
Molecular Detection of Bartonella quintana among Long-Tailed Macaques (Macaca fascicularis) in Thailand

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Molecular Identification of Bartonella spp.
2.2. Nucleotide Sequencing
2.3. Nucleotide Sequence Accession Numbers
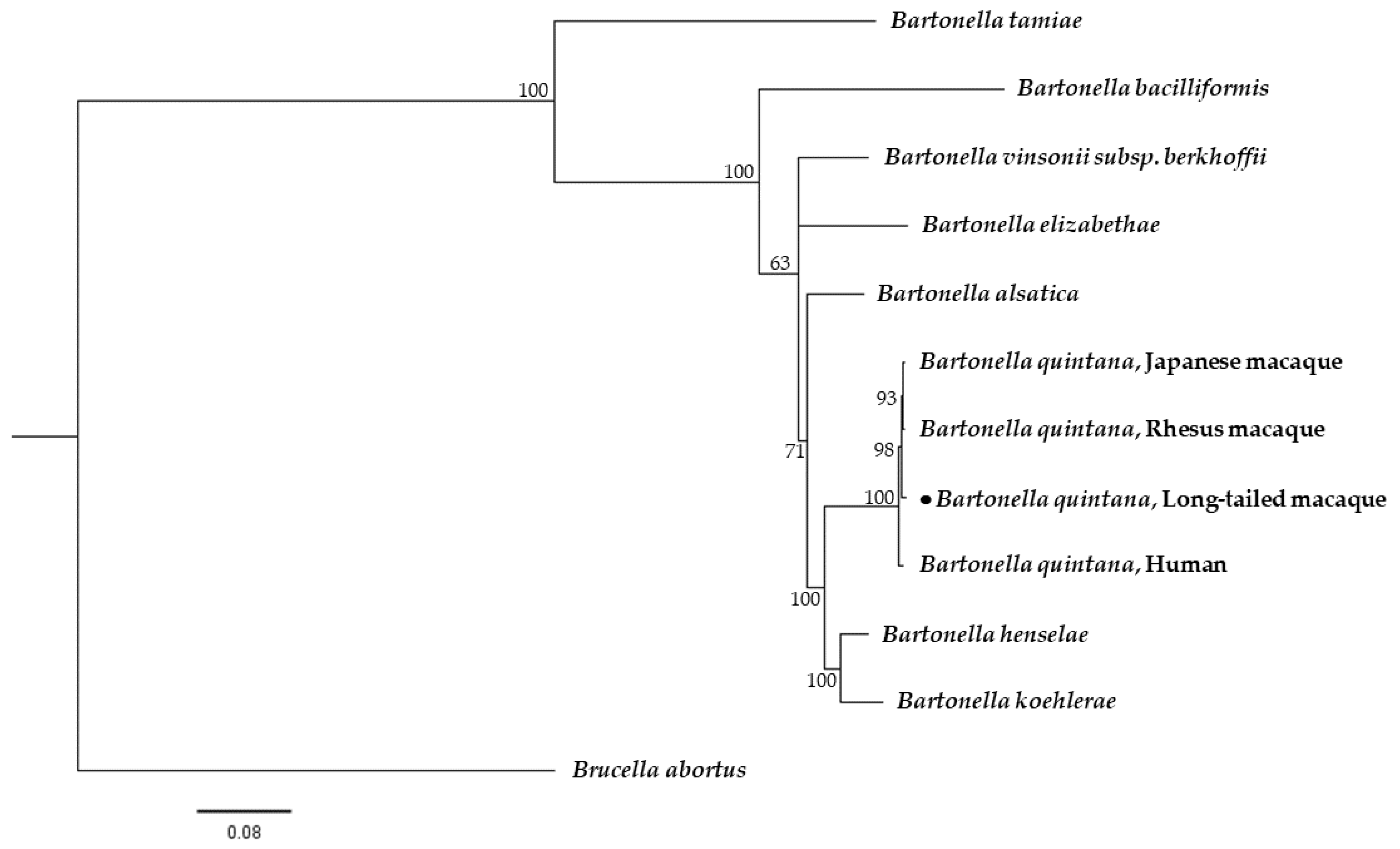
2.4. Phylogenetic Analysis
2.5. Multiple Sequence Alignment Analysis
3. Discussion
4. Materials and Methods
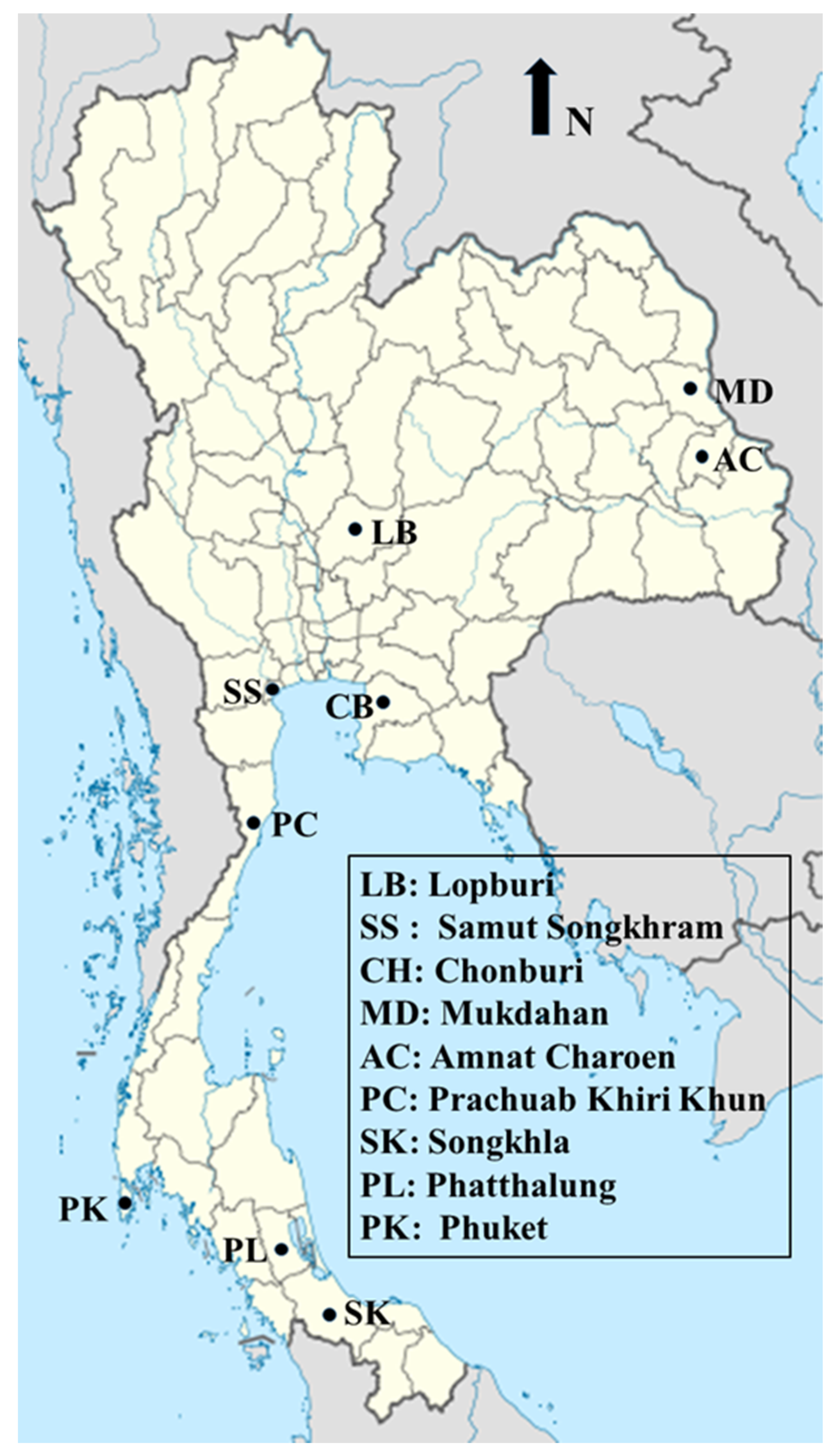
4.1. Study Areas and Sample Collection
4.2. Molecular Analysis
4.3. Phylogenetic Analysis
4.4. Multiple Sequence Alignment Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breitschwerdt, E.B. Bartonellosis: One health perspectives for an emerging infectious disease. ILAR J. 2014, 55, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Breitschwerdt, E.B. Bartonellosis, one health and all creatures great and small. In Advances in Veterinary Dermatology; Torres, S.M., Roudebush, P., Eds.; Wiley: Oxford, UK, 2017; pp. 111–121. [Google Scholar]
- Cheslock, M.A.; Embers, M.E. Human bartonellosis: An underappreciated public health problem? Trop. Med. Infect. Dis. 2019, 4, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, J. Bartonella quintana, past, present, and future of the scourge of World War I. APMIS 2018, 126, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.C.; Fonseca, K.; Pabbaraju, K.; Meatherall, B.L. Case report: Bartonella quintana endocarditis outside of the Europe–African gradient: Comprehensive review of cases within North America. Am. J. Trop. Med. Hyg. 2019, 100, 1125–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Bai, J.Y.; Wang, L.Y.; Zeng, L.; Shi, Y.S.; Qiu, Z.L.; Ye, H.H.; Zhang, X.F.; Lu, Q.B.; Kosoy, M.; et al. Genetic diversity of Bartonella quintana in macaques suggests zoonotic origin of trench fever. Mol. Ecol. 2013, 22, 2118–2127. [Google Scholar] [CrossRef]
- Li, H.; Liu, W.; Zhang, G.Z.; Sun, Z.Z.; Bai, J.Y.; Jiang, B.G.; Zhang, Y.Y.; Zhao, X.G.; Yang, H.; Tian, G. Transmission and maintenance cycle of Bartonella quintana among rhesus macaques, China. Emerg. Infect. Dis. 2013, 19, 297–300. [Google Scholar] [CrossRef]
- O’Rourke, L.G.; Pitulle, C.; Hegarty, B.C.; Kraycirik, S.; Killary, K.A.; Grosenstein, P.; Brown, J.W.; Breitschwerdt, E.B. Bartonella quintana in cynomolgus monkey (Macaca fascicularis). Emerg. Infect. Dis. 2005, 11, 1931–1934. [Google Scholar] [CrossRef] [Green Version]
- Maggi, R.G.; Mascarelli, P.E.; Balakrishnan, N.; Rohde, C.M.; Kelly, C.M.; Ramaiah, L.; Leach, M.W.; Breitschwerdt, E.B. Candidatus Mycoplasma haemomacaque and Bartonella quintana bacteremia in cynomolgus monkeys. J. Clin. Microbiol. 2013, 51, 1408–1411. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Liu, Q.; Li, G.; Li, D.; Song, X.; Birtles, R.J.; Zhao, F. Bartonella quintana infections in captive monkeys, China. Emerg. Infect. Dis. 2011, 17, 1707–1709. [Google Scholar] [CrossRef]
- Sato, S.; Kabeya, H.; Yoshino, A.; Sekine, W.; Suzuki, K.; Tamate, H.B.; Yamazaki, S.; Chomel, B.B.; Maruyama, S. Japanese macaques (Macaca fuscata) as natural reservoir of Bartonella quintana. Emerg. Infect. Dis. 2015, 21, 2168–2170. [Google Scholar] [CrossRef]
- Malaivijitnond, S.; Hamada, Y. Current situation and status of long-tailed macaques (Macaca fascicularis) in Thailand. Nat. Hist. J. Chulalongkorn Univ. 2008, 8, 185–204. [Google Scholar]
- Roos, C.; Zinner, D. Chapter 1—Diversity and evolutionary history of macaques with special focus on Macaca mulatta and Macaca fascicularis. In The Nonhuman Primate in Nonclinical drug Development and Safety Assessment; Bluemel, J., Korte, S., Schenck, E., Weinbauer, G.F., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 3–16. [Google Scholar]
- Norman, A.F.; Regnery, R.; Jameson, P.; Greene, C.; Krause, D.C. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J. Clin. Microbiol. 1995, 33, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- Renesto, P.; Gouvernet, J.; Drancourt, M.; Roux, V.; Raoult, D. Use of rpoB gene analysis for detection and identification of Bartonella species. J. Clin. Microbiol. 2001, 39, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeaiter, Z.; Liang, Z.; Raoult, D. Genetic classification and differentiation of Bartonella species based on comparison of partial ftsZ gene sequences. J. Clin. Microbiol. 2002, 40, 3641–3647. [Google Scholar] [CrossRef] [Green Version]
- Zeaiter, Z.; Fournier, P.E.; Ogata, H.; Raoult, D. Phylogenetic classification of Bartonella species by comparing groEL sequences. Int. J. Syst. Evol. Microbiol. 2002, 52, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.; Ayers, M.; McClure, S.C.C.; Richardson, S.E.; Tellier, R. Detection and identification of Bartonella species pathogenic for humans by PCR amplification targeting the riboflavin synthase gene (ribC). J. Clin. Microbiol. 2003, 41, 1069–1072. [Google Scholar] [CrossRef] [Green Version]
- Maggi, R.G.; Breitschwerdt, E.B. Potential limitations of the 16S-23S rRNA intergenic region for molecular detection of Bartonella species. J. Clin. Microbiol. 2005, 43, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Bonato, L.; Figueiredo, M.A.P.; Gonçalves, L.R.; Machado, R.Z.; André, M.R. Occurrence and molecular characterization of Bartonella spp. and hemoplasmas in neotropical primates from Brazilian Amazon. Comp. Immunol. Microbiol. Infect. Dis. 2015, 42, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Pitassi, L.H.U.; de Paiva Diniz, P.P.V.; Scorpio, D.G.; Drummond, M.R.; Lania, B.G.; Barjas-Castro, M.L.; Gilioli, R.; Colombo, S.; Sowy, S.; Breitschwerdt, E.B.; et al. Bartonella spp. bacteremia in blood donors from Campinas, Brazil. PLoS Negl. Trop. Dis. 2015, 9, e0003467. [Google Scholar] [CrossRef] [Green Version]
- Oteo, J.A.; Maggi, R.; Portillo, A.; Bradley, J.; García-Álvarez, L.; San-Martín, M.; Roura, X.; Breitschwerdt, E. Prevalence of Bartonella spp. by culture, PCR and serology, in veterinary personnel from Spain. Parasit. Vectors. 2017, 10, 553. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Kosoy, M.Y.; Boonmar, S.; Sawatwong, P.; Sangmaneedet, S.; Peruski, L.F. Enrichment culture and molecular identification of diverse Bartonella species in stray dogs. Vet. Microbiol. 2010, 146, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, M.; Bai, Y.; Sheff, K.; Morway, C.; Baggett, H.; Maloney, S.A.; Boonmar, S.; Bhengsri, S.; Dowell, S.F.; Sitdhirasdr, A.; et al. Identification of Bartonella infections in febrile human patients from Thailand and their potential animal reservoirs. Am. J. Trop. Med. Hyg. 2010, 82, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Breitschwerdt, E.B.; Maggi, R.G.; Sigmon, B.; Nicholson, W.L. Isolation of Bartonella quintana from a woman and a cat following putative bite transmission. J. Clin. Microbiol. 2007, 45, 270–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaewmongkol, G.; Maneesaay, P.; Suwanna, N.; Tiraphut, B.; Krajarngjang, T.; Chouybumrung, A.; Kaewmongkol, S.; Sirinarumitr, T.; Jittapalapong, S.; Fenwick, S. First detection of Ehrlichia canis in cerebrospinal fluid from a nonthrombocytopenic dog with meningoencephalitis by broad-range PCR. J. Vet. Intern. Med. 2016, 30, 255–259. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession Number | Homology (%) | Nucleotide at Position | ||||
|---|---|---|---|---|---|---|---|
| gltA | 618 | 648 | |||||
| Reference | JQ314417 | 99.35% | C | A | |||
| MW320725 | T | G | |||||
| ftsZ | 1255 | 1453 | 1530 | 1677 | 1725 | ||
| Reference | JQ314416 | 99.40% | A | C | C | T | G |
| MW341113 | G | T | A | G | A | ||
| Region | Province | Male | Female | Total Number |
|---|---|---|---|---|
| Central | Lopburi | 81 | 112 | 193 |
| Samut Songkhram | 18 | 7 | 25 | |
| East | Chonburi | 253 | 47 | 300 |
| Northeast | Mukdahan | 30 | 0 | 30 |
| Amnat Charoen | 70 | 29 | 99 | |
| West | Prachuab Khiri Khun | 28 | 38 | 66 |
| South | Songkhla | 34 | 5 | 39 |
| Phatthalung | 30 | 0 | 30 | |
| Phuket | 67 | 7 | 74 | |
| Total | 611 | 245 | 856 |
| Gene | Primer | Oligonucleotide Sequences | Amplicon Size (bp) | Ref. |
|---|---|---|---|---|
| 16S rRNA | V1-F(a) | 5′-AGAGTTTGATCCTGGCTCAG-3′ | 1400 | [26] |
| V9-R(a) | 5′-GNTACCTTGTTACGACTT-3′ | |||
| V3-F(b) | 5′-ACTCCTACGGGAGGCAGCAG-3′ | 700 | ||
| V6-R(b) | 5′-CGACAGCCATGCANCACCT-3′ | |||
| gltA | BhCS.781p | 5′-GGGGACCAGCTCATGGTGG-3′ | 390 | [14] |
| BhCS.1137n | 5′-AATGCAAAAAGAACAGTAAACA-3′ | |||
| ftsZ | BaftsZF | 5′-GCTAATCGTATTCGCGAAGAA-3′ | 900 | [16] |
| BaftsZR | 5′-GCTGGTATTTCCAAYTGATCT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sricharern, W.; Kaewchot, S.; Saengsawang, P.; Kaewmongkol, S.; Inpankaew, T. Molecular Detection of Bartonella quintana among Long-Tailed Macaques (Macaca fascicularis) in Thailand. Pathogens 2021, 10, 629. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050629
Sricharern W, Kaewchot S, Saengsawang P, Kaewmongkol S, Inpankaew T. Molecular Detection of Bartonella quintana among Long-Tailed Macaques (Macaca fascicularis) in Thailand. Pathogens. 2021; 10(5):629. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050629
Chicago/Turabian StyleSricharern, Wanat, Supakarn Kaewchot, Phirabhat Saengsawang, Sarawan Kaewmongkol, and Tawin Inpankaew. 2021. "Molecular Detection of Bartonella quintana among Long-Tailed Macaques (Macaca fascicularis) in Thailand" Pathogens 10, no. 5: 629. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050629