
Replication of Influenza A Virus in Secondary Lymphatic Tissue Contributes to Innate Immune Activation

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Mice
2.2. Virus Quantification
2.3. Reagents
2.4. Immunofluorescense
2.5. Flow Cytometry
2.6. Intracellular Cytokine and IAV Staining
2.7. RNA Extraction, cDNA Synthesis and qRT-PCR
2.8. ELISA
2.9. Statistical Analysis
3. Results
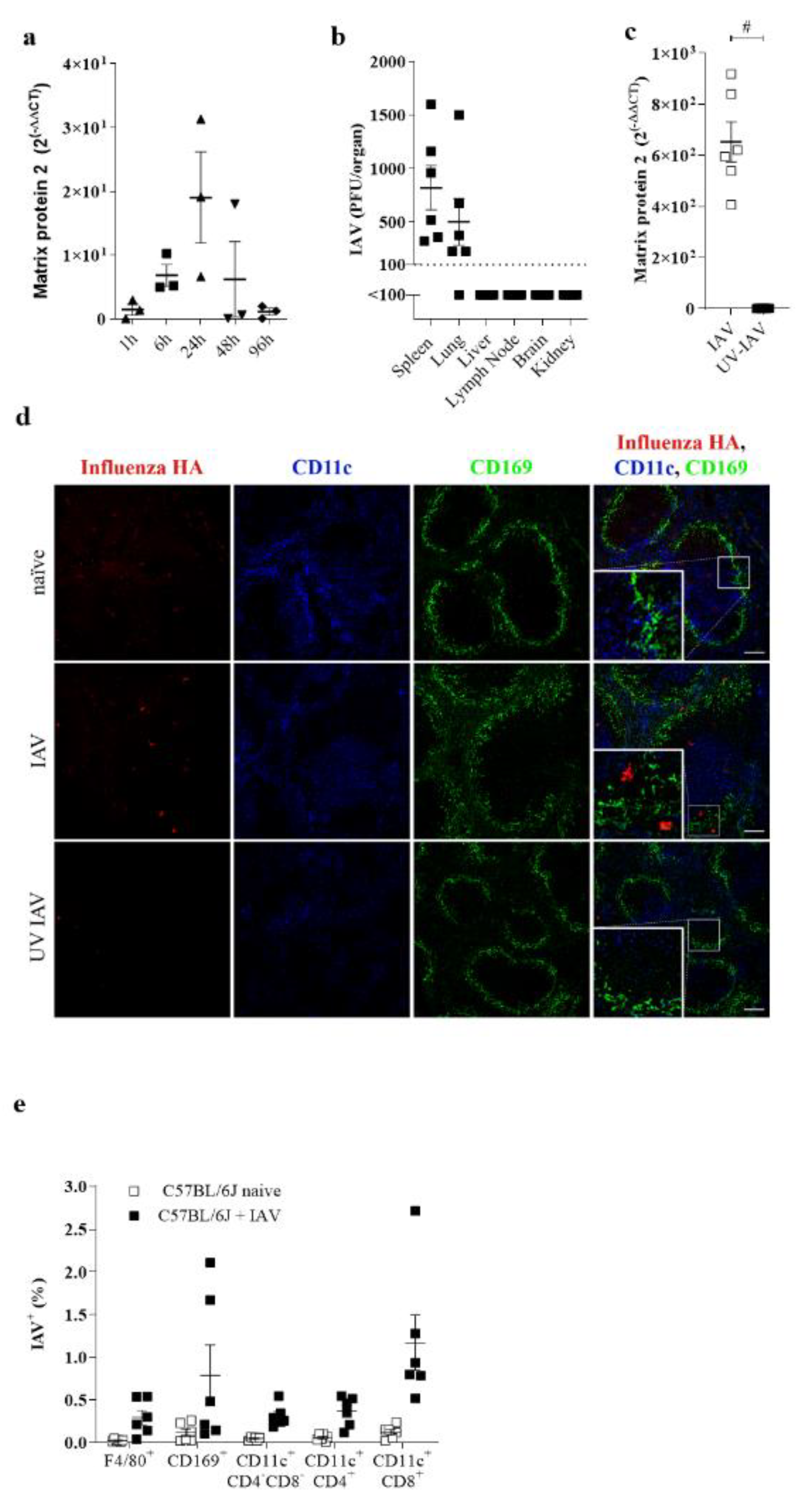
3.1. IAV Replicates in the Spleen during Systemic Infection
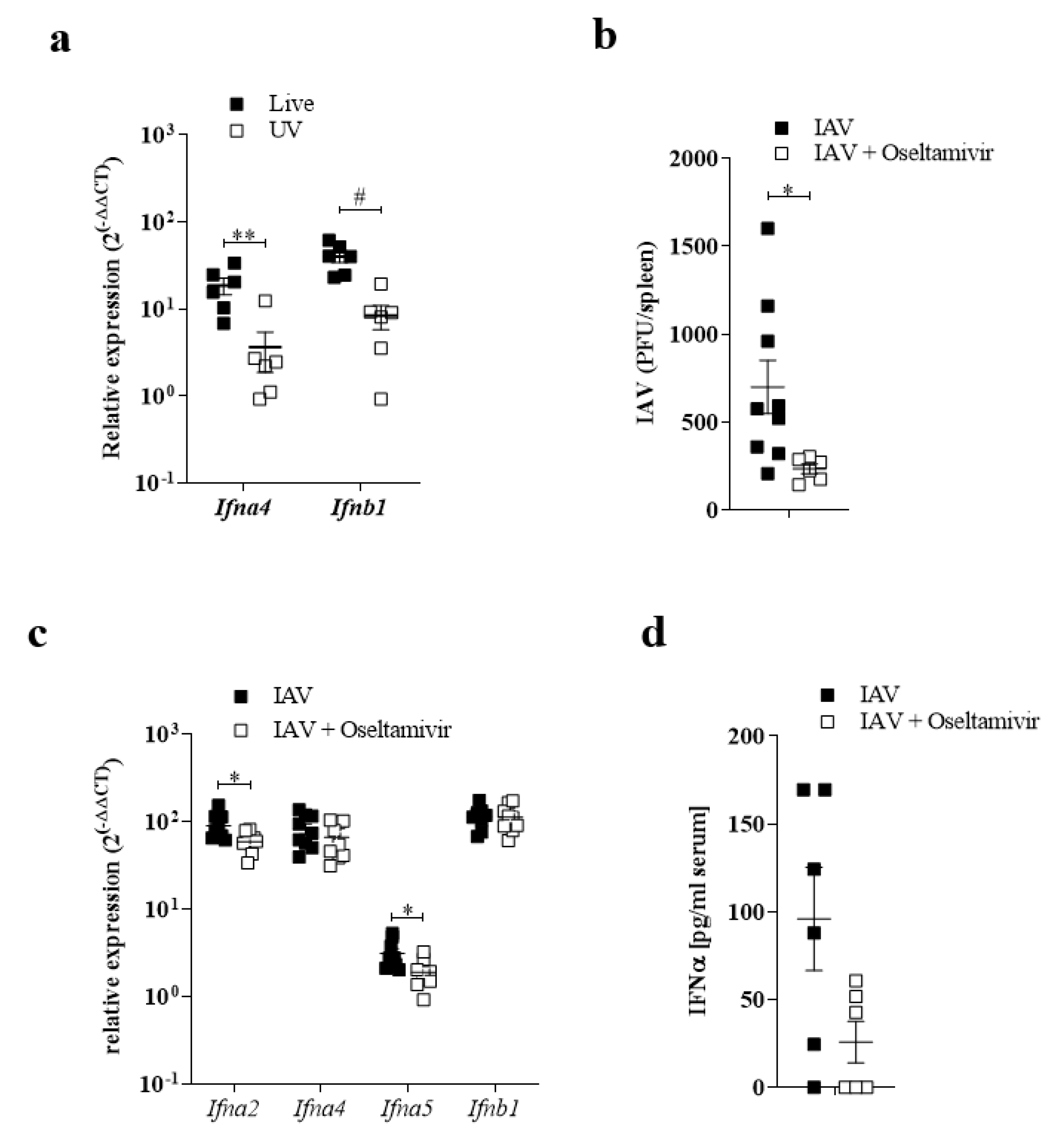
3.2. Replication of IAV in the Spleen Correlated with IFN-I Induction
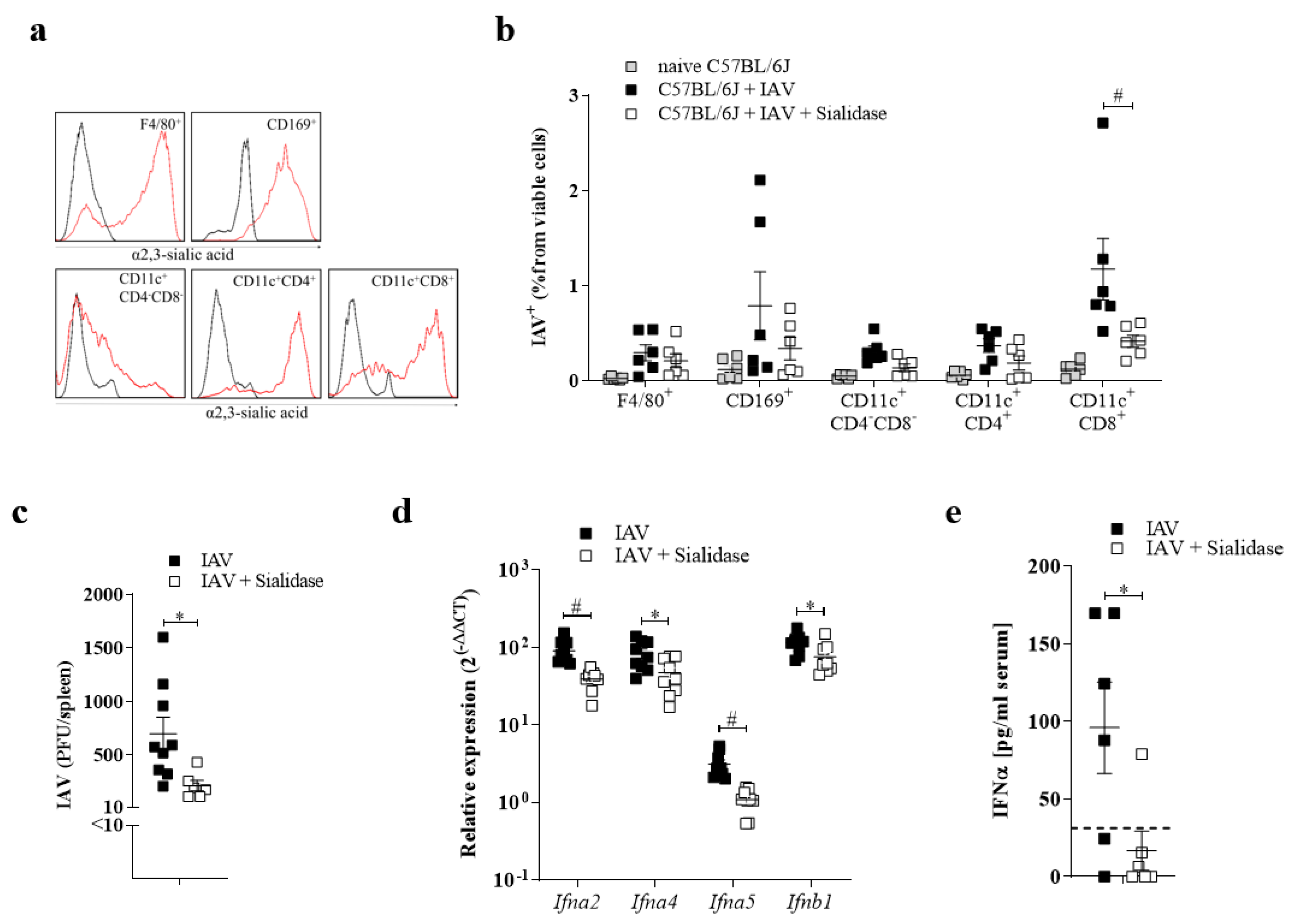
3.3. Sialic Acid Residues Are Essential for IAV Uptake
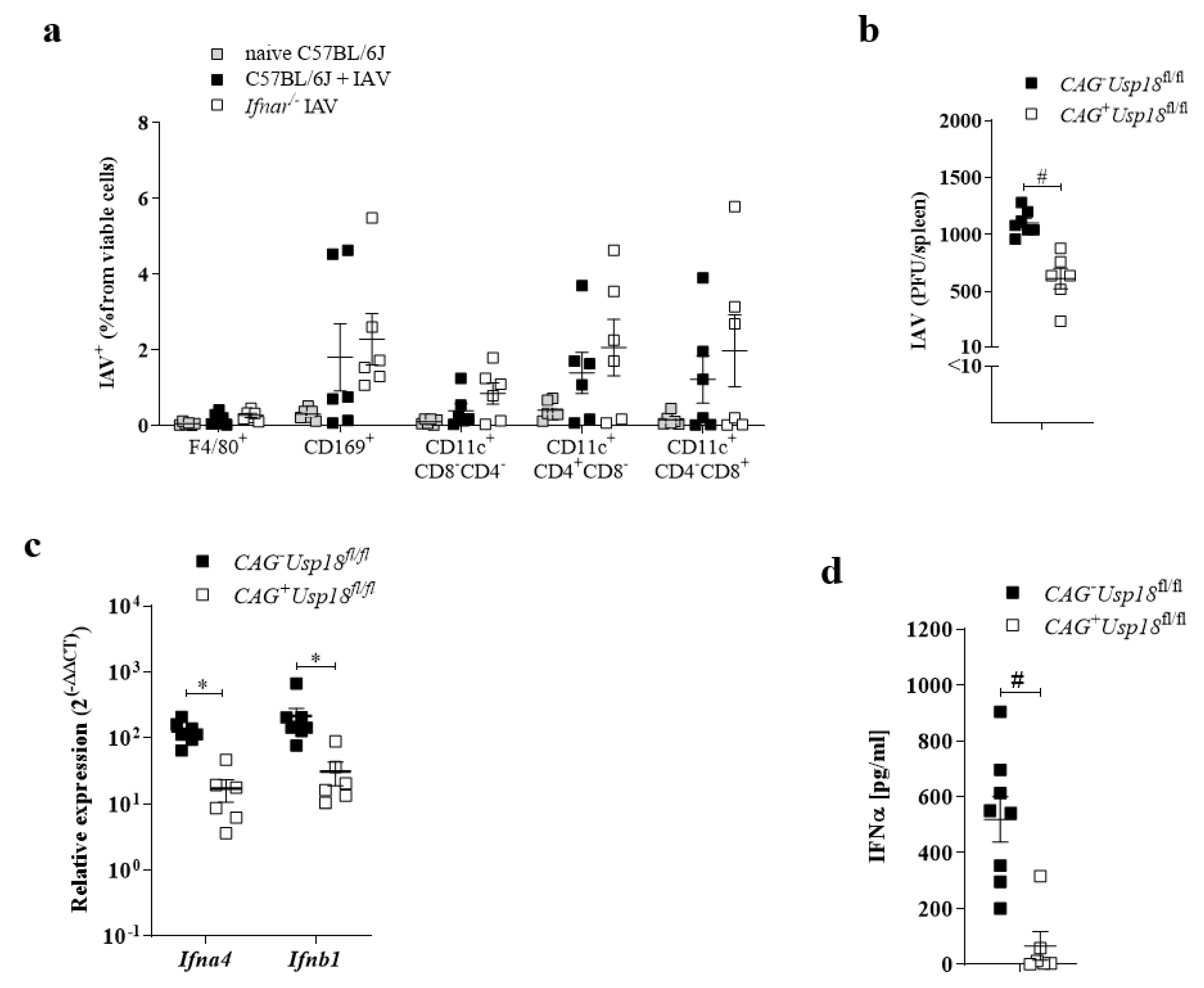
3.4. Usp18 Promotes Replication of IAV in Antigen-Presenting Cells
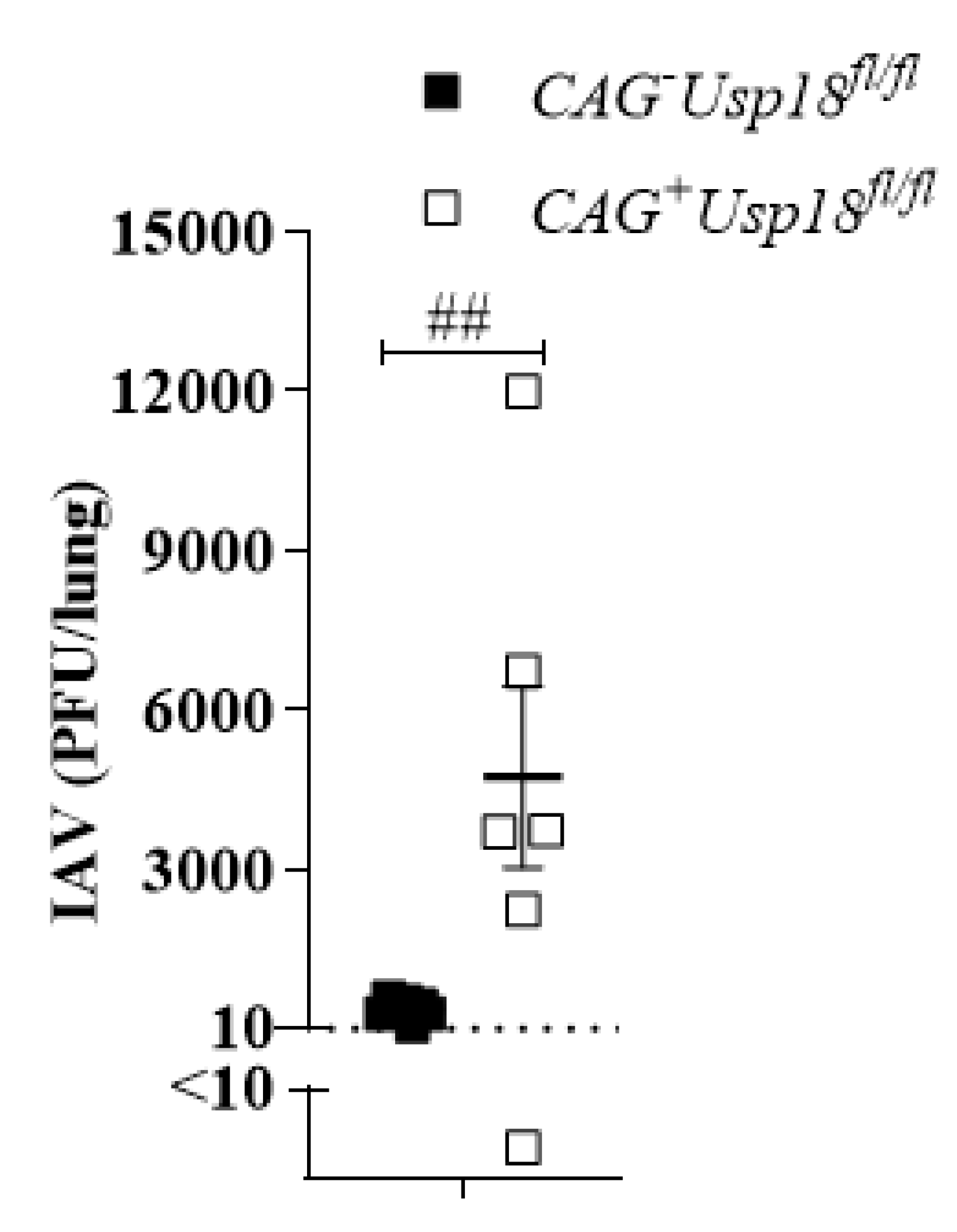
3.5. Usp18 Dependent Replication Controls Virus Propagation in the Lung
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hopp, A.K.; Rupp, A.; Lukacs-Kornek, V. Self-antigen presentation by dendritic cells in autoimmunity. Front. Immunol. 2014, 5, 55. [Google Scholar] [CrossRef] [Green Version]
- Lehuen, A.; Diana, J.; Zaccone, P.; Cooke, A. Immune cell crosstalk in type 1 diabetes. Nat. Rev. Immunol. 2010, 10, 501–513. [Google Scholar] [CrossRef]
- Turley, S.J.; Fletcher, A.L.; Elpek, K.G. The stromal and haematopoietic antigen-presenting cells that reside in secondary lymphoid organs. Nat. Rev. Immunol. 2010, 10, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Schabbauer, G.; Luyendyk, J.; Crozat, K.; Jiang, Z.; Mackman, N.; Bahram, S.; Georgel, P. TLR4/CD14-mediated PI3K activation is an essential component of interferon-dependent VSV resistance in macrophages. Mol. Immunol. 2008, 45, 2790–2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeon, S.H.; Song, M.J.; Kang, H.-R.; Lee, J.Y. Phosphatidylinositol-3-kinase and Akt are required for RIG-I-mediated anti-viral signalling through cross-talk with IPS-1. Immunology 2015, 144, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Uematsu, S.; Akira, S. Toll-like receptors and Type I interferons. J. Biol. Chem. 2007, 282, 15319–15323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colonna, M. TLR pathways and IFN-regulatory factors: To each its own. Eur. J. Immunol. 2007, 37, 306–309. [Google Scholar] [CrossRef]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- York, A.G.; Williams, K.J.; Argus, J.P.; Zhou, Q.D.; Brar, G.; Vergnes, L.; Gray, E.E.; Zhen, A.; Wu, N.C.; Yamada, D.H.; et al. Limiting Cholesterol Biosynthetic Flux Spontaneously Engages Type I IFN Signaling. Cell 2015, 163, 1716–1729. [Google Scholar] [CrossRef] [Green Version]
- Honke, N.; Shaabani, N.; Cadeddu, G.; Sorg, U.R.; Zhang, D.-E.; Trilling, M.; Klingel, K.; Sauter, M.; Kandolf, R.; Gailus, N.; et al. Enforced viral replication activates adaptive immunity and is essential for the control of a cytopathic virus. Nat. Immunol. 2011, 13, 51–57. [Google Scholar] [CrossRef]
- Honke, N.; Shaabani, N.; Zhang, D.-E.; Iliakis, G.; Xu, H.C.; Häussinger, D.; Recher, M.; Löhning, M.; Lang, P.A.; Lang, K.S. Usp18 driven enforced viral replication in dendritic cells contributes to break of immunological tolerance in autoimmune diabetes. PLoS Pathog. 2013, 9, e1003650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaabani, N.; Khairnar, V.; Duhan, V.; Zhou, F.; Tur, R.F.; Häussinger, D.; Recher, M.; Tumanov, A.V.; Hardt, C.; Pinschewer, D.; et al. Two separate mechanisms of enforced viral replication balance innate and adaptive immune activation. J. Autoimmun. 2016, 67, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, J.M.; Kim, J.; Tenson, T.; Min, J.-Y.; Kainov, D.E. Influenza Virus Infection, Interferon Response, Viral Counter-Response, and Apoptosis. Viruses 2017, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Arias, C.F.; Escalera-Zamudio, M.; Río, M.D.L.D.S.-D.; Cobián-Güemes, A.G.; Isa, P.; López, S. Molecular Anatomy of 2009 Influenza Virus A (H1N1). Arch. Med. Res. 2009, 40, 643–654. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Goraya, M.U.; Maarouf, M.; Huang, S.; Chen, J.-L. Host Immune Response to Influenza A Virus Infection. Front. Immunol. 2018, 9, 320. [Google Scholar] [CrossRef] [Green Version]
- Lang, K.S.; Recher, M.; Junt, T.; Navarini, A.A.; Harris, N.L.; Freigang, S.; Odermatt, B.; Conrad, C.; Ittner, L.M.; Bauer, S.; et al. Toll-like receptor engagement converts T-cell autoreactivity into overt autoimmune disease. Nat. Med. 2005, 11, 138–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, S.K.; Schmitz, R.; Bergerhausen, M.; Lang, J.; Cham, L.B.; Duhan, V.; Häussinger, D.; Hardt, C.; Addo, M.; Prinz, M.; et al. Usp18 Expression in CD169+ Macrophages is Important for Strong Immune Response after Vaccination with VSV-EBOV. Vaccines 2020, 8, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldmann, T.; Zeller, N.; Raasch, J.; Kierdorf, K.; Frenzel, K.; Ketscher, L.; Basters, A.; Staszewski, O.; Brendecke, S.M.; Spiess, A.; et al. USP 18 lack in microglia causes destructive interferonopathy of the mouse brain. EMBO J. 2015, 34, 1612–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, K.; Takahashi, N.; Ushiki, M.; Monya, M.; Aihara, F.; Kuboki, E.; Moriyama, S.; Iida, M.; Kitamura, H.; Qiu, C.-H.; et al. Intestinal CD169+ macrophages initiate mucosal inflammation by secreting CCL8 that recruits inflammatory monocytes. Nat. Commun. 2015, 6, 7802. [Google Scholar] [CrossRef] [PubMed]
- Bird, N.L.; Olson, M.R.; Hurt, A.C.; Oshansky, C.M.; Oh, D.Y.; Reading, P.C.; Chua, B.Y.; Sun, Y.; Tang, L.; Handel, A.; et al. Oseltamivir Prophylaxis Reduces Inflammation and Facilitates Establishment of Cross-Strain Protective T Cell Memory to Influenza Viruses. PLoS ONE 2015, 10, e0129768. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Seyfried, T.N.; Choi, H.-S.; Kidd, S.K. Intraventricular Sialidase Administration Enhances GM1 Ganglioside Expression and Is Partially Neuroprotective in a Mouse Model of Parkinson’s Disease. PLoS ONE 2015, 10, e0143351. [Google Scholar] [CrossRef] [PubMed]
- Murali-Krishna, K.; Altman, J.D.; Suresh, M.; Sourdive, D.J.; Zajac, A.J.; Miller, J.D.; Slansky, J.; Ahmed, R. Counting Antigen-Specific CD8 T Cells: A Reevaluation of Bystander Activation during Viral Infection. Immunity 1998, 8, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.U.; Lew, W.; Williams, M.A.; Liu, H.; Zhang, L.; Swaminathan, S.; Bischofberger, N.; Chen, M.S.; Mendel, D.B.; Tai, C.Y.; et al. Influenza neuraminidase inhibitors possessing a novel hydrophobic interaction in the enzyme active site: Design, synthesis, and structural analysis of carbocyclic sialic acid analogues with potent anti-influenza activity. J. Am. Chem. Soc. 1997, 119, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Konig, R.; Stertz, S.; Zhou, Y.; Inoue, A.; Hoffmann, H.-H.; Bhattacharyya, S.; Alamares, J.G.; Tscherne, D.M.; Ortigoza, M.B.; Liang, Y.; et al. Human host factors required for influenza virus replication. Nature 2010, 463, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Nikitina, E.; Larionova, I.; Choinzonov, E.; Kzhyshkowska, J. Monocytes and Macrophages as Viral Targets and Reservoirs. Int. J. Mol. Sci. 2018, 19, 2821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, M.D.; Brooks, A.G.; Reading, P.C. Correlation between sialic acid expression and infection of murine macrophages by different strains of influenza virus. Microbes Infect. 2011, 13, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.S.Y.; Li, O.T.W.; Chan, R.W.Y.; Chan, M.C.W.; Nicholls, J.M.; Poon, L.L.M. Entry of influenza A Virus with a α2,6-linked sialic acid binding preference requires host fibronectin. J. Virol. 2012, 86, 10704–10713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.-T.; Takahashi, N.; Yagi, H.; Kato, K.; Takahashi, T.; Yi, S.-Q.; Chen, Y.; Ito, T.; Otsuki, K.; Kida, H.; et al. The quail and chicken intestine have sialyl-galactose sugar chains responsible for the binding of influenza A viruses to human type receptors. Glycobiology 2007, 17, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honke, N.; Shaabani, N.; Zhang, D.-E.; Hardt, C.; Lang, K.S. Multiple functions of Usp18. Cell Death Dis. 2016, 7, e2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, K.S.; Lang, P.A. Balancing viral replication in spleen and liver determines the outcome of systemic virus infection. Z. Gastroenterol. 2015, 53, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Kuffour, E.O.; Schott, K.; Vasudevan, A.A.J.; Holler, J.; Schulz, W.A.; Lang, P.A.; Lang, K.S.; Kim, B.; Häussinger, D.; König, R.; et al. Usp18 (UBP43) Abrogates p21-Mediated Inhibition of HIV-1. J. Virol. 2018, 92, e00592-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minor, P.D. Live attenuated vaccines: Historical successes and current challenges. Virology 2015, 479–480, 379–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, P.A.; Cervantes-Barragan, L.; Verschoor, A.; Navarini, A.A.; Recher, M.; Pellegrini, M.; Flatz, L.; Bergthaler, A.; Honda, K.; Ludewig, B.; et al. Hematopoietic cell–derived interferon controls viral replication and virus-induced disease. Blood 2009, 113, 1045–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholls, J.M.; Bourne, A.J.; Chen, H.; Guan, Y.; Peiris, J.M. Sialic acid receptor detection in the human respiratory tract: Evidence for widespread distribution of potential binding sites for human and avian influenza viruses. Respir. Res. 2007, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, W.; Tabi, Z.; Cleary, A.; Doherty, P.C. Cellular events in the lymph node and lung of mice with influenza. Consequences of depleting CD4+ T cells. J. Immunol. 1990, 144, 3980–3986. [Google Scholar] [PubMed]
- Chatziandreou, N.; Farsakoglu, Y.; Palomino-Segura, M.; D’Antuono, R.; Pizzagalli, D.U.; Sallusto, F.; Lukacs-Kornek, V.; Uguccioni, M.; Corti, D.; Turley, S.J.; et al. Macrophage Death following Influenza Vaccination Initiates the Inflammatory Response that Promotes Dendritic Cell Function in the Draining Lymph Node. Cell Rep. 2017, 18, 2427–2440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, X.; Lu, J.; Wang, H.; Liu, X.; Wang, J.; Zhou, K.; Jiang, W.; Wang, Y.; Fang, M. Bidirectional factors impact the migration of NK cells to draining lymph node in aged mice during influenza virus infection. Exp. Gerontol. 2017, 96, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Elsner, R.A.; Ernst, D.N.; Baumgarth, N. Single and coexpression of CXCR4 and CXCR5 identifies CD4 T helper cells in distinct lymph node niches during influenza virus infection. J. Virol. 2012, 86, 7146–7157. [Google Scholar] [CrossRef] [Green Version]
- Hamilton-Easton, A.; Eichelberger, M. Virus-specific antigen presentation by different subsets of cells from lung and mediastinal lymph node tissues of influenza virus-infected mice. J. Virol. 1995, 69, 6359–6366. [Google Scholar] [CrossRef] [Green Version]
- Ho, A.W.; Prabhu, N.; Betts, R.J.; Ge, M.Q.; Dai, X.; Hutchinson, P.E.; Lew, F.C.; Wong, K.L.; Hanson, B.J.; Macary, P.A.; et al. Lung CD103+ dendritic cells efficiently transport influenza virus to the lymph node and load viral antigen onto MHC class I for presentation to CD8 T cells. J. Immunol. 2011, 187, 6011–6021. [Google Scholar] [CrossRef] [Green Version]
- Khanna, K.M.; Aguila, C.C.; Redman, J.M.; Suarez-Ramirez, J.E.; Lefrançois, L.; Cauley, L.S. In situ imaging reveals different responses by naïve and memory CD8 T cells to late antigen presentation by lymph node DC after influenza virus infection. Eur. J. Immunol. 2008, 38, 3304–3315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez-Ramírez, J.E.; Wu, T.; Lee, Y.-T.; Aguila, C.C.; Bouchard, K.R.; Cauley, L.S. Division of labor between subsets of lymph node dendritic cells determines the specificity of the CD8+ T-cell recall response to influenza infection. Eur. J. Immunol. 2011, 41, 2632–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.V.; McGill, J.; Legge, K.L. Quantification of the frequency and multiplicity of infection of respiratory- and lymph node-resident dendritic cells during influenza virus infection. PLoS ONE 2010, 5, e12902. [Google Scholar]
- Lund, F.E.; Partida-Sánchez, S.; Lee, B.O.; Kusser, K.L.; Hartson, L.; Hogan, R.J.; Woodland, D.L.; Randall, T.D. Lymphotoxin-alpha-deficient mice make delayed, but effective, T and B cell responses to influenza. J. Immunol. 2002, 169, 5236–5243. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friedrich, S.-K.; Schmitz, R.; Bergerhausen, M.; Lang, J.; Duhan, V.; Hardt, C.; Tenbusch, M.; Prinz, M.; Asano, K.; Bhat, H.; et al. Replication of Influenza A Virus in Secondary Lymphatic Tissue Contributes to Innate Immune Activation. Pathogens 2021, 10, 622. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050622
Friedrich S-K, Schmitz R, Bergerhausen M, Lang J, Duhan V, Hardt C, Tenbusch M, Prinz M, Asano K, Bhat H, et al. Replication of Influenza A Virus in Secondary Lymphatic Tissue Contributes to Innate Immune Activation. Pathogens. 2021; 10(5):622. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050622
Chicago/Turabian StyleFriedrich, Sarah-Kim, Rosa Schmitz, Michael Bergerhausen, Judith Lang, Vikas Duhan, Cornelia Hardt, Matthias Tenbusch, Marco Prinz, Kenichi Asano, Hilal Bhat, and et al. 2021. "Replication of Influenza A Virus in Secondary Lymphatic Tissue Contributes to Innate Immune Activation" Pathogens 10, no. 5: 622. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050622