
The O-Ag Antibody Response to Francisella Is Distinct in Rodents and Higher Animals and Can Serve as a Correlate of Protection

 , , and
, , and
Abstract
:1. Introduction
2. Results
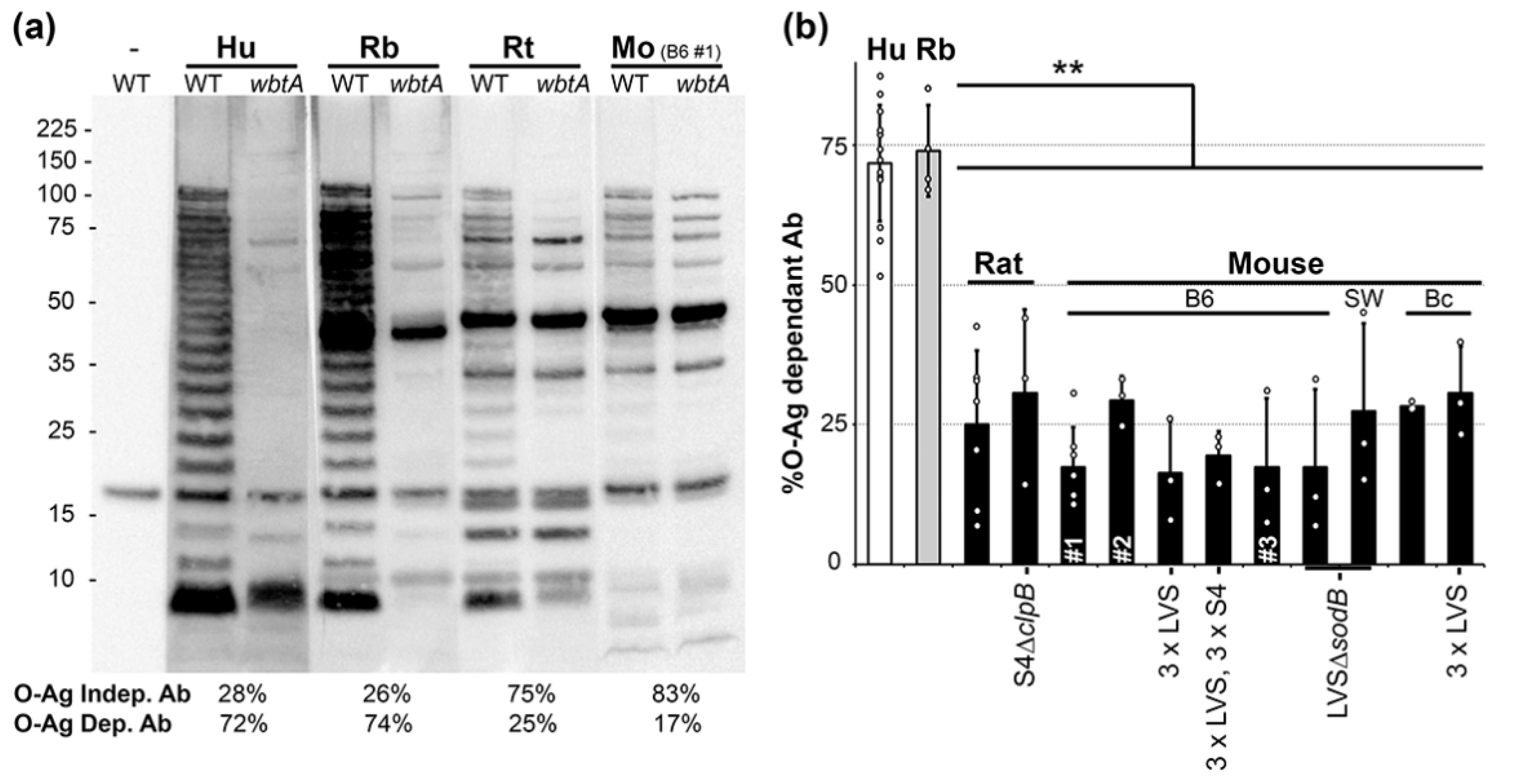
2.1. O-Ag Dependent Ab Reactivity in Vaccinated Humans, Rabbits, Rats and Mice
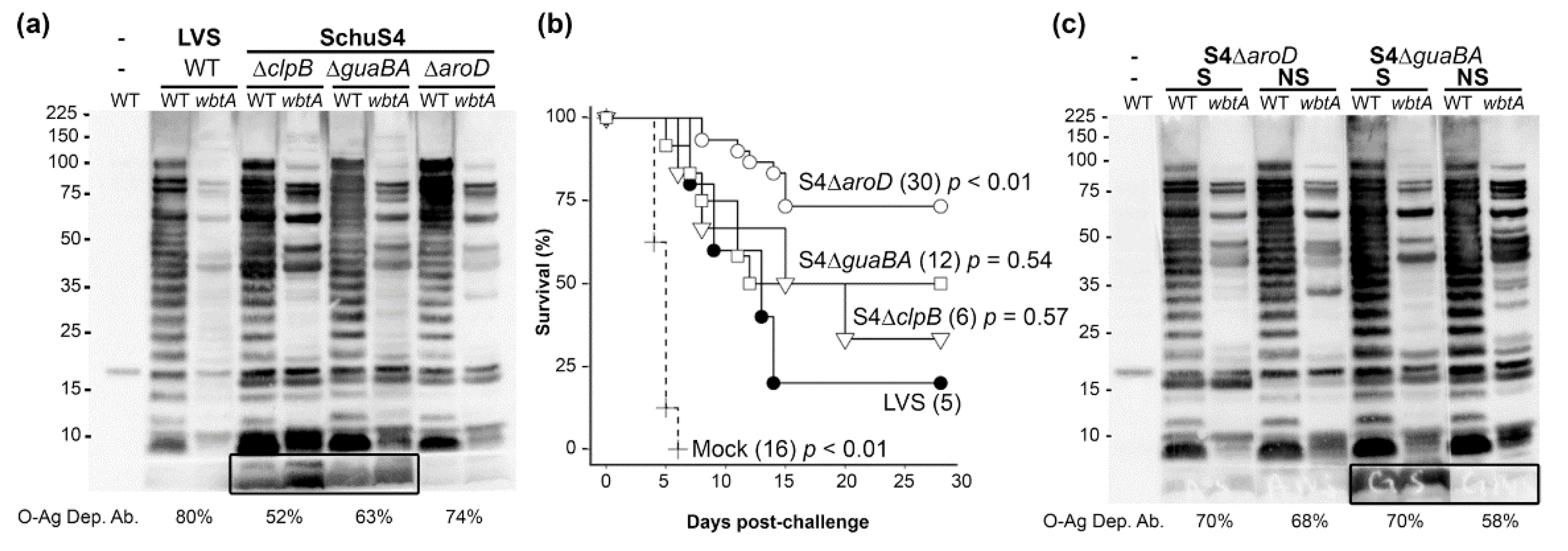
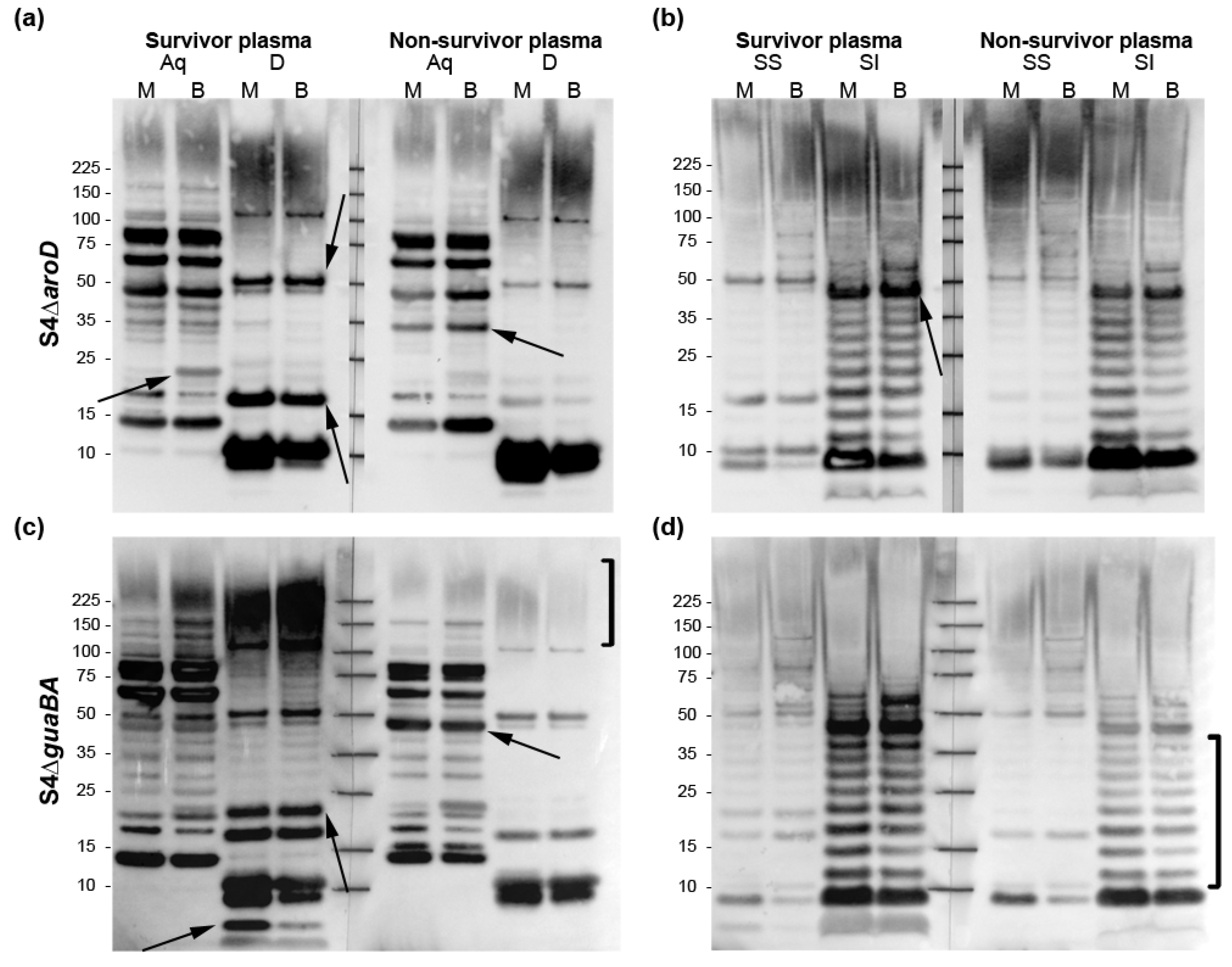
2.2. O-Ag Dep Ab Reactivity in Rabbits Vaccinated with S4ΔclpB, S4ΔguaBA, or S4ΔaroD
2.3. O-Ag Dep Ab Is a Correlate of Protection in S4ΔguaBA-Vaccinated Rabbits
2.4. O-Ag Dep Ab in LVS-Vaccinated Humans
3. Discussion
4. Materials and Methods
4.1. Bacteria
4.2. SDS-PAGE and Western Blot Analysis
4.3. Densitometric Determination of %O-Ag Dep Ab
4.4. Sera/Plasma
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dennis, D.T.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Friedlander, A.M.; Hauer, J.; Layton, M.; et al. Tularemia as a biological weapon: Medical and public health management. JAMA 2001, 285, 2763–2773. [Google Scholar] [CrossRef]
- McCrumb, F.R. Aerosal infection of man with Pasteurella tularensis. Bacteriol. Rev. 1961, 25, 262–267. [Google Scholar] [CrossRef]
- Meibom, K.L.; Charbit, A. The unraveling panoply of Francisella tularensis virulence attributes. Curr. Opin. Microbiol. 2010, 13, 11–17. [Google Scholar] [CrossRef]
- Oyston, P.C.; Griffiths, R. Francisella virulence: Significant advances, ongoing challenges and unmet needs. Expert. Rev. Vaccines 2009, 8, 1575–1585. [Google Scholar] [CrossRef]
- Saslaw, S.; Eigelsbach, H.T.; Prior, J.A.; Wilson, H.E.; Carhart, S. Tularemia vaccine study. II. Respiratory challenge. Arch. Intern. Med. 1961, 107, 702–714. [Google Scholar] [CrossRef]
- Hornick, R.B.; Eigelsbach, H.T. Aerogenic immunization of man with live Tularemia vaccine. Bacteriol. Rev. 1966, 30, 532–538. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Pasetti, M.F.; Barry, E.M.; Nataro, J.P.; Wasserman, S.S.; Sztein, M.B.; Picking, W.D.; Levine, M.M. Deletion in the Shigella enterotoxin genes further attenuates Shigella flexneri 2a bearing guanine auxotrophy in a phase 1 trial of CVD 1204 and CVD 1208. J. Infect. Dis. 2004, 190, 1745–1754. [Google Scholar] [CrossRef] [Green Version]
- Kotloff, K.L.; Simon, J.K.; Pasetti, M.F.; Sztein, M.B.; Wooden, S.L.; Livio, S.; Nataro, J.P.; Blackwelder, W.C.; Barry, E.M.; Picking, W.; et al. Safety and immunogenicity of CVD 1208S, a live, oral DeltaguaBA Deltasen Deltaset Shigella flexneri 2a vaccine grown on animal-free media. Hum. Vaccines 2007, 3, 268–275. [Google Scholar] [CrossRef]
- Tacket, C.O.; Sztein, M.B.; Losonsky, G.A.; Wasserman, S.S.; Nataro, J.P.; Edelman, R.; Pickard, D.; Dougan, G.; Chatfield, S.N.; Levine, M.M. Safety of live oral Salmonella typhi vaccine strains with deletions in htrA and aroC aroD and immune response in humans. Infect. Immun. 1997, 65, 452–456. [Google Scholar] [CrossRef] [Green Version]
- Conlan, J.W.; Sjostedt, A.; Gelhaus, H.C.; Fleming, P.; McRae, K.; Cobb, R.R.; De Pascalis, R.; Elkins, K.L. Modern Development and Production of a New Live Attenuated Bacterial Vaccine, SCHU S4 DeltaclpB, to Prevent Tularemia. Pathogens 2021, 10, 795. [Google Scholar] [CrossRef]
- Cunningham, A.L.; Mann, B.J.; Qin, A.; Santiago, A.E.; Grassel, C.; Lipsky, M.; Vogel, S.N.; Barry, E.M. Characterization of Schu S4 aro mutants as live attenuated tularemia vaccine candidates. Virulence 2020, 11, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Santiago, A.E.; Cole, L.E.; Franco, A.; Vogel, S.N.; Levine, M.M.; Barry, E.M. Characterization of rationally attenuated Francisella tularensis vaccine strains that harbor deletions in the guaA and guaB genes. Vaccine 2009, 27, 2426–2436. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, K.J.; Bowling, J.D.; Barry, E.M.; Hazlett, K.R.O.; Reed, D.S. Development, Characterization, and Standardization of a Nose-Only Inhalation Exposure System for Exposure of Rabbits to Small-Particle Aerosols Containing Francisella tularensis. Infect. Immun. 2019, 87, e00198-19. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, K.J.; Bowling, J.D.; Stinson, E.; Cole, K.S.; Mann, B.J.; Namjoshi, P.; Hazlett, K.R.O.; Barry, E.M.; Reed, D.S. Aerosol prime-boost vaccination provides strong protection in outbred rabbits against virulent type A Francisella tularensis. PLoS ONE 2018, 13, e0205928. [Google Scholar] [CrossRef]
- Chu, P.; Cunningham, A.L.; Yu, J.-J.; Nguyen, J.Q.; Barker, J.R.; Lyons, C.R.; Wilder, J.; Valderas, M.; Sherwood, R.L.; Arulanandam, B.P.; et al. Live attenuated Francisella novicida vaccine protects against Francisella tularensis pulmonary challenge in rats and non-human primates. PLoS Pathog. 2014, 10, e1004439. [Google Scholar] [CrossRef] [Green Version]
- Jemski, J.V. Respiratory tularemia: Comparison of selected routes of vaccination in Fischer 344 rats. Infect. Immun. 1981, 34, 766–772. [Google Scholar] [CrossRef] [Green Version]
- Kostiala, A.A.; McGregor, D.D.; Logie, P.S. Tularaemia in the rat. I. The cellular basis on host resistance to infection. Immunology 1975, 28, 855–869. [Google Scholar]
- Ray, H.J.; Chu, P.; Wu, T.H.; Lyons, C.R.; Murthy, A.K.; Guentzel, M.N.; Klose, K.E.; Arulanandam, B.P. The Fischer 344 rat reflects human susceptibility to francisella pulmonary challenge and provides a new platform for virulence and protection studies. PLoS ONE 2010, 5, e9952. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.H.; Zsemlye, J.L.; Statom, G.L.; Hutt, J.A.; Schrader, R.M.; Scrymgeour, A.A.; Lyons, C.R. Vaccination of Fischer 344 rats against pulmonary infections by Francisella tularensis type A strains. Vaccine 2009, 27, 4684–4693. [Google Scholar] [CrossRef] [Green Version]
- Baskerville, A.; Hambleton, P. Pathogenesis and pathology of respiratory tularaemia in the rabbit. Br. J. Exp. Pathol. 1976, 57, 339–347. [Google Scholar]
- Pasetti, M.F.; Cuberos, L.; Horn, T.L.; Shearer, J.D.; Matthews, S.J.; House, R.V.; Sztein, M.B. An improved Francisella tularensis live vaccine strain (LVS) is well tolerated and highly immunogenic when administered to rabbits in escalating doses using various immunization routes. Vaccine 2008, 26, 1773–1785. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.S.; Smith, L.; Dunsmore, T.; Trichel, A.; Ortiz, L.A.; Cole, K.S.; Barry, E. Pneumonic tularemia in rabbits resembles the human disease as illustrated by radiographic and hematological changes after infection. PLoS ONE 2011, 6, e24654. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.S.; Smith, L.k.P.; Cole, K.S.; Santiago, A.E.; Mann, B.J.; Barry, E.M. Live attenuated mutants of Francisella tularensis protect rabbits against aerosol challenge with a virulent type A strain. Infect. Immun. 2014, 82, 2098–2105. [Google Scholar] [CrossRef] [Green Version]
- Day, W.C.; Berendt, R.F. Experimental tularemia in Macaca mulatta: Relationship of aerosol particle size to the infectivity of airborne Pasteurella tularensis. Infect. Immun. 1972, 5, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Eigelsbach, H.T.; Tulis, J.J.; McGavran, M.H.; White, J.D. Live tularemia vaccine i.: Host-Parasite Relationship in Monkeys Vaccinated Intracutaneously or Aerogenically. J. Bacteriol. 1962, 84, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Schricker, R.L.; Eigelsbach, H.T.; Mitten, J.Q.; Hall, W.C. Pathogenesis of tularemia in monkeys aerogenically exposed to Francisella tularensis 425. Infect. Immun. 1972, 5, 734–744. [Google Scholar] [CrossRef] [Green Version]
- Tulis, J.J.; Eigelsbach, H.T.; Kerpsack, R.W. Host-parasite relationship in monkeys administered live tularemia vaccine. Am. J. Pathol. 1970, 58, 329–336. [Google Scholar]
- Twenhafel, N.A.; Alves, D.A.; Purcell, B.K. Pathology of inhalational Francisella tularensis spp. tularensis SCHU S4 infection in African green monkeys (Chlorocebus aethiops). Vet. Pathol. 2009, 46, 698–706. [Google Scholar] [CrossRef]
- Holland, K.M.; Rosa, S.J.; Hazlett, K.R.O. Francisella tularensis—Immune Cell Activator, Suppressor, or Stealthy Evader: The Evolving View from the Petri Dish. J. Bioterror. Biodef. 2016, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Huntley, J.F.; Conley, P.G.; Hagman, K.E.; Norgard, M.V. Characterization of Francisella tularensis outer membrane proteins. J. Bacteriol. 2007, 189, 561–574. [Google Scholar] [CrossRef] [Green Version]
- Hazlett, K.R.O.; Caldon, S.D.; McArthur, D.G.; Cirillo, K.A.; Kirimanjeswara, G.S.; Magguilli, M.L.; Malik, M.; Shah, A.; Broderick, S.; Golovliov, I.; et al. Adaptation of Francisella tularensis to the mammalian environment is governed by cues which can be mimicked in vitro. Infect. Immun. 2008, 76, 4479–4488. [Google Scholar] [CrossRef] [Green Version]
- Zarrella, T.M.; Singh, A.; Bitsaktsis, C.; Rahman, T.; Sahay, B.; Feustel, P.J.; Gosselin, E.J.; Sellati, T.J.; Hazlett, K.R.O. Host-Adaptation of Francisella tularensis Alters the Bacterium’s Surface-Carbohydrates to Hinder Effectors of Innate and Adaptive Immunity. PLoS ONE 2011, 6, e22335. [Google Scholar] [CrossRef] [Green Version]
- Twine, S.; Shen, H.; Harris, G.; Chen, W.; Sjostedt, A.; Ryden, P.; Conlan, W. BALB/c mice, but not C57BL/6 mice immunized with a DeltaclpB mutant of Francisella tularensis subspecies tularensis are protected against respiratory challenge with wild-type bacteria: Association of protection with post-vaccination and post-challenge immune responses. Vaccine 2012, 30, 3634–3645. [Google Scholar] [CrossRef]
- Sunagar, R.; Kumar, S.; Namjoshi, P.; Rosa, S.J.; Hazlett, K.R.O.; Gosselin, E.J. Evaluation of an outbred mouse model for Francisella tularensis vaccine development and testing. PLoS ONE 2018, 13, e0207587. [Google Scholar] [CrossRef]
- Shen, H.; Harris, G.; Chen, W.; Sjostedt, A.; Ryden, P.; Conlan, W. Molecular immune responses to aerosol challenge with Francisella tularensis in mice inoculated with live vaccine candidates of varying efficacy. PLoS ONE 2010, 5, e13349. [Google Scholar] [CrossRef]
- Ryden, P.; Twine, S.; Shen, H.; Harris, G.; Chen, W.; Sjostedt, A.; Conlan, W. Correlates of protection following vaccination of mice with gene deletion mutants of Francisella tularensis subspecies tularensis strain, SCHU S4 that elicit varying degrees of immunity to systemic and respiratory challenge with wild-type bacteria. Mol. Immunol. 2013, 54, 58–67. [Google Scholar] [CrossRef]
- Griffin, A.J.; Crane, D.D.; Wehrly, T.D.; Bosio, C.M. Successful protection against tularemia in C57BL/6 mice is correlated with expansion of Francisella tularensis-specific effector T cells. Clin. Vaccine Immunol. 2015, 22, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Golovliov, I.; Lindgren, H.; Eneslatt, K.; Conlan, W.; Mosnier, A.; Henry, T.; Sjostedt, A. An In Vitro Co-culture Mouse Model Demonstrates Efficient Vaccine-Mediated Control of Francisella tularensis SCHU S4 and Identifies Nitric Oxide as a Predictor of Efficacy. Front. Cell. Infect. Microbiol. 2016, 6, 152. [Google Scholar] [CrossRef] [Green Version]
- De Pascalis, R.; Mittereder, L.; Kennett, N.J.; Elkins, K.L. Activities of Murine Peripheral Blood Lymphocytes Provide Immune Correlates That Predict Francisella tularensis Vaccine Efficacy. Infect. Immun. 2016, 84, 1054–1061. [Google Scholar] [CrossRef] [Green Version]
- De Pascalis, R.; Mittereder, L.; Chou, A.Y.; Kennett, N.J.; Elkins, K.L. Francisella tularensis Vaccines Elicit Concurrent Protective T- and B-Cell Immune Responses in BALB/cByJ Mice. PLoS ONE 2015, 10, e0126570. [Google Scholar] [CrossRef] [Green Version]
- De Pascalis, R.; Hahn, A.; Brook, H.M.; Ryden, P.; Donart, N.; Mittereder, L.; Frey, B.; Wu, T.H.; Elkins, K.L. A panel of correlates predicts vaccine-induced protection of rats against respiratory challenge with virulent Francisella tularensis. PLoS ONE 2018, 13, e0198140. [Google Scholar]
- De Pascalis, R.; Chou, A.Y.; Bosio, C.M.; Huang, C.Y.; Follmann, D.A.; Elkins, K.L. Development of functional and molecular correlates of vaccine-induced protection for a model intracellular pathogen, F. tularensis LVS. PLoS Pathog. 2012, 8, e1002494. [Google Scholar] [CrossRef]
- Twine, S.M.; Petit, M.D.; Fulton, K.M.; House, R.V.; Conlan, J.W. Immunoproteomics analysis of the murine antibody response to vaccination with an improved Francisella tularensis live vaccine strain (LVS). PLoS ONE 2010, 5, e10000. [Google Scholar] [CrossRef]
- Twine, S.M.; Petit, M.D.; Shen, H.; Mykytczuk, N.C.S.; Kelly, J.F.; Conlan, J.W. Immunoproteomic analysis of the murine antibody response to successful and failed immunization with live anti-Francisella vaccines. Biochem. Biophys. Res. Commun. 2006, 346, 999–1008. [Google Scholar] [CrossRef]
- Rahhal, R.M.; Vanden Bush, T.J.; McLendon, M.K.; Apicella, M.A.; Bishop, G.A. Differential effects of Francisella tularensis lipopolysaccharide on B lymphocytes. J. Leukoc. Biol. 2007, 82, 813–820. [Google Scholar] [CrossRef]
- Beura, L.K.; Hamilton, S.E.; Bi, K.; Schenkel, J.M.; Odumade, O.A.; Casey, K.A.; Thompson, E.A.; Fraser, K.A.; Rosato, P.C.; Filali-Mouhim, A.; et al. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 2016, 532, 512–516. [Google Scholar] [CrossRef]
- Sangesland, M.; Yousif, A.S.; Ronsard, L.; Kazer, S.W.; Zhu, A.L.; Gatter, G.J.; Hayward, M.R.; Barnes, R.M.; Quirindongo-Crespo, M.; Rohrer, D.; et al. A Single Human VH-gene Allows for a Broad-Spectrum Antibody Response Targeting Bacterial Lipopolysaccharides in the Blood. Cell Rep. 2020, 32, 108065. [Google Scholar] [CrossRef]
- Plotkin, S.A. Updates on immunologic correlates of vaccine-induced protection. Vaccine 2020, 38, 2250–2257. [Google Scholar] [CrossRef]
- Geier, H.; Celli, J. Phagocytic receptors dictate phagosomal escape and intracellular proliferation of Francisella tularensis. Infect. Immun. 2011, 79, 2204–2214. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Yang, J.; Zhao, D.; Kawula, T.H.; Banas, J.A.; Zhang, J.R. Genome-wide identification of Francisella tularensis virulence determinants. Infect. Immun. 2007, 75, 3089–3101. [Google Scholar] [CrossRef] [Green Version]
- Holland-Tummillo, K.M.; Shoudy, L.E.; Steiner, D.; Kumar, S.; Rosa, S.J.; Namjoshi, P.; Singh, A.; Sellati, T.J.; Gosselin, E.J.; Hazlett, K.R. Autotransporter-Mediated Display of Complement Receptor Ligands by Gram-Negative Bacteria Increases Antibody Responses and Limits Disease Severity. Pathogens 2020, 9, 375. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NS | S | |
|---|---|---|
| Range | 43–70% | 68–81% |
| Mean | 60% | 76% |
| WRS | p = 0.009 | |
| ROC | AUC = 0.94, 95% CI (0.82, 1.00) 70% cut-off = 83% Sensitivity, Specificity. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shoudy, L.E.; Namjoshi, P.; Giordano, G.; Kumar, S.; Bowling, J.D.; Gelhaus, C.; Barry, E.M.; Hazlett, A.J.; Hazlett, B.A.; Cooper, K.L.; et al. The O-Ag Antibody Response to Francisella Is Distinct in Rodents and Higher Animals and Can Serve as a Correlate of Protection. Pathogens 2021, 10, 1646. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121646
Shoudy LE, Namjoshi P, Giordano G, Kumar S, Bowling JD, Gelhaus C, Barry EM, Hazlett AJ, Hazlett BA, Cooper KL, et al. The O-Ag Antibody Response to Francisella Is Distinct in Rodents and Higher Animals and Can Serve as a Correlate of Protection. Pathogens. 2021; 10(12):1646. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121646
Chicago/Turabian StyleShoudy, Lauren E., Prachi Namjoshi, Gabriela Giordano, Sudeep Kumar, Jennifer D. Bowling, Carl Gelhaus, Eileen M. Barry, Allan J. Hazlett, Brian A. Hazlett, Kristine L. Cooper, and et al. 2021. "The O-Ag Antibody Response to Francisella Is Distinct in Rodents and Higher Animals and Can Serve as a Correlate of Protection" Pathogens 10, no. 12: 1646. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121646