
Differences in Genotype and Antimicrobial Resistance between Campylobacter spp. Isolated from Organic and Conventionally Produced Chickens in Sweden

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Antibiotic Resistance
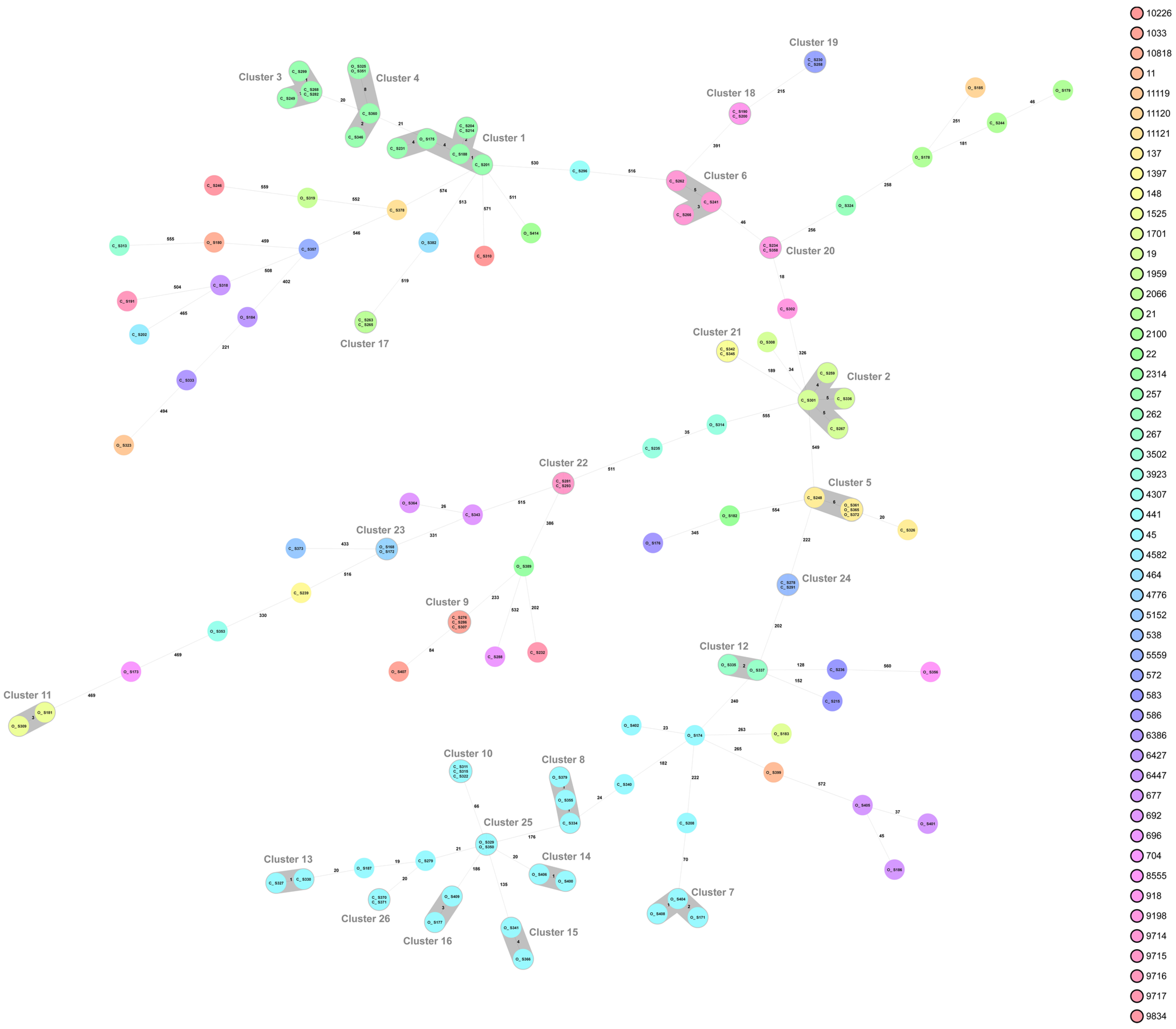
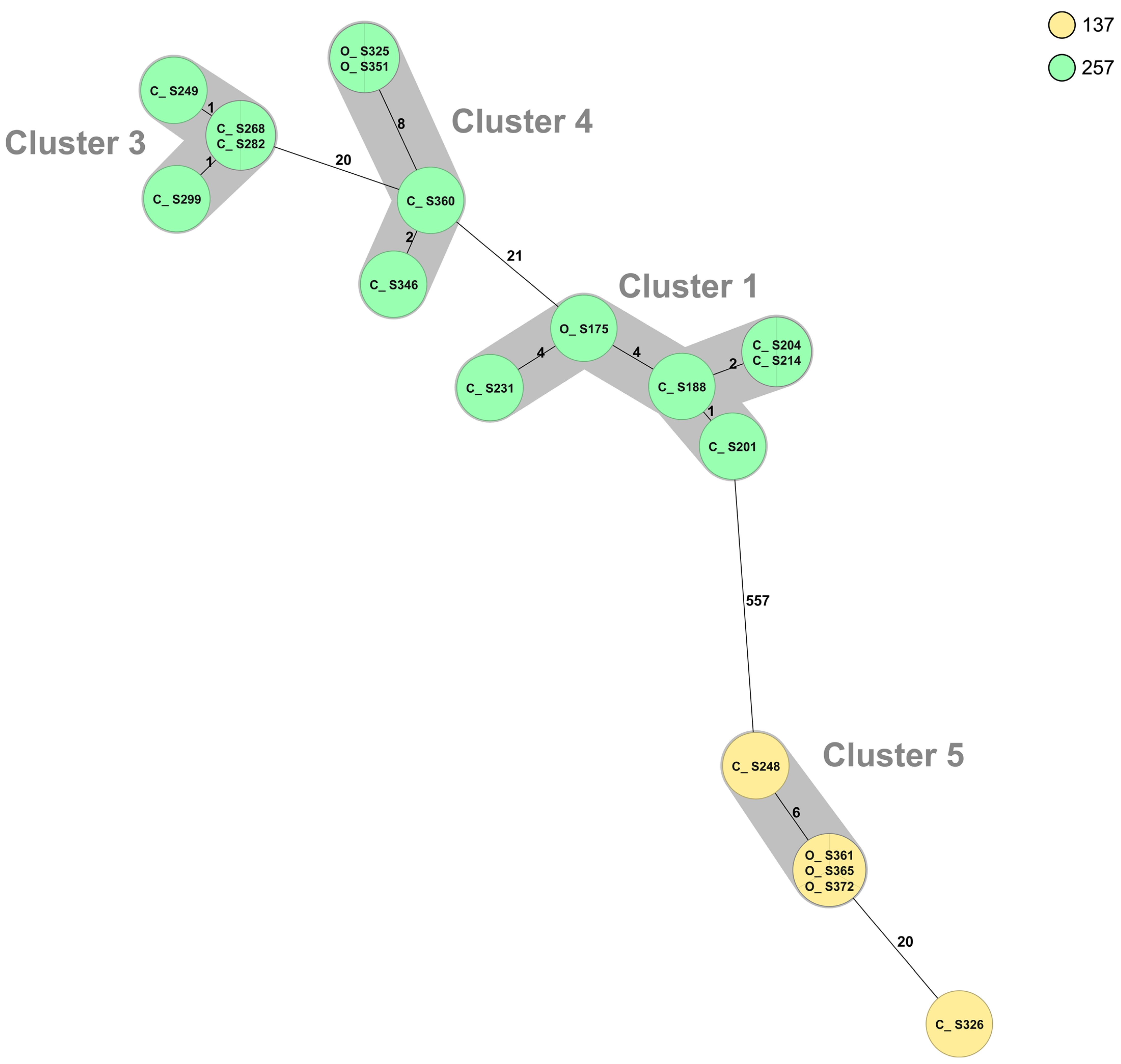
2.2. Campylobacter Genotypes Determined by Whole-Genome Sequencing
3. Discussion
4. Materials and Methods
4.1. Broiler Population
4.2. Sampling and Bacteriological Analysis
4.3. Antibiotic Susceptibility Testing
4.4. Whole-Genome Sequencing
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NNDSS, Annual Report Working Group. Australia’s notifiable disease status, 2014: Annual report of the National Notifiable Diseases Surveillance System. Commun. Dis. Intell. 2016, 40, E48–E145. [Google Scholar]
- Geissler, A.L.; Bustos Carrillo, F.; Swanson, K.; Patrick, M.E.; Fullerton, K.E.; Bennett, C.; Barrett, K.; Mahon, B.E. Increasing Campylobacter Infections, Outbreaks, and Antimicrobial Resistance in the United States, 2004–2012. Clin. Infect. Dis. 2017, 65, 1624–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Institute of Environmental Science and Research Ltd. (ESR). Notifiable Diseases in New Zealand; Annual Report. 2020. Available online: www.surv.esr.cri.nz (accessed on 28 June 2021).
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 286.
- Center for Disease Control and Prevention (CDC). Campylobacter (Campylobacteriosis). 2020. Available online: https://www.cdc.gov/campylobacter/faq.html (accessed on 28 June 2021).
- Hansson, I.; Gustafsson, P.; Hellquist, B.; Lahti, E.; Pudas, N.; Olsson Engvall, E. Campylobacter in Swedish small scale chicken production. A comparison with findings in conventionally produced Swedish and European broilers. In Proceedings of the CHRO, 17th International Workshop on Campylobacter, Helicobacter and Related Organisms, Aberdeen, Scotland, 15–20 September 2013. [Google Scholar]
- Heuer, O.E.; Pedersen, K.; Andersen, J.S.; Madsen, M. Prevalence and antimicrobial susceptibility of thermophilic Campylobacter in organic and conventional broiler flocks. Lett. Appl. Microbiol. 2001, 33, 269–274. [Google Scholar] [CrossRef]
- Allen, V.M.; Ridley, A.M.; Harris, J.A.; Newell, D.G.; Powell, L. Influence of production system on the rate of onset of Campylobacter colonization in chicken flocks reared extensively in the United Kingdom. Br. Poult. Sci. 2011, 52, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Rosenquist, H.; Boysen, L.; Krogh, A.L.; Jensen, A.N.; Nauta, M. Campylobacter contamination and the relative risk of illness from organic broiler meat in comparison with conventional broiler meat. Int. J. Food Microbiol. 2013, 162, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Golden, C.E.; Mishra, A. Prevalence of Salmonella and Campylobacter spp. in Alternative and Conventionally Produced Chicken in the United States: A Systematic Review and Meta-Analysis. J. Food Prot. 2020, 83, 1181–1197. [Google Scholar] [CrossRef]
- Bouwknegt, M.; van de Giessen, A.W.; Dam-Deisz, W.D.; Havelaar, A.H.; Nagelkerke, N.J.; Henken, A.M. Risk factors for the presence of Campylobacter spp. in Dutch broiler flocks. Prev. Vet. Med. 2004, 62, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Näther, G.; Alter, T.; Martin, A.; Ellerbroek, L. Analysis of risk factors for Campylobacter species infection in broiler flocks. Poult. Sci. 2009, 88, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Sommer, H.M.; Heuer, O.E.; Sørensen, A.I.; Madsen, M. Analysis of factors important for the occurrence of Campylobacter in Danish broiler flocks. Prev. Vet. Med. 2013, 111, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Government Offices of Sweden Homepage. 2018. Available online: https://www.regeringen.se/pressmeddelanden/2018/04/jordbruksverket-ska-jobba-for-okad-ekologisk-produktion (accessed on 1 April 2021).
- World Health Organization (WHO). WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. 2000. Available online: https://www.who.int/news-room/detail/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 28 May 2020).
- Bengtsson, B.; Wierup, M. Antimicrobial resistance in Scandinavia after ban of antimicrobial growth promoters. Anim. Biotechnol. 2006, 17, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.L.; Caffrey, N.P.; Nóbrega, D.B.; Cork, S.C.; Ronksley, P.E.; Barkema, H.W.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.D.; et al. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: A systematic review and meta-analysis. Lancet Planet Health 2017, 8, e316–e327. [Google Scholar] [CrossRef]
- Varga, C.; Guerin, M.T.; Brash, M.L.; Slavic, D.; Boerlin, P.; Susta, L. Antimicrobial resistance in Campylobacter jejuni and Campylobacter coli isolated from small poultry flocks in Ontario, Canada: A two-year surveillance study. PLoS ONE 2019, 14, e0221429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wierup, M.; Wahlström, H.; Bengtsson, B. Successful Prevention of Antimicrobial Resistance in Animals-A Retrospective Country Case Study of Sweden. Antibiotics 2021, 10, 129. [Google Scholar] [CrossRef]
- Swedres-Svarm. Sales of Antibiotics and Occurrence of Resistance in Sweden; Public Health Agency and National Veterinary Institute: Solna, Sweden, 2020. [Google Scholar]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2017/2018. EFSA J. 2020, 18, e06007. [Google Scholar]
- Hansson, I.; Olsson Engvall, E.; Lahti, E.; Landen, A.; Harbom, B.; Bengtsson, B. Antimicrobial resistance of Campylobacter in Swedish chicken. In Proceedings of the CHRO, 18th International Workshop on Campylobacter, Helicobacter and Related Organisms, Rotorua, New Zealand, 1–5 November 2015. [Google Scholar]
- Frosth, S.; Karlsson-Lindsjö, O.; Niazi, A.; Fernström, L.L.; Hansson, I. Identification of Transmission Routes of Campylobacter and On-Farm Measures to Reduce Campylobacter in Chicken. Pathogens 2020, 9, 363. [Google Scholar] [CrossRef]
- Luangtongkum, T.; Morishita, T.Y.; Ison, A.J.; Huang, S.; McDermott, P.F.; Zhang, Q. Effect of conventional and organic production practices on the prevalence and antimicrobial resistance of Campylobacter spp. in poultry. Appl. Environ. Microbiol. 2006, 72, 3600–3607. [Google Scholar] [CrossRef] [Green Version]
- Tenhagen, B.A.; Alt, K.; Käsbohrer, A.; Kollas, C.; Pfefferkorn, B.; Naumann, S.; Wiehle, L.; Thieck, M.; Stingl, K. Comparison of Antimicrobial Resistance of Thermophilic Campylobacter Isolates from Conventional and Organic Turkey Meat in Germany. Foodborne Pathog. Dis. 2020, 17, 750–757. [Google Scholar] [CrossRef]
- Innes, G.K.; Nachman, K.E.; Abraham, A.G.; Casey, J.A.; Patton, A.N.; Price, L.B.; Tartof, S.Y.; Davis, M.F. Contamination of Retail Meat Samples with Multidrug-Resistant Organisms in Relation to Organic and Conventional Production and Processing: A Cross-Sectional Analysis of Data from the United States National Antimicrobial Resistance Monitoring System, 2012–2017. Environ. Health Perspect. 2021, 129, 57004. [Google Scholar] [PubMed]
- Engberg, J.; Neimann, J.; Nielsen, E.M.; Aarestrup, F.M.; Fussing, V. Quinolone-resistant Campylobacter infections: Risk factors and clinical consequences. Emerg. Infect. Dis. 2004, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, M.; Salata, C.; Martini, M.; Montesissa, C.; Piccirillo, A. Antimicrobial resistance of Campylobacter jejuni and Campylobacter coli from poultry in Italy. Microb. Drug Resist. 2014, 20, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Jacobs-Reitsma, W.F.; Koenraad, P.M.; Bolder, N.M.; Mulder, R.W. In vitro susceptibility of Campylobacter and Salmonella isolates from broilers to quinolones, ampicillin, tetracycline, and erythromycin. Vet. Q. 1994, 16, 206–208. [Google Scholar] [CrossRef]
- Velàzquez, J.B.; Jimenez, A.; Chomon, B.; Villa, T.G. Incidence and transmission of antibiotic resistance in Campylobacter jejuni and Campylobacter coli. J. Antimicrob. Chemother. 1995, 35, 173–178. [Google Scholar] [CrossRef]
- Maxwell, A. The molecular basis of quinolone action. J. Antimicrob. Chemother. 1992, 30, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Huang, W.M.; Taylor, D.E. Cloning and nucleotide sequence of the Campylobacter jejuni gyrA gene and characterization of quinolone resistance mutations. Antimicrob. Agents Chemother. 1993, 37, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payot, S.; Bolla, J.M.; Corcoran, D.; Fanning, S.; Mégraud, F.; Zhang, Q. Mechanisms of fluoroquinolone and macrolide resistance in Campylobacter spp. Microb. Infect. 2006, 8, 1967–1971. [Google Scholar] [CrossRef]
- Haldenby, S.; Bronowski, C.; Nelson, C.; Kenny, J.; Martinez-Rodriguez, C.; Chaudhuri, R.; Williams, N.J.; Forbes, K.; Strachan, N.J.; Pulman, J.; et al. Increasing prevalence of a fluoroquinolone resistance mutation amongst Campylobacter jejuni isolates from four human infectious intestinal disease studies in the United Kingdom. PLoS ONE 2020, 15, e0227535. [Google Scholar] [CrossRef] [Green Version]
- Hansson, I.; Tamminen, L.M.; Frosth, S.; Fernström, L.L.; Emanuelson, U.; Boqvist, S. Occurrence of Campylobacter spp. in Swedish calves, common sequence types and antibiotic resistance patterns. J. Appl. Microbiol. 2020, 130, 2111–2122. [Google Scholar] [CrossRef] [PubMed]
- Kovač, J.; Šimunović, K.; Wu, Z.; Klančnik, A.; Bucar, F.; Zhang, Q.; Možina, S.S. Antibiotic resistance modulation and modes of action of (-)-α-pinene in Campylobacter jejuni. PLoS ONE 2015, 1, e0122871. [Google Scholar] [CrossRef] [Green Version]
- Hakkinen, M.; Heiska, H.; Hänninen, M.L. Prevalence of Campylobacter spp. in cattle in Finland and antimicrobial susceptibilities of bovine Campylobacter jejuni strains. Appl. Environ. Microbiol. 2007, 73, 3232–3238. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.A.; Taylor, R.M.; Brar, J.S.; Corkran, S.C.; Velásquez, C.; Novoa Rama, E.; Oliver, H.F.; Singh, M. Prevalence and antimicrobial resistance of Campylobacter from antibiotic-free broilers during organic and conventional processing. Poult. Sci. 2019, 98, 1447–1454. [Google Scholar] [CrossRef]
- Pedersen, K.; Wedderkopp, A. Resistance to quinolones in Campylobacter jejuni and Campylobacter coli from Danish broilers at farm level. J. Appl. Microbiol. 2003, 94, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, O.; Greko, C.; Bengtsson, B.; Englund, S. Genetic diversity among VRE isolates from Swedish broilers with the coincidental finding of transferrable decreased susceptibility to narasin. J. Appl. Microbiol. 2012, 112, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Avrain, L.; Humbert, F.; L’Hospitalier, R.; Sanders, P.; Vernozy-Rozand, C.; Kempf, I. Antimicrobial resistance in Campylobacter from broilers: Association with production type and antimicrobial use. Vet. Microbiol. 2003, 96, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Willer, T.; Pielsticker, C.; Gerzova, L.; Rychlik, I.; Rautenschlein, S. Differences in host breed and diet influence colonization by Campylobacter jejuni and induction of local immune responses in chicken. Gut Pathog. 2016, 8, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KRAV. Available online: https://www.krav.se/om-krav/krav-markningen/djur/sa-blir-kycklingen-krav-certifierad/ (accessed on 21 March 2020).
- Rezaei, M.; Yngvesson, J.; Gunnarsson, S.; Jönsson, L.; Wallenbeck, A. Feed efficiency, growth performance, and carcass characteristics of a fast-and a slower-growing broiler hybrid fed low-or high-protein organic diets. Org. Agric. 2018, 8, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Hansson, I.; Forshell, L.P.; Gustafsson, P.; Boqvist, S.; Lindblad, J.; Engvall, E.O.; Andersson, Y.; Vågsholm, I. Summary of the Swedish Campylobacter program in broilers, 2001 through 2005. J. Food Prot. 2007, 70, 2008–2014. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; CLSI standard VET01 Ed5; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Souvorov, A.; Agarwala, R.; Lipman, D.J. SKESA: Strategic k-mer extension for scrupulous assemblies. Genome Biol. 2018, 19, 153. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Nennig, M.; Llarena, A.-K.; Herold, M.; Mossong, J.; Penny, C.; Losch, S.; Tresse, O.; Ragimbeau, S. Investigating major recurring Campylobacter jejuni lineages in Luxembourg using four core or whole genome sequencing typing schemes. Front. Cell. Infect. Microbiol. 2021, 10, 608020. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2020, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
 |
 |
| Total | Organic | Conventional | ||||
|---|---|---|---|---|---|---|
| CC | MLST | No. of Isolates | No. of Isolates | Resistance | No. of Isolates | Resistance 1 |
| 21 | 19 | 5 | 1 | - | 4 | 4 (cip + nal) |
| 21 | 21 | 3 | 2 | - | 1 | - |
| 21 | 148 | 2 | - | - | 2 | 2 (cip + nal) |
| 21 | 262 | 1 | 1 | - | - | - |
| 21 | 7419 | 3 | - | - | 3 | 3 (cip + nal + tet) |
| 21 | 9198 | 3 | - | - | 3 | - |
| 21 | 11120 | 1 | 1 | - | - | - |
| 22 | 22 | 1 | 1 | - | - | - |
| 45 | 11 | 1 | 1 | - | - | - |
| 45 | 45 | 27 | 16 | - | 11 | - |
| 45 | 137 | 5 | 3 | - | 2 | - |
| 45 | 538 | 3 | - | - | 3 | - |
| 45 | 583 | 2 | - | - | 2 | - |
| 45 | 1701 | 1 | 1 | - | - | - |
| 48 | 918 | 2 | - | - | 2 | - |
| 52 | 2066 | 2 | - | - | 2 | - |
| 52 | 2100 | 1 | 1 | 1 (tet) | - | - |
| 206 | 572 | 2 | - | - | 2 | 2 (cip + nal) |
| 257 | 257 | 14 | 3 | - | 11 | 2 (cip + nal) |
| 283 | 267 | 2 | 2 | - | - | - |
| 464 | 464 | 1 | 1 | 1 (tet) | - | - |
| 677 | 677 | 3 | 3 | - | - | - |
| 692 | 692 | 2 | 1 | - | 1 | - |
| 692 | 4776 | 2 | 2 | - | - | - |
| 702 | 5152 | 1 | - | - | 1 | - |
| 952 | 4582 | 1 | - | - | 1 | - |
| 952 | 6447 | 1 | - | - | 1 | - |
| 952 | 9716 | 1 | - | - | 1 | - |
| 1034 | 1033 | 4 | 1 | - | 3 | - |
| 1034 | 1034 | 1 | - | - | 1 | 1 (cip + nal) |
| 1034 | 2314 | 1 | 1 | - | - | - |
| 1034 | 9715 | 2 | - | - | 2 | - |
| 1332 | 696 | 1 | - | - | 1 | - |
| NA | 441 | 1 | - | - | 1 | 1 (cip + nal + tet) |
| NA | 586 | 1 | 1 | - | - | - |
| NA | 704 | 1 | 1 | - | - | - |
| NA | 1397 | 1 | - | - | 1 | - |
| NA | 1525 | 2 | 2 | - | - | - |
| NA | 1959 | 1 | 1 | - | - | - |
| NA | 3502 | 1 | - | - | 1 | - |
| NA | 3923 | 2 | 1 | - | 1 | - |
| NA | 4307 | 1 | 1 | - | - | - |
| NA | 5559 | 1 | - | - | 1 | - |
| NA | 6386 | 1 | - | - | 1 | - |
| NA | 6427 | 1 | 1 | - | - | - |
| NA | 8555 | 1 | 1 | - | - | - |
| NA | 9834 | 1 | - | -- | 1 | - |
| NA | 10226 | 1 | - | - | 1 | - |
| NA | 10818 | 1 | 1 | - | - | - |
| NA | 11119 | 1 | 1 | - | - | - |
| NA | 11121 | 1 | - | - | 1 | - |
| Total | Organic | Conventional | ||||
|---|---|---|---|---|---|---|
| CC | MLST | No. of Isolates | No. of Isolates | Resistant | No. of Isolates | Resistant |
| 82 | 829 | 19 | 17 | - | 2 | - |
| 82 | 855 | 8 | 8 | - | - | - |
| 82 | 1142 | 1 | - | - | 1 | - |
| 82 | 1544 | 1 | - | - | 1 | 1 (strept) |
| 82 | 2178 | 1 | - | - | 1 | 1 (cip + nal) |
| 82 | 4709 | 3 | - | - | 3 | - |
| Conventional | Organic | |
|---|---|---|
| Chicken producers within the Swedish Campylobacter program | 110 | 15 |
| Chickens produced (2017/2018/2019) | 100/99/103 million | 855,000/660,000/720,000 |
| Chickens in one compartment | Up to 60,000 | Up to 4800 |
| Maximum stocking density (kg/m2) | 36 kg | 20 kg |
| Maximum stocking number (chickens/m2) | 25 | 10 |
| Age of slaughter | 28–35 days * | 60–70 days |
| Outdoor access | Not at all | May to Sep, >4 m2/chicken |
| Breed | Ross and Cobb | Hubbard, Rowan Ranger |
| Coccidiostats | Narasin until 3 d before slaughter | Not at all |
| Campylobacter status (2017/2018/2019) | 11%/9%/5% | 40%/38%/57% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hansson, I.; Ellström, P.; Nilsson, O.; Chaba, M.; Skarin, M.; Fernström, L.-L.; Frosth, S. Differences in Genotype and Antimicrobial Resistance between Campylobacter spp. Isolated from Organic and Conventionally Produced Chickens in Sweden. Pathogens 2021, 10, 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121630
Hansson I, Ellström P, Nilsson O, Chaba M, Skarin M, Fernström L-L, Frosth S. Differences in Genotype and Antimicrobial Resistance between Campylobacter spp. Isolated from Organic and Conventionally Produced Chickens in Sweden. Pathogens. 2021; 10(12):1630. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121630
Chicago/Turabian StyleHansson, Ingrid, Patrik Ellström, Oskar Nilsson, Matilda Chaba, Moa Skarin, Lise-Lotte Fernström, and Sara Frosth. 2021. "Differences in Genotype and Antimicrobial Resistance between Campylobacter spp. Isolated from Organic and Conventionally Produced Chickens in Sweden" Pathogens 10, no. 12: 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121630