–5.43) | 3.7 (3–5) | 51.1 (++) | 3.7 | 72 (Type-I) | Rap/Nz/Rm | Black | 74 (HV) |
| 5 | BmCgRj4 | 14.8 (9.26–20.96) | 4.3 (2.48–4.78) | 4.2 (2–9) | 72.3 (+++) | 2.9 | 48 (Type-II) | Rr/Nz/Rm | Light black | 58 (MoV) |
| 6 | BmPnDl2 | 19.3 (13.27–21.36) | 5.0 (3.91–5.73) | 4.3 (3–7) | 68.8 (++) | 2.7 | 72 (Type-III) | Rr/Z/Irm | Black | 54 (MoV) |
| 7 | BmDnRj4 | 11.1 (6.52–14.94) | 4.7 (3.63–5.51) | 3.0 (2–5) | 69.0 (++) | 1.9 | 48 (Type-II) | Sap/Nz/Rm | Light grey | 38 (MV) |
| 8 | BmKgUa1 | 10.1 (7.34–15.43) | 4.5 (3.89–5.46) | 3.1 (2–6) | 63.4 (++) | 2.4 | 76 (Type-II) | Sap/Nz/Rm | Dark grey | 48 (MV) |
| 9 | BmKrHr2 | 15.1 (19.25–11.03) | 4.4 (4.19–4.93) | 6.1 (4–7) | 74.3 (+++) | 3.5 | 48 (Type-I) | Sap/Nz/Rm | Dark grey | 70 (HV) |
| 10 | BmKtRj4 | 13.0 (8.92–19.4) | 3.9 (2.54–5.22) | 3.7 (2–5) | 54.3 (++) | 1.7 | 96 (Type-III) | Sap/Nz/Rm | Black | 34 (MV) |
| 11 | BmLdPj2 | 14.3 (8.78–23.57) | 4.4 (2.55–5.77) | 5.1 (3–6) | 37.4 (+) | 4.3 | 72 (Type-II) | Rr/Nz/Irm | Dark grey | 86 (HV) |
| 12 | BmPtUa1 | 14.7 (11.89–17.74) | 4.9 (4.23–5.43) | 4.1 (3–4) | 45.5 (++) | 3.2 | 72 (Type-II) | Rr/Nz/Rm | Dark grey | 64 (MoV) |
| 13 | BmMyKa6 | 13.1 (10.77–17.03) | 3.8 (2.46–4.78) | 4.1 (3–4) | 78.9 (+++) | 2.7 | 72 (Type-IV) | Sap/Z/Rm | Light grey | 54 (MoV) |
| 14 | BmMdKa6 | 11.6 (6.88–15.32) | 3.6 (4.12–5.36) | 3.8 (4–2) | 81.8 (+++) | 1.7 | 72 (Type-I) | Sap/Z/Rm | Light grey | 34 (MV) |
| 15 | BmGdGj5 | 7.6 (6.83–9.77) | 3.1 (3.61–4.52) | 1.0 (1–3) | 79.3 (+++) | 3.3 | 48 (Type-I) | Rr/Nz/Rm | Black | 66 (MoV) |
| 16 | BmAdGj5 | 12.1 (8.23–15.98) | 4.3 (3.84–5.86) | 3.0 (2–9) | 51.6 (++) | 4.5 | 72 (Type-VI) | Rap/Nz/Rm | Black | 90 (HV) |
| 17 | BmLhRj4 | 17.2 (10.33–23.13) | 3.7 (2.44–4.57) | 5.0 (4–8) | 59.3 (++) | 3.7 | 72 (Type-III) | Rr/Z/Irm | Dark grey | 74 (HV) |
| 18 | BmSkRj4 | 16.8 (9.78–20.11) | 3.1 (2.98–4.91) | 3.0 (2–8) | 37.7 (+) | 3.6 | 72 (Type-V) | Sap/Nz/Rm | Dark grey | 72 (HV) |
| 19 | BmSmBh3 | 7.8 (6.12–10.11) | 6.1 (4.21–7.93) | 2.0 (2–6) | 42.6 (++) | 4.3 | 76 (Type-V) | Sap/Z/Rm | Dark grey | 86 (HV) |
| 20 | BmAmRj4 | 18.6 (7.33–21.67) | 5.8 (3.22–8.44) | 6.0 (4–8) | 67.3 (++) | 4.1 | 72 (Type-IV) | Rr/Nz/Irm | Black | 82 (HV) |
| CD at 5% | 6.2 | 1.2 | 2.0 | 1.28 | 1.1 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, V.; Lakshman, D.K.; Roberts, D.P.; Ismaiel, A.; Abhishek, A.; Kumar, S.; Hooda, K.S. Fungal Species Causing Maize Leaf Blight in Different Agro-Ecologies in India. Pathogens 2021, 10, 1621. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121621
Singh V, Lakshman DK, Roberts DP, Ismaiel A, Abhishek A, Kumar S, Hooda KS. Fungal Species Causing Maize Leaf Blight in Different Agro-Ecologies in India. Pathogens. 2021; 10(12):1621. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121621
Chicago/Turabian StyleSingh, Vimla, Dilip K. Lakshman, Daniel P. Roberts, Adnan Ismaiel, Alok Abhishek, Shrvan Kumar, and Karambir S. Hooda. 2021. "Fungal Species Causing Maize Leaf Blight in Different Agro-Ecologies in India" Pathogens 10, no. 12: 1621. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121621