
Anti-Leishmania infantum Antibody-Producing Plasma Cells in the Spleen in Canine Visceral Leishmaniasis

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. General Characteristics of the Dogs Used in the Study
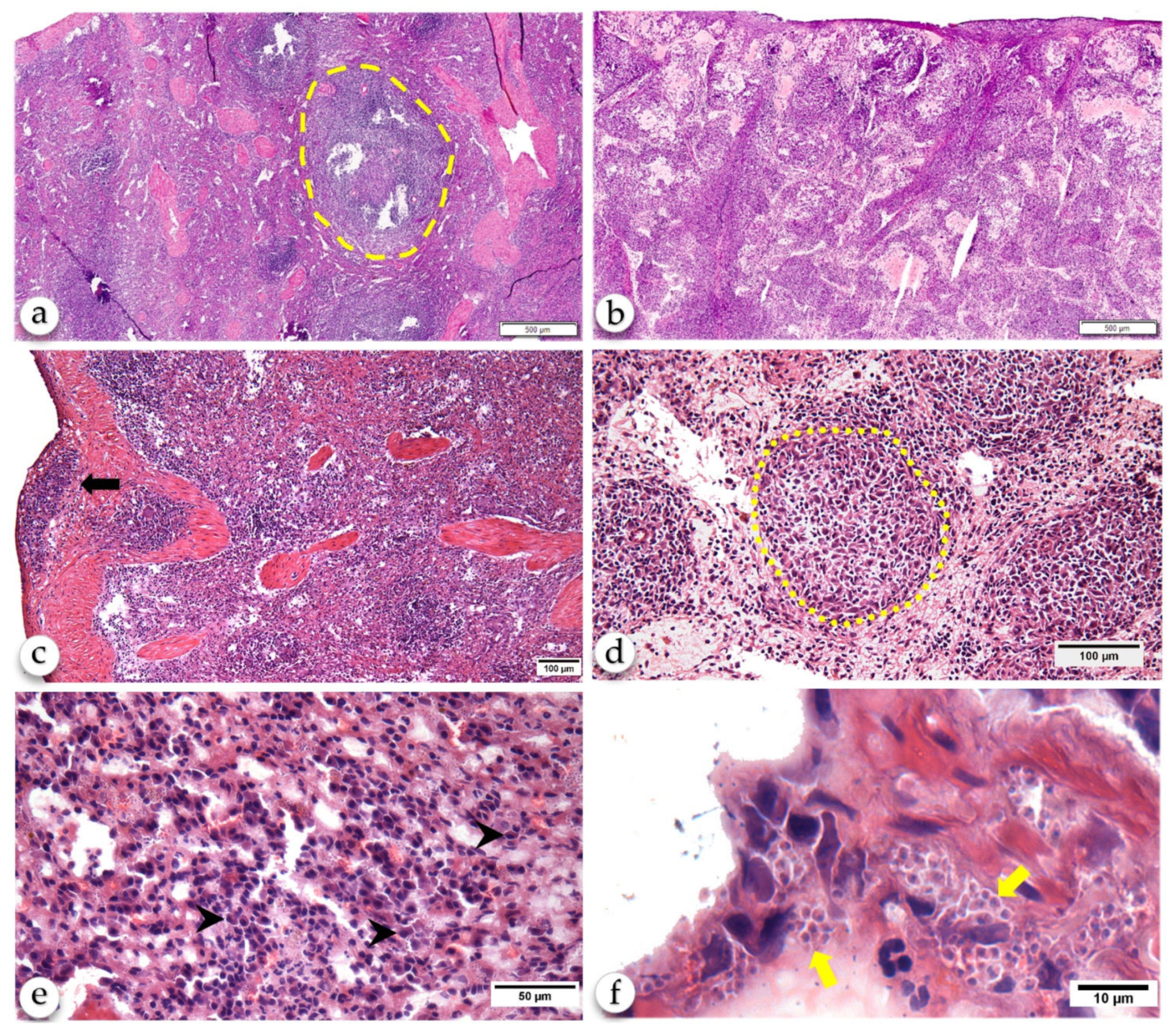
2.2. Histological Changes of the Spleen
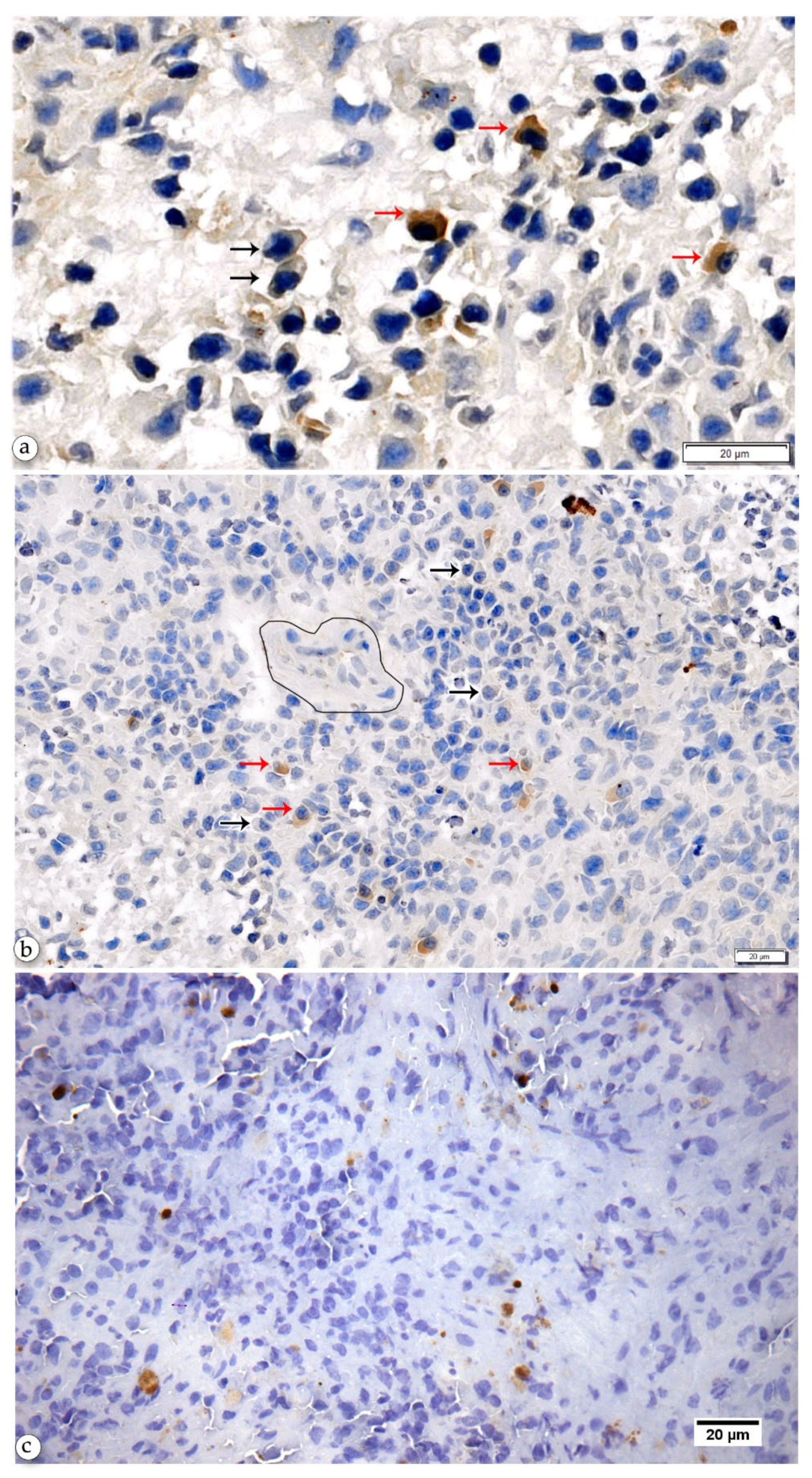
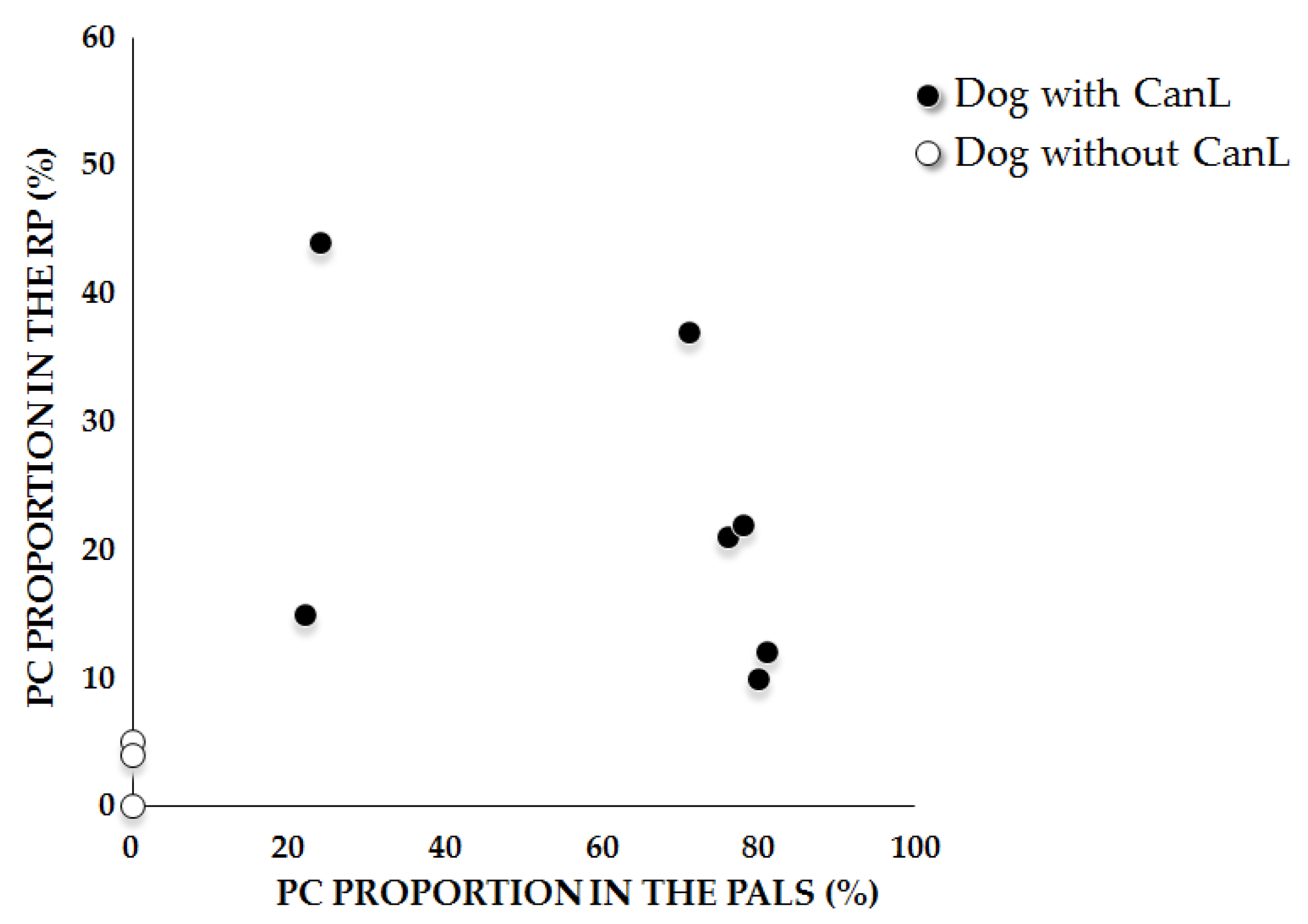
2.3. Anti-Leishmania-Specific Antibody-Producing Cells in the Spleen
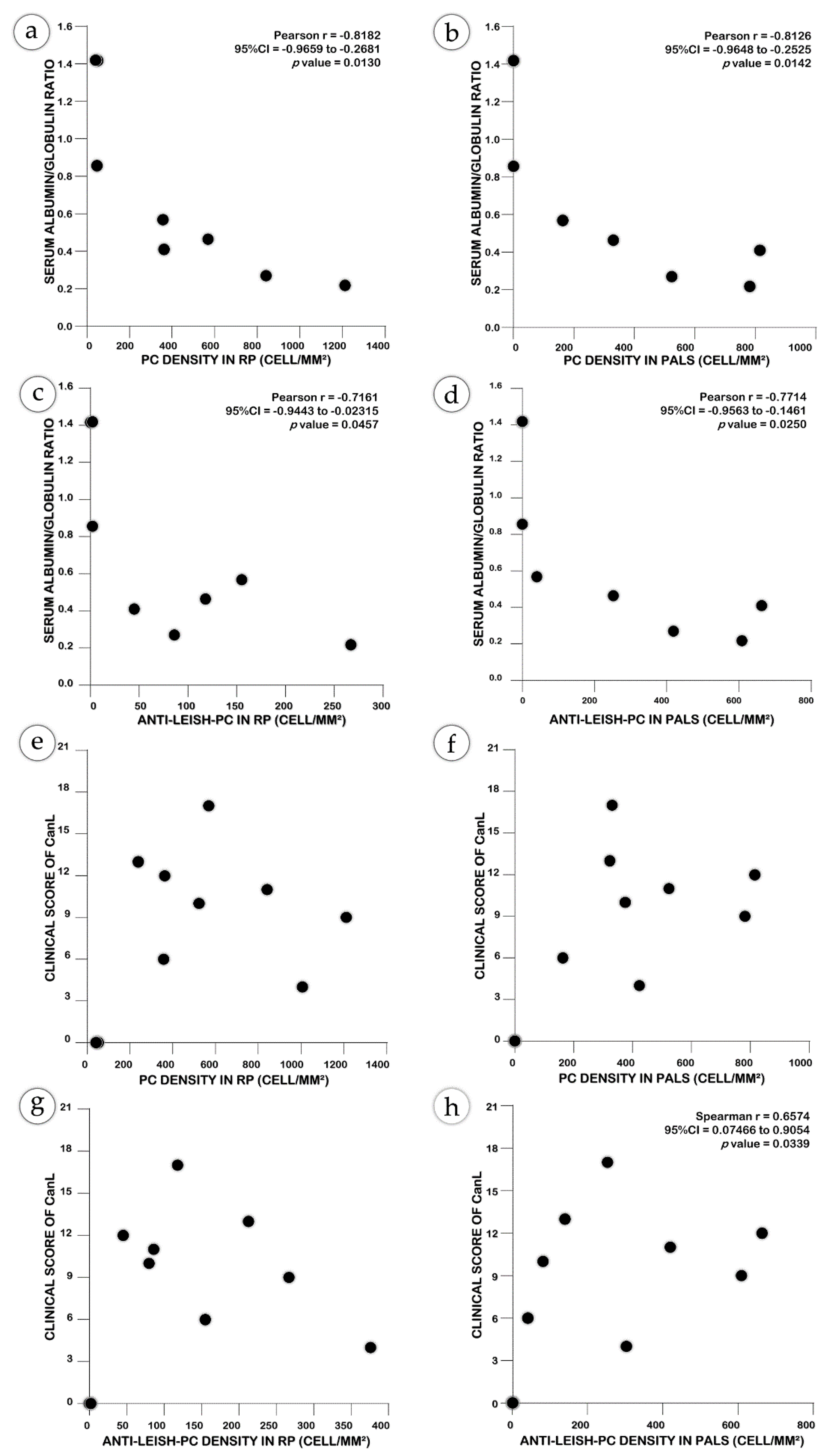
2.4. Non-Specific Plasma Cells may Contribute to the Severity of CanL
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Animal and Specimens
4.3. Production of Biotinylated Soluble Leishmania Membrane Antigen (bSLMA)
4.4. Modified Immunohistochemistry
4.5. Histological Analysis and Cell Quantification
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Health Organization Control of the Leishmaniases. In World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Ready, P. Epidemiology of visceral leishmaniasis. Clin. Epidemiol. 2014, 6, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermida, M.D.-R.; De Melo, C.V.B.; Lima, I.D.S.; Oliveira, G.G.D.S.; Dos-Santos, W.L.C. Histological Disorganization of Spleen Compartments and Severe Visceral Leishmaniasis. Front. Cell. Infect. Microbiol. 2018, 8, 394. [Google Scholar] [CrossRef] [Green Version]
- Lima, I.; Silva, J.S.; Almeida, V.A.; Junior, F.G.L.; Souza, P.A.; Larangeira, D.F.; Moura-Neto, J.P.; Fraga, D.B.; De Freitas, L.A.R.; Dos-Santos, W.L. Severe Clinical Presentation of Visceral Leishmaniasis in Naturally Infected Dogs with Disruption of the Splenic White Pulp. PLoS ONE 2014, 9, e87742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, A.S.; Ribeiro-Alves, M.; Pereira, L.D.O.R.; Mestre, G.L.; Ferreira, A.B.R.; Morgado, F.; Boité, M.C.; Cupolillo, E.; Moraes, M.O.; Porrozzi, R. Parasite Load Induces Progressive Spleen Architecture Breakage and Impairs Cytokine mRNA Expression in Leishmania infantum-Naturally Infected Dogs. PLoS ONE 2015, 10, e0123009. [Google Scholar] [CrossRef] [PubMed]
- Abbehusen, M.M.C.; Almeida, V.D.A.; Solcà, M.D.S.; Pereira, L.D.S.; Costa, D.; Gil-Santana, L.; Bozza, P.; Fraga, D.B.M.; Veras, P.S.T.; Dos-Santos, W.L.C.; et al. Clinical and immunopathological findings during long term follow-up in Leishmania infantum experimentally infected dogs. Sci. Rep. 2017, 7, 15914. [Google Scholar] [CrossRef] [Green Version]
- De Melo, C.V.B.; Hermida, M.D.-R.; Mesquita, B.R.; Fontes, J.L.M.; Koning, J.J.; Solcà, M.D.S.; Benevides, B.B.; Mota, G.B.S.; Freitas, L.A.R.; Mebius, R.E.; et al. Phenotypical Characterization of Spleen Remodeling in Murine Experimental Visceral Leishmaniasis. Front. Immunol. 2020, 11, 653. [Google Scholar] [CrossRef] [PubMed]
- Nutt, S.L.; Hodgkin, P.D.; Tarlinton, D.M.; Corcoran, L.M. The generation of antibody-secreting plasma cells. Nat. Rev. Immunol. 2015, 15, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Silva-O’Hare, J.; De Oliveira, I.S.; Klevorn, T.; Almeida, V.A.; Oliveira, G.G.D.S.; Atta, A.M.; De Freitas, L.A.R.; Dos-Santos, W.L.C. Disruption of Splenic Lymphoid Tissue and Plasmacytosis in Canine Visceral Leishmaniasis: Changes in Homing and Survival of Plasma Cells. PLoS ONE 2016, 11, e0156733. [Google Scholar] [CrossRef] [PubMed]
- Silva-Barrios, S.; Smans, M.; Duerr, C.U.; Qureshi, S.T.; Fritz, J.H.; Descoteaux, A.; Stäger, S. Innate Immune B Cell Activation by Leishmania donovani Exacerbates Disease and Mediates Hypergammaglobulinemia. Cell Rep. 2016, 15, 2427–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omachi, S.; Fujii, W.; Azuma, N.; Morimoto, A.; Sanjoba, C.; Matsumoto, Y.; Goto, Y. B-cell activating factor deficiency suppresses splenomegaly during Leishmania donovani infection. Biochem. Biophys. Res. Commun. 2017, 489, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Deak, E.; Jayakumar, A.; Cho, K.W.; Goldsmith-Pestana, K.; Dondji, B.; Lambris, J.; McMahon-Pratt, D. Murine visceral leishmaniasis: IgM and polyclonal B-cell activation lead to disease exacerbation. Eur. J. Immunol. 2010, 40, 1355–1368. [Google Scholar] [CrossRef] [PubMed]
- Giunchetti, R.C.; Mayrink, W.; Carneiro, C.M.; Correa-Oliveira, R.; Martins-Filho, O.A.; Marques, M.J.; Tafuri, W.L.; Reis, A.B. Histopathological and immunohistochemical investigations of the hepatic compartment associated with parasitism and serum biochemical changes in canine visceral leishmaniasis. Res. Veter. Sci. 2008, 84, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Barrios, S.; Stäger, S. Hypergammaglobulinemia sustains the development of regulatory responses during chronic Leishmania donovani infection in mice. Eur. J. Immunol. 2019, 49, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Zhuang, H.; Shumyak, S.; Yang, L.; Reeves, W.H. Mechanisms of Autoantibody Production in Systemic Lupus Erythematosus. Front. Immunol. 2015, 6, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogarth, P. Fc receptors are major mediators of antibody based inflammation in autoimmunity. Curr. Opin. Immunol. 2002, 14, 798–802. [Google Scholar] [CrossRef]
- Jönsson, F.; Mancardi, D.A.; Albanesi, M.; Bruhns, P. Neutrophils in local and systemic antibody-dependent inflammatory and anaphylactic reactions. J. Leukoc. Biol. 2013, 94, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Gonzalez, J.; Edwards, K.; Mallajosyula, V.; Buzzanco, A.S.; Sherwood, R.; Buffone, C.; Kathale, N.; Providenza, S.; Xie, M.M.; et al. Proinflammatory IgG Fc structures in patients with severe COVID-19. Nat. Immunol. 2021, 22, 67–73. [Google Scholar] [CrossRef]
- Argov, S.; Jaffe, C.L.; Krupp, M.; Slor, H.; Shoenfeld, Y. Autoantibody production by patients infected with Leishmania. Clin. Exp. Immunol. 1989, 76, 190–197. [Google Scholar]
- Montes, C.L.; Acosta-Rodríguez, E.V.; Merino, M.C.; Bermejo, D.A.; Gruppi, A. Polyclonal B cell activation in infections: Infectious agents’ devilry or defense mechanism of the host? J. Leukoc. Biol. 2007, 82, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Liberopoulos, E.; Kei, A.; Apostolou, F.; Elisaf, M. Autoimmune manifestations in patients with visceral leishmaniasis. J. Microbiol. Immunol. Infect. 2013, 46, 302–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.V.; Barreira, M.C.R.; Sartori, A.; Campos-Neto, A.; Rossi, M.A. Mesangial proliferative glomerulonephritis associated with progressive amyloid deposition in hamsters experimentally infected with Leishmania donovani. Am. J. Pathol. 1985, 120, 256–262. [Google Scholar]
- Riça-Capela, M.; Cortes, S.; Leandro, C.; Peleteiro, M.; Santos-Gomes, G.; Campino, L. Immunological and histopathological studies in a rodent model infected with Leishmania infantum promastigotes or amastigotes. Parasitol. Res. 2003, 89, 163–169. [Google Scholar] [CrossRef]
- De Vallière, S.; Bullani, R.; Buffet, P.A.; Mary, C.; Tarr, P.E.; Greub, G.; Joneberg, J.E.; Rotman, S.; Gillmore, J.D. AA-Amyloidosis Caused by Visceral Leishmaniasis in a Human Immunodeficiency Virus-Infected Patient. Am. J. Trop. Med. Hyg. 2009, 81, 209–212. [Google Scholar] [CrossRef]
- Mizutani, Y.; Tsuge, S.; Shiogama, K.; Shimomura, R.; Kamoshida, S.; Inada, K.-I.; Tsutsumi, Y. Enzyme-labeled Antigen Method. J. Histochem. Cytochem. 2008, 57, 101–111. [Google Scholar] [CrossRef]
- Oracki, S.A.; Walker, J.A.; Hibbs, M.L.; Corcoran, L.M.; Tarlinton, D.M. Plasma cell development and survival. Immunol. Rev. 2010, 237, 140–159. [Google Scholar] [CrossRef] [PubMed]
- Eikelenboom, P.; Boorsma, D.M.; van Rooijen, N. The development of IgM- and IgG-containing plasmablasts in the white pulp of the spleen after stimulation with a thymus-independent antigen (LPS) and a thymus-dependent antigen (SRBC). Z. Zellforsch. Mikrosk. Anat. 1982, 226, 83–95. [Google Scholar] [CrossRef]
- Silva, J.S.; Andrade, A.C.; Santana, C.C.; Santos, L.Q.; De Oliveira, C.I.; Veras, P.S.T.; Vassallo, J.; Dos-Santos, W.L.C. Low CXCL13 Expression, Splenic Lymphoid Tissue Atrophy and Germinal Center Disruption in Severe Canine Visceral Leishmaniasis. PLoS ONE 2012, 7, e29103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, Y.; Sanjoba, C.; Omachi, S.; Matsumoto, Y. Elevation of Serum B-Cell Activating Factor Levels During Visceral Leishmaniasis. Am. J. Trop. Med. Hyg. 2014, 91, 912–914. [Google Scholar] [CrossRef]
- Silva-Barrios, S.; Charpentier, T.; Stäger, S. The Deadly Dance of B Cells with Trypanosomatids. Trends Parasitol. 2018, 34, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Krawczak, F.D.S.; Reis, I.A.; Silveira, J.; Avelar, D.M.; Marcelino, A.P.; Werneck, G.L.; Labruna, M.B.; Paz, G.F. Leishmania, Babesia and Ehrlichia in urban pet dogs: Co-infection or cross-reaction in serological methods? Rev. Soc. Bras. Med. Trop. 2015, 48, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Baxarias, M.; Álvarez-Fernández, A.; Martinez-Orellana, P.; Montserrat-Sangrà, S.; Ordeix, L.; Rojas, A.; Nachum-Biala, Y.; Baneth, G.; Solano-Gallego, L. Does co-infection with vector-borne pathogens play a role in clinical canine leishmaniosis? Parasites Vectors 2018, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Solcà, M.D.S.; Andrade, B.; Abbehusen, M.M.C.; Teixeira, C.R.; Khouri, R.; Valenzuela, J.G.; Kamhawi, S.; Bozza, P.; Fraga, D.B.M.; Borges, V.; et al. Circulating Biomarkers of Immune Activation, Oxidative Stress and Inflammation Characterize Severe Canine Visceral Leishmaniasis. Sci. Rep. 2016, 6, 32619. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Tsuge, S.; Takeda, H.; Hasegawa, Y.; Shiogama, K.; Onouchi, T.; Inada, K.; Sawasaki, T.; Tsutsumi, Y. In situ visualization of plasma cells producing antibodies reactive to P orphyromonas gingivalis in periodontitis: The application of the enzyme-labeled antigen method. Mol. Oral Microbiol. 2014, 29, 156–173. [Google Scholar] [CrossRef] [PubMed]
- Onouchi, T.; Mizutani, Y.; Shiogama, K.; Inada, K.; Okada, T.; Naito, K.; Tsutsumi, Y. Application of an enzyme-labeled antigen method for visualizing plasma cells producing antibodies against Strep A, a carbohydrate antigen of Streptococcus pyogenes, in recurrent tonsillitis. Microbiol. Immunol. 2015, 59, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Santana, C.C.; Vassallo, J.; DE Freitas, L.A.R.; Oliveira, G.G.S.; Pontes-De-Carvalho, L.C.; Dos-Santos, W.L.C. Inflammation and structural changes of splenic lymphoid tissue in visceral leishmaniasis: A study on naturally infected dogs. Parasite Immunol. 2008, 30, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | With CanL N(%) | Without CanL N(%) |
|---|---|---|
| N (%) | 8 (100%) | 3 (100%) |
| Sex: | ||
| Male | 7 (88%) | 0 (0%) |
| Female | 1 (12%) | 3 (100%) |
| Estimated age (year): | ||
| <1 | 1 (12%) | 0 (0%) |
| 1 to 2 | 2 (25%) | 0 (0%) |
| >3 | 5 (62%) | 3 (100%) |
| Clinical score: | ||
| Subclinic (≤3) | 0 (0%) | 3 (100%) |
| Mild (≥4 and <7) | 2 (25%) | 0 (0%) |
| Severe (≥7) | 6 (75%) | 0 (0%) |
| Clinical signs of disease: | ||
| Conjunctivitis | 7 (88%) | 0 (0%) |
| Pinna crust | 6 (75%) | 0 (0%) |
| Emaciation | 5 (63%) | 0 (0%) |
| Mucous hipocorated | 5 (63%) | 0 (0%) |
| Onychogryphosis | 5 (63%) | 0 (0%) |
| Splenomegaly | 4 (50%) | 0 (0%) |
| Periocular dermatitis | 4 (50%) | 0 (0%) |
| Alopecia | 4 (50%) | 0 (0%) |
| Seborrheic dermatitis | 4 (50%) | 0 (0%) |
| Lymphadenomegaly | 3 (38%) | 0 (0%) |
| Muzzle depigmentation | 3 (38%) | 0 (0%) |
| Ear ulcer | 2 (25%) | 0 (0%) |
| Hyperkeratosis | 2 (25%) | 0 (0%) |
| Laboratory signs of infection/disease: | ||
| Serum proteins 1: | ||
| Total | 7.6 [6.8–8.0] | 5.8 [5.8–6.8] |
| Albumin | 2.0 [1.7–2.5] | 3.4 [3.4–3.5] |
| A/G ratio | 0.4 [0.2–0.5] | 1.4 [0.8–1.4] |
| Positive qPCR | 6 (75%) | 0 (0%) |
| Positive ELISA | 8 (100%) | - |
| Positive Culture of spleen aspirate | 8 (100%) | - |
| Histological Parameters | With CanL N (%) | Without CanL N (%) |
|---|---|---|
| Perisplenitis | 2 (25%) | 0 (0%) |
| Granuloma | 1 (12.5%) | 0 (0%) |
| Amastigotes | 4 (50%) | 0 (0%) |
| White pulp classification | ||
| Type 1 (organized) | 5 (62.5%) | 3 (100%) |
| Type 3 (disorganized) | 3 (37.5%) | - |
| Lymphoid follicle size | ||
| Normal | 5 (62.5%) | 3 (100%) |
| Athrophied | 2 (25%) | - |
| Hiperplasic | 1 (12.5%) | - |
| Plasmacytosis (cell/mm2) | ||
| Red pulp | 639 ± 346 a | 44.7 ± 5.7 a |
| PALS | 467 ± 228 | 0 |
| Spleen Type 1 | Red Pulp | PALS | ||||
|---|---|---|---|---|---|---|
| Total PC | Anti-Leish-PC | (%) | Total PC | Anti-Leish-PC | (%) | |
| With CanL | ||||||
| 1 | 362 | 45 | (12%) | 815 | 663 | (81%) |
| 1 | 523 | 80 | (15%) | 375 | 81 | (22%) |
| 1 | 569 | 118 | (21%) | 330 | 252 | (76%) |
| 1 | 1212 | 267 | (22%) | 782 | 608 | (78%) |
| 1 | 357 | 155 | (44%) | 164 | 40 | (24%) |
| 3 | 1006 | 377 | (37%) | 424 | 302 | (71%) |
| 3 | 842 | 86 | (10%) | 524 | 419 | (80%) |
| 3 | 239 | 213 | (89%) | 323 | 139 | (41%) |
| Mean ± sd | 639 ± 346 | 168 ± 112 | (23 ± 13%) | 467 ± 228 | 313 ± 234 | (59 ± 26%) |
| Without CanL | ||||||
| 1 | 41 | 2 | (5%) | 0 | 0 | (0%) |
| 1 | 52 | 0 | (0%) | 0 | 0 | (0%) |
| 1 | 47 | 2 | (4.2%) | 0 | 0 | (0%) |
| Mean ± sd | 47± 5.5 | 1.3 ± 1.1 | (3 ± 2.6%) | 0 | 0 | (0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontes, J.L.M.; Mesquita, B.R.; Brito, R.; Gomes, J.C.S.; de Melo, C.V.B.; dos Santos, W.L.C. Anti-Leishmania infantum Antibody-Producing Plasma Cells in the Spleen in Canine Visceral Leishmaniasis. Pathogens 2021, 10, 1635. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121635
Fontes JLM, Mesquita BR, Brito R, Gomes JCS, de Melo CVB, dos Santos WLC. Anti-Leishmania infantum Antibody-Producing Plasma Cells in the Spleen in Canine Visceral Leishmaniasis. Pathogens. 2021; 10(12):1635. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121635
Chicago/Turabian StyleFontes, Jonathan L. M., Bianca R. Mesquita, Reginaldo Brito, Juliana C. S. Gomes, Caroline V. B. de Melo, and Washington L. C. dos Santos. 2021. "Anti-Leishmania infantum Antibody-Producing Plasma Cells in the Spleen in Canine Visceral Leishmaniasis" Pathogens 10, no. 12: 1635. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121635