
Tryptophanyl tRNA Synthetase from Human Macrophages Infected by Porphyromonas gingivalis Induces a Proinflammatory Response Associated with Atherosclerosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
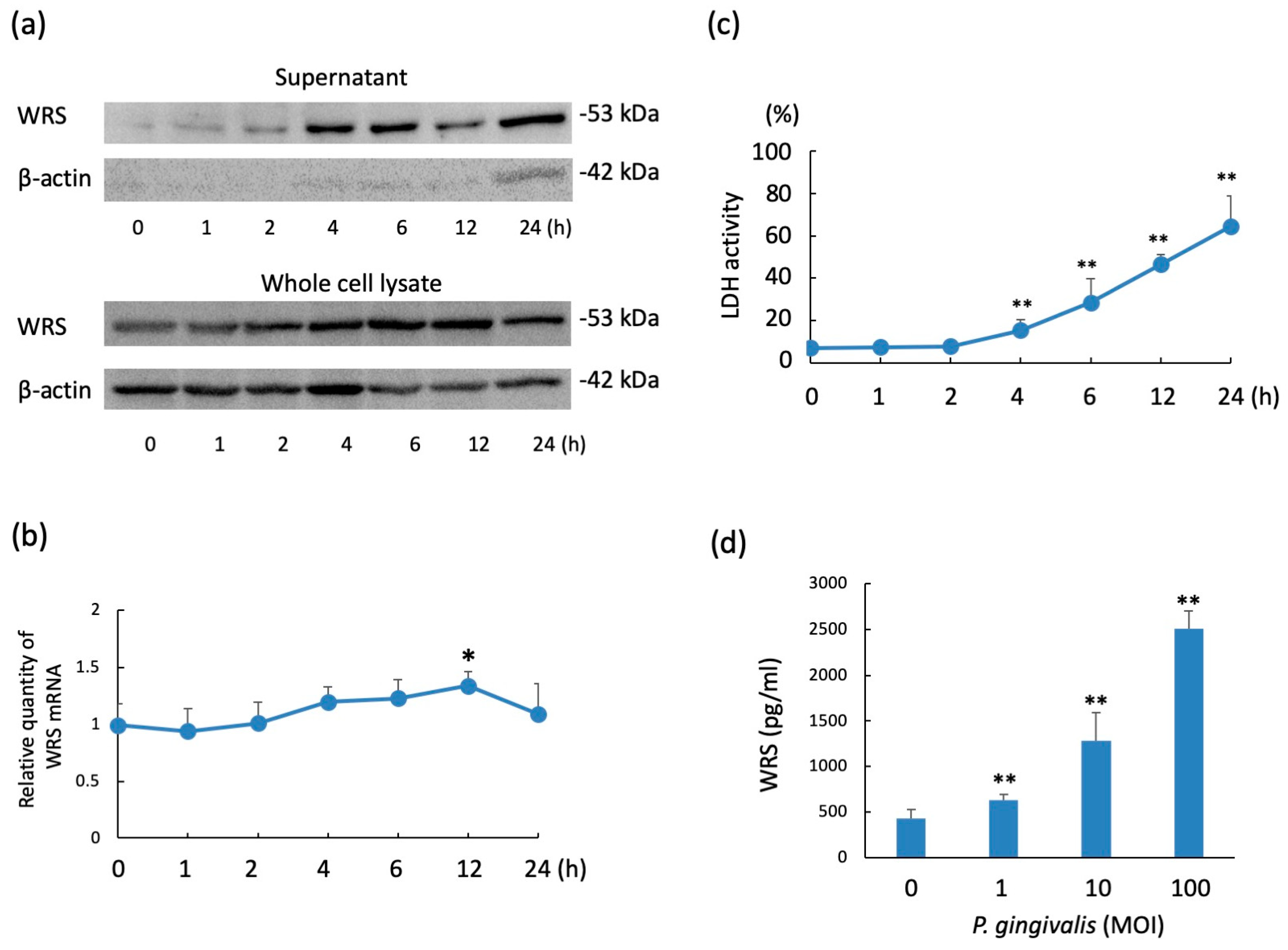
2.1. WRS Levels in THP-1 Cells Stimulated with P. gingivalis
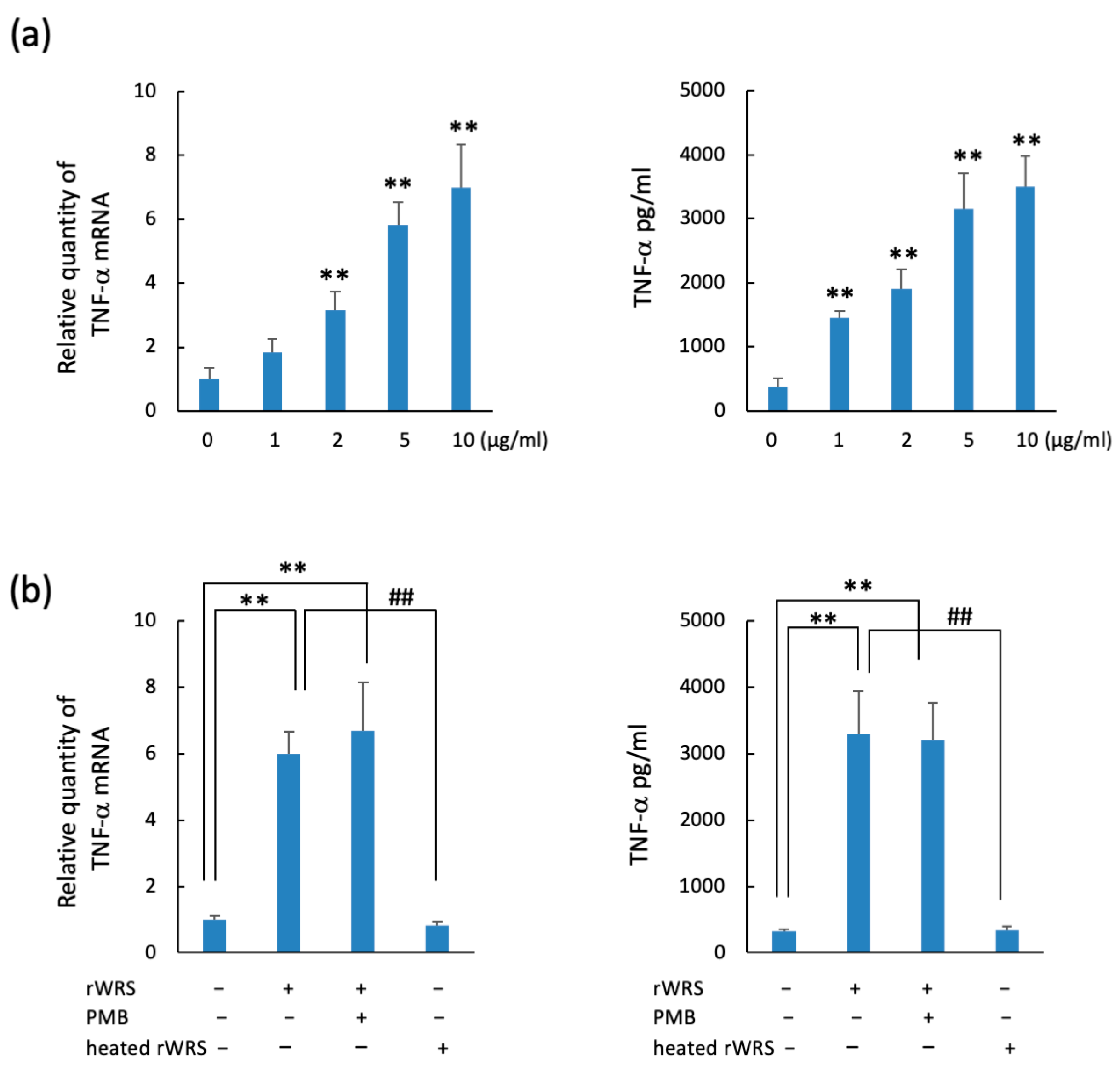
2.2. TNF-α and IL-8 Expressions in THP-1 Cells Treated with Recombinant WRS (rWRS)
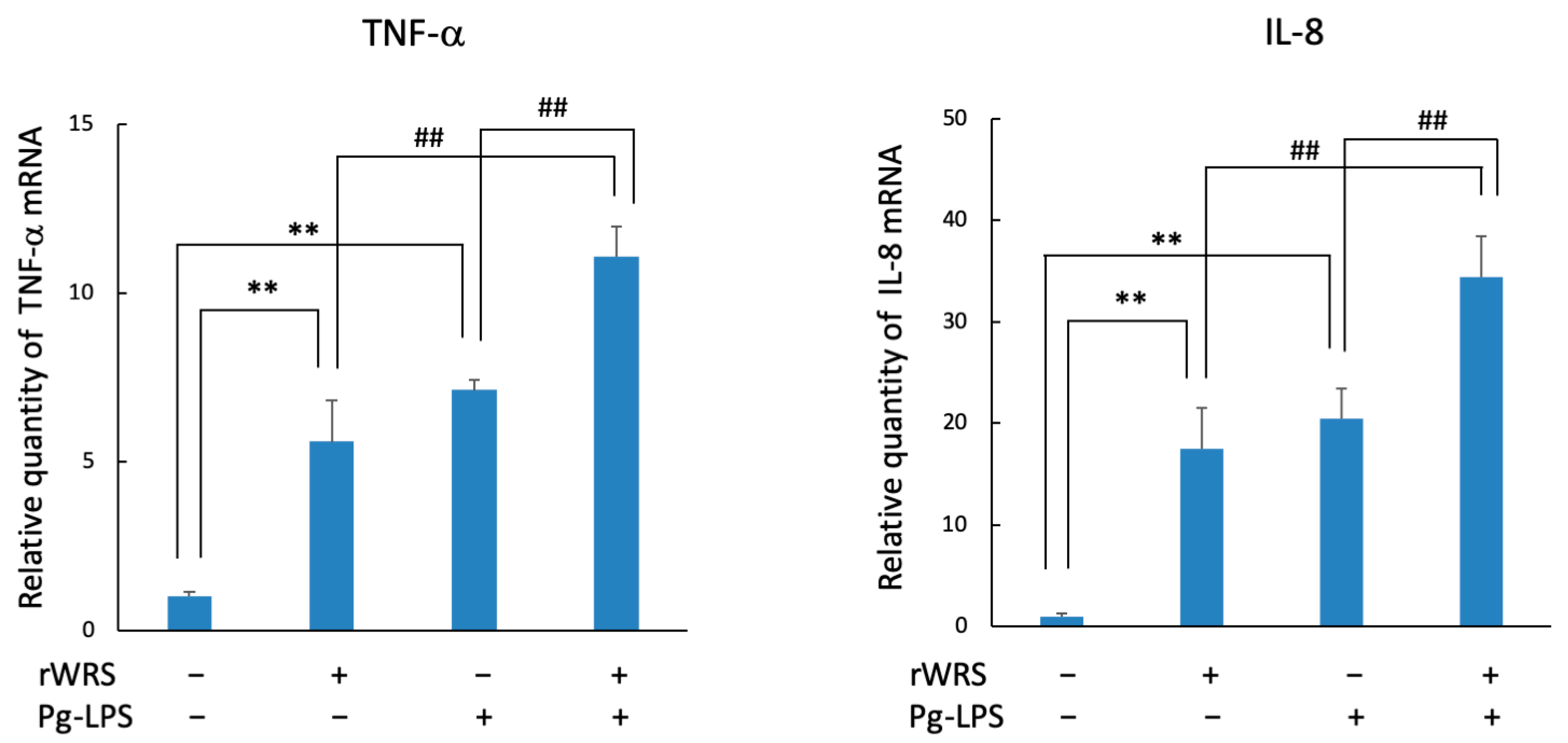
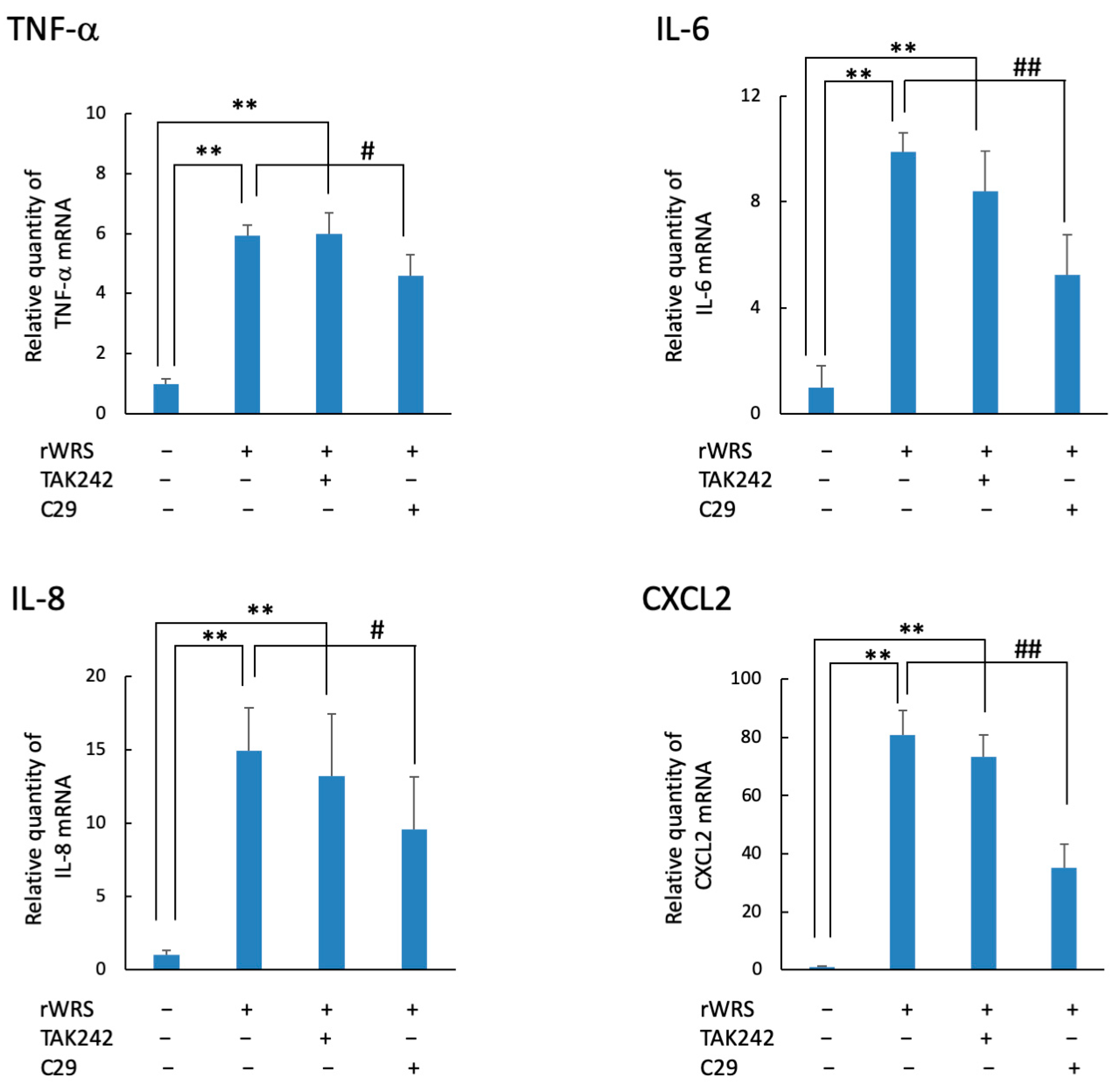
2.3. Cytokine Expression in THP-1 Cells Treated with rWRS and the Effect of the Inhibitor of TLR Signaling
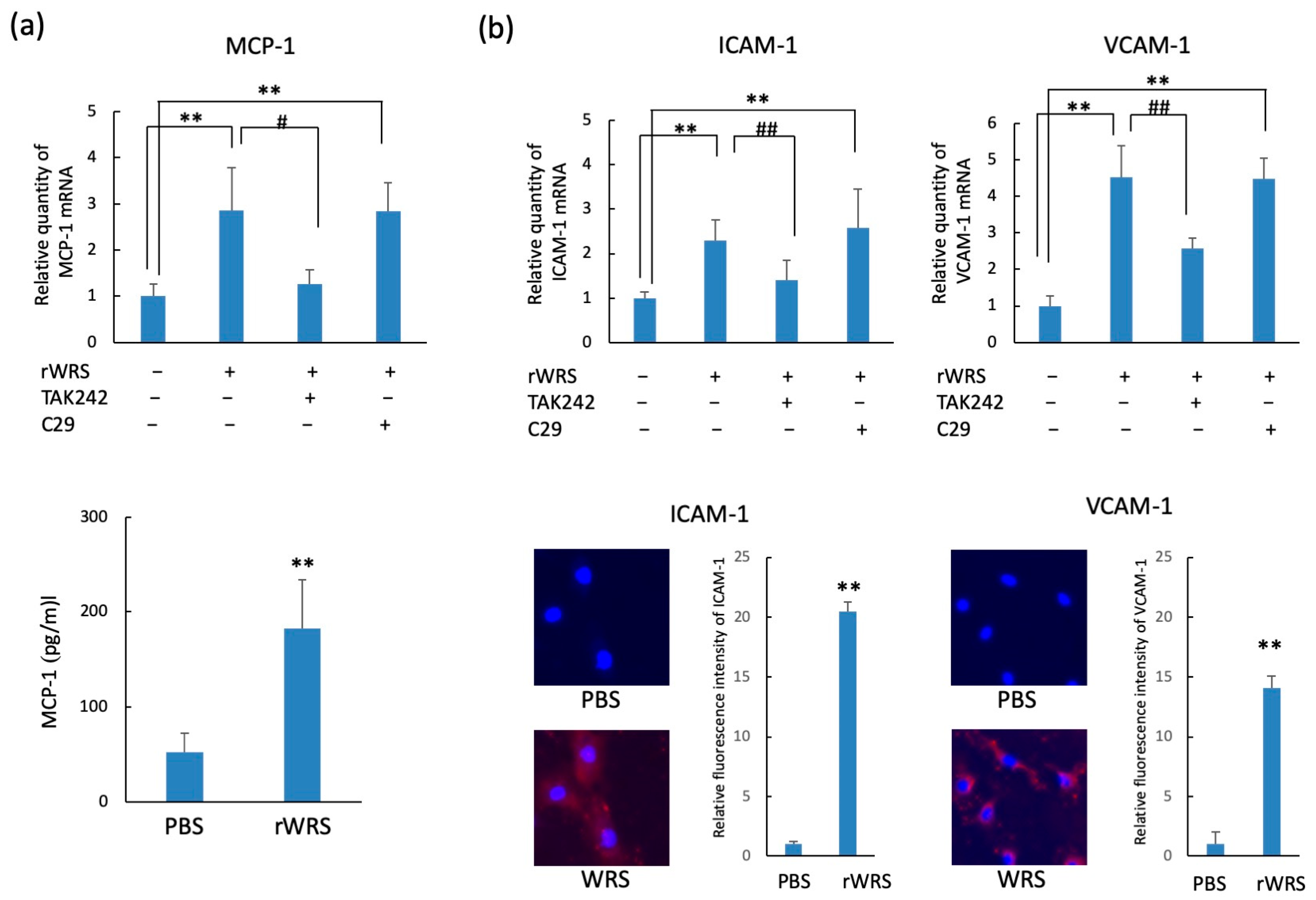
2.4. Chemokine and Adhesion Molecule Expressions in HUVECs Treated with rWRS
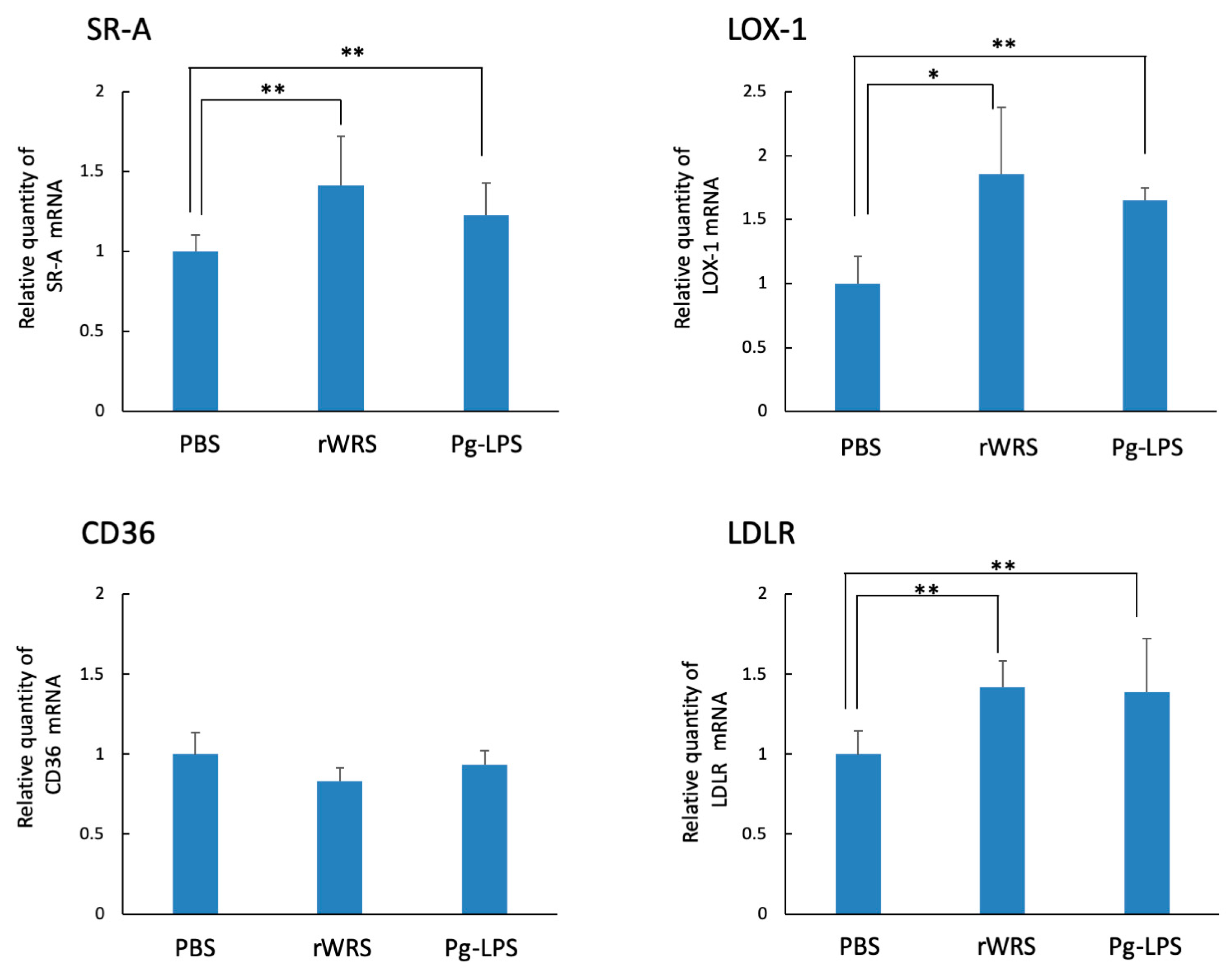
2.5. Effect of rWRS on Native LDLR and oxLDLR Gene Expressions in THP-1 Cells
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Cells and Culture Conditions
4.3. Detection of WRS in Culture Supernatants and Whole Cell Lysate of THP-1 Cells Infected with P. gingivalis
4.4. Construction of an Expression Vector for WRS Protein and Expression and Purification of rWRS
4.5. Cytokine, Chemokine, and Leukocyte Adhesion Molecule Expression
4.6. Immunofluorescence Study
4.7. Expressions of LDLR mRNA
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Bartova, J.; Janatova, T.; Prochazkova, J.; Duskova, J. Porphyromonas gingivalis: Major periodontopathic pathogen overview. J. Immunol. Res. 2014, 2014, 476068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Gaudilliere, D.K.; Culos, A.; Djebali, K.; Tsai, A.S.; Ganio, E.A.; Choi, W.M.; Han, X.; Maghaireh, A.; Choisy, B.; Baca, Q.; et al. Systemic immunologic consequences of chronic periodontitis. J. Dent. Res. 2019, 98, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Genco, R.J. Current view of risk factors for periodontal diseases. J. Periodontol. 1996, 67 (Suppl. 10), 1041–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasanayake, A.P.; Russell, S.; Boyd, D.; Madianos, P.N.; Forster, T.; Hill, E. Preterm low birth weight and periodontal disease among African Americans. Dent. Clin. N. Am. 2003, 47, 115–125. [Google Scholar] [CrossRef]
- Fiorillo, L.; Cervino, G.; Laino, L.; D’Amico, C.; Mauceri, R.; Tozum, T.F.; Gaeta, M.; Cicciù, M. Porphyromonas gingivalis, periodontal and systemic implications: A systematic review. Dent. J. 2019, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, F.; Xie, M.; Huang, X.; Long, Y.; Lu, X.; Wang, X.; Chen, L. Porphyromonas gingivalis and its systemic impact: Current status. Pathogens 2020, 9, 944. [Google Scholar] [CrossRef]
- Bostanci, N.; Belibasakis, G.N. Porphyromonas gingivalis: An invasive and evasive opportunistic oral pathogen. FEMS Microbiol. Lett. 2012, 333, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Zhou, W.; Wang, H.; Liang, S. Roles of Porphyromonas gingivalis and its virulence factors in periodontitis. Adv. Protein Chem. Struct. Biol. 2020, 120, 45–84. [Google Scholar] [CrossRef]
- Nishida, E.; Hara, Y.; Kaneko, T.; Ikeda, Y.; Ukai, T.; Kato, I. Bone resorption and local interleukin-1alpha and interleukin-1beta synthesis induced by Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis lipopolysaccharide. J. Periodont. Res. 2001, 36, 1–8. [Google Scholar] [CrossRef]
- Zhou, Q.; Desta, T.; Fenton, M.; Graves, D.T.; Amar, S. Cytokine profiling of macrophages exposed to Porphyromonas gingivalis, its lipopolysaccharide, or its FimA protein. Infect. Immun. 2005, 73, 935–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.K.; Yeh, K.C.; Wu, M.F.; Li, C.L.; Tseng, C.C. An acute injection of Porphyromonas gingivalis lipopolysaccharide modulates the OPG/RANKL system and interleukin-6 in an ovariectomized mouse model. Oral Microbiol. Immunol. 2008, 23, 220–225. [Google Scholar] [CrossRef]
- Akkaoui, J.; Yamada, C.; Duarte, C.; Ho, A.; Vardar-Sengul, S.; Kawai, T.; Movila, A. Contribution of Porphyromonas gingivalis lipopolysaccharide to experimental periodontitis in relation to aging. GeroScience 2021, 43, 367–376. [Google Scholar] [CrossRef]
- Amano, A.; Nakagawa, I.; Okahashi, N.; Hamada, N. Variations of Porphyromonas gingivalis fimbriae in relation to microbial pathogenesis. J. Periodont. Res. 2004, 39, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Davey, M.; Yumoto, H.; Gibson, F.C., 3rd; Genco, C.A. Fimbria-dependent activation of pro-inflammatory molecules in Porphyromonas gingivalis infected human aortic endothelial cells. Cell. Microbiol. 2006, 8, 738–757. [Google Scholar] [CrossRef] [PubMed]
- Enersen, M.; Nakano, K.; Amano, A. Porphyromonas gingivalis fimbriae. J. Oral Microbiol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Lourbakos, A.; Potempa, J.; Travis, J.; D’Andrea, M.R.; Andrade-Gordon, P.; Santulli, R.; Mackie, E.J.; Pike, R.N. Arginine-specific protease from Porphyromonas gingivalis activates protease-activated receptors on human oral epithelial cells and induces interleukin-6 secretion. Infect. Immun. 2001, 69, 5121–5130. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Nguyen, K.A.; Potempa, J. Dichotomy of gingipains action as virulence factors: From cleaving substrates with the precision of a surgeon’s knife to a meat chopper-like brutal degradation of proteins. Periodontol. 2000 2010, 54, 15–44. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, Y.; Matono, S.; Aida, Y.; Hirofuji, T.; Maeda, K. Gingipains in the culture supernatant of Porphyromonas gingivalis cleave CD4 and CD8 on human T cells. J. Periodont. Res. 2002, 37, 464–468. [Google Scholar] [CrossRef]
- Belibasakis, G.N.; Bostanci, N.; Reddi, D. Regulation of protease-activated receptor-2 expression in gingival fibroblasts and Jurkat T cells by Porphyromonas gingivalis. Cell Biol. Int. 2010, 34, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Lee, E.S.; Uddin, M.B.; Kim, T.H.; Kim, J.H.; Chathuranga, K.; Chathuranga, W.A.G.; Jin, M.; Kim, S.; Kim, C.J.; et al. Released tryptophanyl-tRNA synthetase stimulates innate immune responses against viral infection. J. Virol. 2019, 93, e01291-18. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.T.; Yoon, H.K.; Kim, Y.T.; Choi, Y.H.; Lee, W.K.; Jin, M. Tryptophanyl-tRNA synthetase 1 signals activate TREM-1 via TLR2 and TLR4. Biomolecules 2020, 10, 1283. [Google Scholar] [CrossRef] [PubMed]
- Wakasugi, K.; Schimmel, P. Two distinct cytokines released from a human aminoacyl-tRNA synthetase. Science 1999, 284, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Wakasugi, K.; Schimmel, P. Highly differentiated motifs responsible for two cytokine activities of a split human tRNA synthetase. J. Biol. Chem. 1999, 274, 23155–23159. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.H.; Park, S.; Choi, J.J.; Park, B.K.; Rhee, K.H.; Kang, E.; Ahn, S.; Lee, C.H.; Lee, J.S.; Inn, K.S.; et al. Secreted tryptophanyl-tRNA synthetase as a primary defence system against infection. Nat. Microbiol. 2016, 2, 16191. [Google Scholar] [CrossRef]
- De Winther, M.P.; van Dijk, K.W.; Havekes, L.M.; Hofker, M.H. Macrophage scavenger receptor class A: A multifunctional receptor in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 290–297. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Chen, Y.; Lei, H.; Liu, Q.; Moorhead, J.F.; Varghese, Z.; Ruan, X.Z. Inflammatory stress increases unmodified LDL uptake via LDL receptor: An alternative pathway for macrophage foam-cell formation. Inflamm. Res. 2009, 58, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Zhi, A.; Lai, P.F.H.; Wang, G.; Xia, Y.; Xiong, Z.; Zhang, H.; Che, N.; Ai, L. The oral microbiota—A mechanistic role for systemic diseases. Br. Dent. J. 2018, 224, 447–455. [Google Scholar] [CrossRef]
- Graves, D.T.; Corrêa, J.D.; Silva, T.A. The oral microbiota is modified by systemic diseases. J. Dent. Res. 2019, 98, 148–156. [Google Scholar] [CrossRef]
- Chiu, B. Multiple infections in carotid atherosclerotic plaques. Am. Heart J. 1999, 138, S534–S536. [Google Scholar] [CrossRef]
- Ohki, T.; Itabashi, Y.; Kohno, T.; Yoshizawa, A.; Nishikubo, S.; Watanabe, S.; Yamane, G.; Ishihara, K. Detection of periodontal bacteria in thrombi of patients with acute myocardial infarction by polymerase chain reaction. Am. Heart J. 2012, 163, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I.; Taubman, M.A.; Singhrao, S.K. Porphyromonas gingivalis suppresses adaptive immunity in periodontitis, atherosclerosis, and Alzheimer’s disease. J. Oral Microbiol. 2016, 8, 33029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bale, B.F.; Doneen, A.L.; Vigerust, D.J. High-risk periodontal pathogens contribute to the pathogenesis of atherosclerosis. Postgrad. Med. J. 2017, 93, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Carrizales-Sepúlveda, E.F.; Ordaz-Farías, A.; Vera-Pineda, R.; Flores-Ramírez, R. Periodontal disease, systemic inflammation and the risk of cardiovascular disease. Heart Lung Circ. 2018, 27, 1327–1334. [Google Scholar] [CrossRef]
- Greenberg, Y.; King, M.; Kiosses, W.B.; Ewalt, K.; Yang, X.; Schimmel, P.; Reader, J.S.; Tzima, E. The novel fragment of tyrosyl tRNA synthetase, mini-TyrRS, is secreted to induce an angiogenic response in endothelial cells. FASEB J. 2008, 22, 1597–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobin, P.G.; Solis, N.; Machado, Y.; Bell, P.A.; Rai, S.K.; Kwon, N.H.; Kim, S.; Overall, C.M.; Butler, G.S. Moonlighting matrix metalloproteinase substrates: Enhancement of proinflammatory functions of extracellular tyrosyl-tRNA synthetase upon cleavage. J. Biol. Chem. 2020, 295, 2186–2202. [Google Scholar] [CrossRef] [Green Version]
- Park, S.G.; Kim, H.J.; Min, Y.H.; Choi, E.C.; Shin, Y.K.; Park, B.J.; Lee, S.W.; Kim, S. Human lysyl-tRNA synthetase is secreted to trigger proinflammatory response. Proc. Natl. Acad. Sci. USA 2005, 102, 6356–6361. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.H.; Jung, W.Y.; Lee, H.; Kang, Y.; Jang, Y.J.; Hong, S.W.; Jeong, H.J.; Yoon, S.O. Lysyl-tRNA synthetase (KRS) expression in gastric carcinoma and tumor-associated inflammation. Ann. Surg. Oncol. 2014, 21, 2020–2027. [Google Scholar] [CrossRef]
- Lee, M.S.; Kwon, H.; Nguyen, L.T.; Lee, E.Y.; Lee, C.Y.; Choi, S.H.; Kim, M.H. Shiga toxins trigger the secretion of lysyl-tRNA synthetase to enhance proinflammatory responses. J. Microbiol. Biotechnol. 2016, 26, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Shuang Chen, W.; Wong, M.H.; Schulte, D.J.; Arditi, M.; Michelsen, K.S. Differential expression of toll-like receptor 2 (TLR2) and responses to TLR2 ligands between human and murine vascular endothelial cells. J. Endotoxin Res. 2007, 13, 281–296. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V. Toll-like receptors in sepsis-associated cytokine storm and their endogenous negative regulators as future immunomodulatory targets. Int. Immunopharmacol. 2020, 89, 107087. [Google Scholar] [CrossRef] [PubMed]
- Wada, J.; Makino, H. Innate immunity in diabetes and diabetic nephropathy. Nat. Rev. Nephrol. 2016, 12, 13–26. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Wei, L.L.; Zhao, S.; Sverdlov, D.Y.; Vaid, K.A.; Miyamoto, M.; Kuramitsu, K.; Lai, M.; Popov, Y.V. Hepatocyte mitochondria-derived danger signals directly activate hepatic stellate cells and drive progression of liver fibrosis. Nat. Commun. 2020, 11, 2362. [Google Scholar] [CrossRef] [PubMed]
- Audrito, V.; Messana, V.G.; Deaglio, S. NAMPT and NAPRT: Two metabolic enzymes with key roles in inflammation. Front. Oncol. 2020, 10, 358. [Google Scholar] [CrossRef] [Green Version]
- Cicco, S.; Cicco, G.; Racanelli, V.; Vacca, A. Neutrophil extracellular traps (NETs) and damage-associated molecular patterns (DAMPs): Two potential targets for COVID-19 treatment. Mediat. Inflamm. 2020, 2020, 7527953. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Sukhova, G.; Lee, R.T.; Galis, Z.S. Cytokines regulate vascular functions related to stability of the atherosclerotic plaque. J. Cardiovasc. Pharmacol. 1995, 25 (Suppl. 2), S9–S12. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; Di Raimondo, D.; Pecoraro, R.; Arnao, V.; Pinto, A.; Licata, G. Atherosclerosis as an inflammatory disease. Curr. Pharm. Des. 2012, 18, 4266–4288. [Google Scholar] [CrossRef]
- Vabulas, R.M.; Ahmad-Nejad, P.; da Costa, C.; Miethke, T.; Kirschning, C.J.; Häcker, H.; Wagner, H. Endocytosed HSP60s use toll-like receptor 2 (TLR2) and TLR4 to activate the toll/interleukin-1 receptor signaling pathway in innate immune cells. J. Biol. Chem. 2001, 276, 31332–31339. [Google Scholar] [CrossRef] [Green Version]
- Vielma, S.A.; Krings, G.; Lopes-Virella, M.F. Chlamydophila pneumoniae induces ICAM-1 expression in human aortic endothelial cells via protein kinase C-dependent activation of nuclear factor-kappaB. Circ. Res. 2003, 92, 1130–1137. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.; Alp, N.J. Role of Chlamydia pneumoniae in atherosclerosis. Clin. Sci. 2008, 114, 509–531. [Google Scholar] [CrossRef]
- Maekawa, T.; Takahashi, N.; Tabeta, K.; Aoki, Y.; Miyashita, H.; Miyauchi, S.; Miyazawa, H.; Nakajima, T.; Yamazaki, K. Chronic oral infection with Porphyromonas gingivalis accelerates atheroma formation by shifting the lipid profile. PLoS ONE 2011, 6, e20240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.Y.; Wang, L.; Wang, H.M.; Wang, Y.Q.; Liang, Y.F.; Zhao, T.T.; Wu, Y.Z. TLR2 and TLR4 agonists synergistically upregulate SR-A in RAW264.7 through p38. Mol. Immunol. 2007, 44, 2315–2323. [Google Scholar] [CrossRef] [PubMed]
- Hossain, E.; Ota, A.; Karnan, S.; Takahashi, M.; Mannan, S.B.; Konishi, H.; Hosokawa, Y. Lipopolysaccharide augments the uptake of oxidized LDL by upregulating lectin-like oxidized LDL receptor-1 in macrophages. Mol. Cell. Biochem. 2015, 400, 29–40. [Google Scholar] [CrossRef]
- Yu, X.H.; Fu, Y.C.; Zhang, D.W.; Yin, K.; Tang, C.K. Foam cells in atherosclerosis. Clin. Chim. Acta Int. J. Clin. Chem. 2013, 424, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiser, L.; De Winther, M.P.; Makepeace, K.; Hollinshead, M.; Coull, P.; Plested, J.; Kodama, T.; Moxon, E.R.; Gordon, S. The class A macrophage scavenger receptor is a major pattern recognition receptor for Neisseria meningitidis which is independent of lipopolysaccharide and not required for secretory responses. Infect. Immun. 2002, 70, 5346–5354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lega, J.C.; Fabien, N.; Reynaud, Q.; Durieu, I.; Durupt, S.; Dutertre, M.; Cordier, J.F.; Cottin, V. The clinical phenotype associated with myositis-specific and associated autoantibodies: A meta-analysis revisiting the so-called antisynthetase syndrome. Autoimmun. Rev. 2014, 13, 883–891. [Google Scholar] [CrossRef]
- Sasaki, M.; Kodama, Y.; Shimoyama, Y.; Ishikawa, T.; Tajika, S.; Kimura, S. Abiotrophia defectiva adhere to saliva-coated hydroxyapatite beads via interactions between salivary proline-rich-proteins and bacterial glyceralaldehyde-3-phosphate dehydrogenase. Microbiol. Immunol. 2020, 64, 719–729. [Google Scholar] [CrossRef]
- Sasaki, M.; Shimoyama, Y.; Kodama, Y.; Ishikawa, T. Abiotrophia defectiva DnaK promotes fibronectin-mediated adherence to HUVECs and induces a proinflammatory response. Int. J. Mol. Sci. 2021, 22, 8528. [Google Scholar] [CrossRef]
- Matsunaga, N.; Tsuchimori, N.; Matsumoto, T.; Ii, M. TAK-242 (resatorvid), a small-molecule inhibitor of Toll-like receptor (TLR) 4 signaling, binds selectively to TLR4 and interferes with interactions between TLR4 and its adaptor molecules. Mol. Pharmacol. 2011, 79, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Mistry, P.; Laird, M.H.; Schwarz, R.S.; Greene, S.; Dyson, T.; Snyder, G.A.; Xiao, T.S.; Chauhan, J.; Fletcher, S.; Toshchakov, V.Y.; et al. Inhibition of TLR2 signaling by small molecule inhibitors targeting a pocket within the TLR2 TIR domain. Proc. Natl. Acad. Sci. USA 2015, 112, 5455–5460. [Google Scholar] [CrossRef] [Green Version]
- Kodama, Y.; Shimoyama, Y.; Ishikawa, T.; Kimura, S.; Sasaki, M. Characterization and pathogenicity of fibronectin binding protein FbpI of Streptococcus intermedius. Arch. Microbiol. 2020, 202, 2071–2081. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, M.; Shimoyama, Y.; Kodama, Y.; Ishikawa, T. Tryptophanyl tRNA Synthetase from Human Macrophages Infected by Porphyromonas gingivalis Induces a Proinflammatory Response Associated with Atherosclerosis. Pathogens 2021, 10, 1648. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121648
Sasaki M, Shimoyama Y, Kodama Y, Ishikawa T. Tryptophanyl tRNA Synthetase from Human Macrophages Infected by Porphyromonas gingivalis Induces a Proinflammatory Response Associated with Atherosclerosis. Pathogens. 2021; 10(12):1648. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121648
Chicago/Turabian StyleSasaki, Minoru, Yu Shimoyama, Yoshitoyo Kodama, and Taichi Ishikawa. 2021. "Tryptophanyl tRNA Synthetase from Human Macrophages Infected by Porphyromonas gingivalis Induces a Proinflammatory Response Associated with Atherosclerosis" Pathogens 10, no. 12: 1648. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121648