
Cryptosporidium: Still Open Scenarios


1
Department of Diagnostic and Laboratory Medicine, Bambino Gesù Children’s Hospital, IRCCS, Unit of Microbiology and Diagnostic Immunology, Unit of Microbiomics, 00146 Rome, Italy
2
Department of Diagnostic and Laboratory Medicine, Bambino Gesù Children’s Hospital, IRCCS, Unit of Microbiology and Diagnostic Immunology, Unit of Microbiomics and Multimodal Laboratory Medicine Research Area, Unit of Human Microbiome, 00146 Rome, Italy
*
Author to whom correspondence should be addressed.
Pathogens 2022, 11(5), 515; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050515
Submission received: 8 March 2022
/
Revised: 20 April 2022
/
Accepted: 23 April 2022
/
Published: 26 April 2022
(This article belongs to the Special Issue Intestinal Parasites Infection)
Abstract
:Cryptosporidiosis is increasingly identified as a leading cause of childhood diarrhea and malnutrition in both low-income and high-income countries. The strong impact on public health in epidemic scenarios makes it increasingly essential to identify the sources of infection and understand the transmission routes in order to apply the right prevention or treatment protocols. The objective of this literature review was to present an overview of the current state of human cryptosporidiosis, reviewing risk factors, discussing advances in the drug treatment and epidemiology, and emphasizing the need to identify a government system for reporting diagnosed cases, hitherto undervalued.
Keywords:
Cryptosporidium; zoonosis; humans; animals; transmission; epidemiology; treatment; drug; vaccine1. Introduction
In the early 1980s, Cryptosporidium spp. was observed as a pathogen affecting both humans and animals and as a cause of acute gastroenteritis, abdominal pain, and diarrhea [1]. Cryptosporidiosis is mainly transmitted via the fecal-oral, zoonotic, or anthroponotic route, through contaminated water or food [2]. Due to the presence of multiple transmission routes, the epidemiology of cryptosporidiosis is complex (Figure 1). Indeed, the investigation of both sporadic cases and outbreaks has contributed to a better understanding of risk factors and infection sources [3]. For example, cryptosporidiosis is increasingly identified as the leading cause of chronic diarrhea in immunocompromised patients and as a cause of infant malnutrition and premature death in children [4]. Despite this, parasitosis is substantially underdiagnosed and underestimated, lacks standardized preventive protocols, and requires optimal treatment on a large scale.
Today, nitazoxanide is the only drug approved by the US Food and Drug Administration (FDA), European Medicines Agency (EMA), and Pharmaceuticals and Medical Devices Agency (PMDA) for the treatment of cryptosporidiosis in immunocompetent people, but its use is not conclusive in young children and immunocompromised individuals [5,6,7], thus having a deep impact on public health today.
2. Epidemiology of Cryptosporidium: Recent Updates
Over a million deaths from gastroenteritis associated to Cryptosporidium spp. have been described in the last quarter of the 20th century and at the beginning of the 21st century [8].
However, the impact of this protozoan parasite remains more widespread in low income countries, where endemicity, poor sanitation, and limited access to filtered water limit diagnostic practice and health care, determining high morbidity and mortality rates, particularly in children under five years of age [9]. The impact of variability in the prevalence of cryptosporidiosis between underdeveloped and industrialized countries makes it difficult to interpret surveillance data and reinforce the necessity to promote prompt diagnosis and improve related methods. The development of 18s rRNA/gp60 gene sequencing has been reported to be useful in the investigation and surveillance of outbreaks and for the identification of unknown species genotypes, reinforcing the understanding of transmission and identifying new key points for effective prevention of infection [10] (Table 1).
About 44 species of Cryptosporidium and more than 120 genotypes [21,58] have been found so far, including C. hominis and C. parvum, which represent the 95% of cryptosporidiosis cases in humans. C. meleagridis, C. felis, C. canis, C. ubiquitum, C. cuniculus, C. muris, C. andersoni, C. erinacei, C. tyzzeri, C. bovis, C. suis, C. scrofarum, C. occultus, C. xiaoi, C. fayeri, C. ditrichi, C. viatorum, and other species are mostly reported in animal infections (https://www.cdc.gov/dpdx/cryptosporidiosis/index.html accessed on 20 May 2019) [11,12,16,18,59,60]. A few cases of C. meleagridis infections have also been reported in humans [61], as well as C. canis, C. felis, C. suis, C. muris, and C. andersoni in immunocompromised individuals [18,62]. C. parvum appears to be the main culprit of zoonotic infections in humans from cattle [14]. However, the absence of host specificity, notably for C. parvum, is responsible for infection in multiple hosts, such as mammals, birds, reptiles, and fish [13,63,64].
Cryptosporidium spp. is transmitted primarily by fecal-oral contamination, via the ingestion of contaminated water or food, or following contact with infected persons or animals [2], although respiratory infections have also been reported [65].
Waterborne cryptosporidiosis outbreaks are the outcome of contaminated municipal and recreational waters, including swimming pools and/or water playgrounds, even in the presence of water treatment plants, because Cryptosporidium oocyst stage can withstand disinfections, including chlorination, aiding the parasite to sustain for extended time as viable in the environment [59,66]. In some regions of Africa and Southeast Asia, cryptosporidiosis accounts for 78% of total deaths [60], and is closely related with socio-environmental factors, such as population density, which have a significant impact on the circulation of pathogens [67].
Most developing countries have no governmental system for recording incidence. Furthermore, tests for protozoa are not performed frequently and the prevalence of protozoal infections or water-related epidemics is underestimated, despite the expected high prevalence of water-related parasitic diseases in developing countries (e.g., Latin America, Asia and Africa) where people live in unsanitary conditions [30]. Indeed, there is still no international agreement on the reporting structure of cases, while the surveillance and reporting systems vary widely between the different countries and the comparison of data is not always possible [31].
The contamination of vegetables and fruits with Cryptosporidium spp. Has been extensively analyzed and documented [19,32,33,34].
In addition to aqueous and foodborne transmission, Cryptosporidium oocysts can be transmitted by the inhalation of aerosolized droplets. Since several studies have shown that respiratory aerosol droplets from infected individuals may represent one of the risk factors in the rapid transmission of Cryptosporidium oocysts [68,69,70], it is important that hospital staff use personal protective equipment, especially for the management of immunocompromised patients, to avoid the spread of Cryptosporidium oocysts through coughing and sneezing.
Indeed, oocysts have been detected in the sputum and bronchial aspirates of children with intestinal cryptosporidiosis and cough [71], supporting the hypothesis that Cryptosporidium can be transmitted by coughing, sneezing, and expectorate from infected patients.
Considering the different transmission factors and the importance of personal hygiene, Cryptosporidium is also responsible for travelers’ diarrhea, which can be considered another source of infection [63].
3. Genotyping and Spreading
The identification of Cryptosporidium at species and subspecies levels is more and more essential for understanding infection sources and gauging impact on public health in outbreak scenarios. O’ Leary et al. recently analyzed 149 C. hominis isolates from Ireland and the United Kingdom using a high resolution fusion melting (HRM) genotyping assay which characterizes and differentiates the six globally prevalent C. hominis gp60 subtypes, thus providing a profound link between epidemiological cases [8].
Several subtypes of C. hominis, C. parvum, C. meleagridis, C. tyzzeri, C. cuniculus, C. fayeri, and C. ubiquitum have been identified based on the gp60 sequence analysis, which is the most widely used genetic marker for subtyping Cryptosporidium isolates due to its high degree of genetic divergence [64].
Indeed, reported incidence rates of cryptosporidiosis in Ireland are consistently among the highest in Europe (ranging from 6.58 to 14.17 per 100,000 population, period 2006–2017) [68], with an average of 15 outbreaks annually occurring. The gp60 gene was sequenced to distinguish C. parvum and C. hominis subtypes, resulting in an incidence of C. parvum species during the spring months, while the incidence of C. hominis was confined to late summer and autumnal months. There is experimental evidence to suggest that the IIaA18G3R1 is the dominant subtype infecting cattle and humans in Ireland and Australia. In Portugal, conversely, C. parvum IIaA15G2R1 is the most frequent subtype in human and cattle [41,72].
Studies conducted in New South Wales, Australia identified C. hominis IbA10G2 and C. parvum IIaA18G3R1 as the most common subtypes and are equally responsible for the disease [73]. Recent studies suggest an important presence of C. hominis in cattle [74,75], which serve as reservoir for the protozoan. Therefore, poor hygiene and intensive farming systems are risk factors for bovine cryptosporidiosis. Epidemiological studies on the presence of Cryptosporidium spp. in Italian farms show that the prevalence does not exceed 25% for calves in central Italy [69] or in Sicily, where C. parvum was the predominant species [70], while in Sardinian farms the presence of the protozoan was significantly greater (33%) due to much more intensive farming [76].
There are several other Cryptosporidium species, including C. meleagridis, C. canis, and C. felis, that have been reported to be associated with human cases, particularly in developing countries [21]. Indeed, C. meleagridis has a broad host range and has been isolated from wild and domestic animals, such as birds, foxes, rats, farm animals, dogs, respectively, and humans. Genotyping studies reported the IIIgA31G3R1 subtype in both poultry and humans, while the IIIbA26G1R1b and IIIbA22G1R1c subtypes have been found in both children and chicken farms in China. In addition, subtypes IIIeA17G2R1, IIIeA19G2R1, IIIeA21G2R1, and IIIeA22G1R1 have been reported in Swedish and Canadian patients as well as in Asian rodents and chickens. C. canis and C. felis species infect dogs and cats, respectively, but have also been found in humans. In fact, the sequencing analysis of the gp60 gene has allowed to identify two subtypes (XIXa and XIXb) in C. felis and two subtypes (XXa1 and XXa4) in C. canis, which were also detected in humans, thus supporting zoonotic transmission [21].
Relatively little is known about the epidemiology of zoonotic species of Cryptosporidium in captive wildlife. The importance of wildlife in the dissemination of Cryptosporidium to drinking water sources and the associated human health risk are still poorly addressed. Indeed, conclusive molecular evidence linking the contamination of water supplies by wild animals as a reservoir of outbreaks of cryptosporidiosis in human populations are scarce. However, pilot studies have reported on waterborne outbreaks link to wildlife origin, such as the case of the UK outbreak caused by C. cuniculus from rabbits [77].
A variety of C. parvum subtypes, including IIdA15G1, IIdA18G1, and IIdA19G1, isolated from golden takins, lemurs, chipmunks, and hamsters, and IIaA15G2R1, IIaA19G2R1, IIaA19G3R1, IIaA19G4R1, IIaA20G3R1, IIaA20G4R1, IIaA20G3R2, and IIaA21G3R1, from deer and Eastern grey kangaroos has been reported in humans [78,79,80,81]. Other genotypes of C. ubiquitum, such as XIIb, XIIc, and XIId [82], from rodents, wild ruminants, carnivores, marsupials, and primates have also been reported sporadically in humans in the US [81].
C. erinacei subtypes, including XIIIaA21R11, XIIIaA22R9, XIIIaA21R10, XIIIa20R10, XIIIaA19R12, and XIIIaA22R11, have been identified in hedgehogs, horses, and finally in humans [83].
C. tyzzeri subtypes, such as IXaA5R2, IXaA6R1, IXaA6R2, IXaA6R3, IXbA6, and IXbA6R2, commonly reported in reptiles, have been occasionally reported in humans [81].
Two studies conducted in Australia identified C. fayeri in both immunocompetent [38] and immunosuppressed [37] women. In the first study, C. fayeri subtype IVaA9G4T1R1 was identified upon molecular screening and previously identified in Eastern grey kangaroos [84], suggesting a zoonotic transmission. In the second study, the C. fayeri subtype IVgA10G1T1R1 identified in the immunosuppressed female patient was previously reported in Western grey kangaroos [85]. Hence, the description of C. fayeri clinical infection in human, besides the above reported cases, should be considered a public health concern, overcoming the idea of “host-adapted” species, and reinforcing the idea of zoonotic transmission also for typical wildlife Cryptosporidium species.
The accuracy of genetic analysis has corroborated the understanding of the zoonotic and anthroponotic transmission potential of each species and has improved the knowledge of the epidemiology of Cryptosporidium species, leading to advanced strategies for the prevention, surveillance, and control of cryptosporidiosis in humans and other animals [86]. However, one of the limitations of the Cryptosporidium subtyping methodology is the false negative rate due to the use of species-specific markers that are able to amplify only C. parvum, C. hominis, and related species or genotypes, hence failing to detect the presence of other species potentially present in clinical samples. Furthermore, their applications are still limited due to costs [64].
4. Geographical Spreading
Only certain countries in the European Union/European Economic Area (EU/EEA) apply Cryptosporidium genotyping routinely in surveillance and outbreak investigations, which is an indication of continued single case and outbreak underreporting.
In the European Centre for Disease Prevention and Control (ECDC) report on cryptosporidiosis of 2019, in 2018, the notification rate of reported cryptosporidiosis cases was marginally higher than in the previous four years (2014–2017), with a notification rate of 4.4 per 100,000 population.
Germany, the Netherlands, Spain, and the UK accounted for 76% of all confirmed cases, with the UK alone accounting for 41%. Notification rates tended to be lower in Eastern Europe than in Western and Northern Europe. Belgium, Finland, Iceland, and the Netherlands report an increase in cryptosporidiosis cases, compared to 2017. The distribution of notified cases increased in April and peaked in September. Furthermore, the highest notification rate was observed in the 0–4 age group, with 17.7 confirmed cases per 100,000 males and 13.7 per 100,000 females. The highest notification rate in this age group was reported from Ireland (87.2 cases per 100,000 population), followed by Belgium (76.3) and the United Kingdom (32.6). Thirteen of the 23 countries for which the rates could be calculated reported less than one case per 100,000 inhabitants in this age group. In Kenya, Mali, Mozambique, Gambia, and South Asia, Cryptosporidium infection has caused a higher risk of death in toddlers aged 1–2 years with moderate-to-severe diarrhea (4%) [87].
The male-female incidence rate was nearly equivalent and varied by age group. As in previous years, notifications were higher among boys aged 0–4 years (male-female ratio 1.3/1), as well as among women of reproductive age, with male-to-female ratios of 0.6/1 and 0.5/1 in the 15–24 and 25–44 age groups, respectively. Overall, most of the reported cases (2229) among women in the EU/EEA countries were in the age group 25–44 years. Indeed, advanced genotyping techniques have enabled a fine understanding of the cryptosporidiosis epidemiology in different geographical, seasonal, and even socioeconomic contexts (https://www.cdc.gov/dpdx/cryptosporidiosis/index.html accessed on 20 May 2019).
5. Cryptosporidium Treatments
To date, despite the substantial disease burden caused by Cryptosporidium spp, treatment options remain limited and only nitazoxanide is approved for treatment, as also demonstrated by experimental studies documenting the inhibition of oocyst excretion [88,89,90].
The studies conducted on the treatment of human intestinal cryptosporidiosis have been different (Table 2). Hussein et al. used drugs such as letrazuril, a chemoprophylactic drug targeting coccidia infections, miltefosine, and clofazimine, which, however, did not show exceptional results on clinical improvement. Paromomycin, in immunocompetent children, also did not show a better effect than nitazoxanide [91].
Even high doses of albendazole, or the use of probiotics or somatostatin analogues showed a substantial reduction in the severity of diarrhea and the number of Cryptosporidim oocysts excreted in the faces. A similar result, in the reduction of oocysts, was obtained with the administration of macrolides, such as azithromycin, erythromycin, and roxithromycin [93]. Conversely, the use of acetylated spiramycin in asymptomatic individuals showed a good oocyst reduction rate [92].
Benzoxaborole inhibitors have been identified for protozoal pathogens, appearing to be parasiticidal to C. hominis and to potentially inhibit the development of intracellular C. parvum. The pyrazolopyridine was tested in neonatal calves and in mice reducing diarrhea and oocyst excretion within three days of treatment [95].
Regarding human trials, in Malawi, protocol trial team members have evaluated the safety, tolerability, pharmacokinetics, and efficacy of clofazimine on HIV-infected individuals with and without Cryptosporidium diarrhea, measuring the reduction in oocysts excretion as primary outcome, and non-cessation of clinical symptoms, which is a secondary outcome. The aim of clinical trial is also the future use of clofazimine in children of 6–18 months of age, who so far do not have a definitive treatment [94].
In general, the different therapeutic treatments take into account the clinical improvement defined as a reduction of diarrhea and complete eradication or reduction of oocysts. Several other trials are currently seeking to clarify the protective immune response against Cryptosporidium infection for effective vaccine development [95].
Currently, there is not yet a recognized vaccine and the only available pharmacological treatment approved by the FDA, EMA, and PMDA, as discussed, is nitazoxanide. Indeed, the development of new effective pharmacological treatments and vaccines is very slow, due to the lack of efficient Cryptosporidium in vitro systems and animal models able to validate in vivo the efficacy of drugs, and due to still prohibitive costs affecting trials [64,96].
These important limitations need to be overcome by access to new treatments, exploiting recent findings and filling gaps in the current clinical knowledge and management of cryptosporidiosis [97].
The use of probiotics is an alternative emerging therapeutic strategy for Cryptosporidium. Sindhu et al. reported the effect of the probiotic Lactobacillus rhamnosus GG on bowel function, immune response, and clinical outcomes in Indian children with Cryptosporidium diarrhea. According to their studies, the probiotic can prevent further damage and promote intestinal integrity in children [98]. Moreover, Pickerd et al. reported the benefits of Lactobacillus rhamnosus GG and Lactobacillus casei in the treatment of cryptosporidiosis in humans resulting in the timely clinical improvement and resolution of the infection [99]. Furthermore, since probiotics are alive microorganisms, it is believed that they can balance the gastrointestinal microbiota, thus preventing C. difficile associated diarrhea (CDAD) caused by antibiotics [100].
6. Microbiota and Cryptosporidium spp.
The human gut is a highly complex ecosystem with an extensive microbial community, and the influence of the intestinal microbiota reaches the entire host organism. Hence, the maintenance of homeostasis between the gut microbiota and the rest of the body is crucial for health [101].
The alterations of the eubiotic composition of the gut microbiota have been investigated in neonatal calves as the result of C. parvum infection, showing an increase in Fusobacterium abundance, followed by genera belonging to Bacteroidetes, Proteobacteria, Fusobacteria, and Actinobacteria phyla [102].
In human, Carey et al., analyzed 72 fecal samples from Bangladesh children to profile the microbiota during cryptosporidiosis. The authors noted that the composition of the microbiota was predictive of diarrheal symptoms both before and during the infection. Megasphaera genus was observed at high abundance in cases of subclinical Cryptosporidium infection while it was scarce or absent in cases of diarrhea. Interestingly, this observation could suggest that Megasphaera may prevent acute diarrhea during the parasite infection or at least can serve as biomarker for still unknown protective factors that could be further investigated [103].
Still, the complex interplay between parasites and the gut microbiota is poorly understood, and there are only few and often conflicting results both in humans and in animal models [104]. In the study of Bednarska et al., an investigation was conducted to determine the prevalence of intestinal pathogens, such as Cryptosporidium, Giardia, Blastocystis, Cyclospora, and microsporidia, in hospitalized patients with different immunological statuses. Cryptosporidium and Cyclospora were diagnosed as the main cause of heavy diarrhea and adult patients were positive mainly for Blastocystis and microsporidia, while children were more often for Cryptosporidium species [105].
To assess the comparative role of Cryptosporidium spp. and other enteric pathogens in animals, a pilot study was conducted in neonatal calves [106]. In this study, neonatal calves were screened for Cryptosporidium parvum, Escherichia coli K99, Rotavirus, and Coronavirus. Such mixed infections are not uncommon and are usually believed to exacerbate the clinical severity when they occur [107].
In another study, Garro et al. evaluated cryptosporidiosis of neonatal dairy calves in the presence of other enteropathogens, such as Rotavirus of group A, bovine Coronavirus, and enterotoxic E. coli. Cryptosporidium spp. was found to be the main etiological factor of diarrhea in the neonatal calf group, while Rotavirus of group A appeared to play only a secondary role in the etiology of diarrhea, suggesting that these infectious events could be independent. In contrast, mixed infection were virtually absent in older calves [108].
Charania et al. examined changes in the mouse gut microbiota following antibiotic treatment to determine how cryptosporidial infections and gut integrity were affected by alterations of the microbiome. There was a significant decrease in anaerobes and an overgrowth of Enterobacteriaceae in mice treated with cloxacillin. Moreover, there was a significant decrease in acetate, propionate, and butyrate in these same mice. Parallel to the decrease in bacterial infection, a significant increase in the severity of cryptosporidial infection and increase in gut permeability were registered. Treatment with other antibiotics significantly altered the microbiome but did not change the infection, suggesting that specific alterations in the host microbiome allow for a diversified growth of the parasite [109].
Mammeri et al. characterized the impact of C. parvum infection on the goat kid microbiome. C. parvum was orally administered to parasite-naïve goats, and infection was monitored for 26 days in fecal samples. C. parvum decreased the abundance of butyrate-producing pathways in bacteria and increased mucosal inflammation and tissue repair [110].
Clearly, the understanding of the interactions between the intestinal microbiota and enteric pathogens, such as Cryptosporidium or other parasites, is of great interest in the development of alternative treatments (e.g., probiotics, prebiotics, synbiotics) not relying on chemotherapy, and for this reason is an emerging area of research [111].
7. Study Models
Progress towards development of therapeutics for cryptosporidiosis has been hampered by lack of optimal models to support long-term cultivation of the parasite.
Several in vitro culture systems have been reported to model Cryptosporidium infection [112,113]. Two-dimensional (2D) cultures of colorectal adenocarcinoma cell lines have been most frequently used, but most of these only support short term infection (<5 days) and incomplete propagation of the parasites [114].
For this reason, new organoid technology may represent a powerful and alternative model to be used to culture and propagate Cryptosporidium and to understand the host–parasite interactions (Figure 2).
The organoid is an in vitro 3D cell cluster derived from stem cells or organ progenitors that reproduces the general structure of an organ as in vivo.
Organoids simulate the in vivo situation much better than the intestinal cell lines currently used for studying host–parasite interactions in vitro [87,115], and can support continuous parasite culture for 28 days.
Organoids may enable the identification of genes as possible therapeutic targets to develop new effective therapies. Heo et al. used small intestinal and lung organoids from healthy human donors to model the infection of C. parvum. Indeed, differentiated intact 3D intestinal organoids microinjected with sporozoites supported the full replicative cycle, resulting in newly generated oocysts that were infectious to neonatal mice. As observed in small intestinal organoids, C. parvum also forms oocysts within lung organoids because they replicate the in vivo conditions of infection [116].
8. Materials and Methods
The literature search was performed using the PubMed database in combination with the following search terms: Cryptosporidium, zoonosis, humans, animals, transmission, epidemiology, treatment, drug, nitazoxanide, vaccine. We searched PubMed original article, case reports, bulletins of CDC and ECDC, and reviews, accessing the literature published prior to 31 December 2021. Moreover, we reported on the cryptosporidiosis in humans and animals by preferring the literature focused on emerging epidemiological and drug treatment data.
9. Conclusions
Cryptosporidiosis remains a global health emergency, especially for children and severely immunocompromised patients. Currently, the lack of a government system for recording disease incidence and gold standard identification methods leads to an underestimation of the incidence of cryptosporidiosis. Cryptosporidium spp. Is the pathogen most frequently associated with food- and water-borne outbreaks and enteric infections [117]. To better control the spread of this pathogen, an educational prevention program on this parasitosis should be implemented, focusing on hygienic behavior recommendations for individuals. In fact, adopting some simple hygienic procedures, both at home and personally, such as cleaning contaminated surfaces with appropriate disinfectants, boiling tap water, or washing hands, can reduce the risk of spreading the infection. It is also important to apply efficient wastewater treatment processes that can prevent the environmental transmission of Cryptosporidium.
Although cryptosporidiosis is a worldwide cause of diarrheal diseases, no antiprotozoal agent or vaccine exists for its effective treatment or prevention. Pharmacological therapies fail to reduce the spread of oocysts to obtain clinical benefits. Considering the limited treatment options, there is therefore a need to develop a safe and effective therapy against cryptosporidiosis to improve the lives of children with acute illness and immunodeficient individuals [118]. In this regard, new disease study models based on organoid platforms have been suggested, which allow to study, in vitro, host–parasite interactions to identify new therapeutic targets.
Thus, the effectiveness of local and national surveillance and progress in drug development remain of paramount importance to contain the epidemiology of this important but often underestimated pathogen.
Author Contributions
Methodology and writing, S.P.; editing, S.P. and L.P.; conceptualization, L.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
Authors thank Federica Del Chierico for critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Putignani, L. Cryptosporidium. In Book Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Putignani, L.; Menichella, D. Global Distribution, Public Health and Clinical Impact of the Protozoan Pathogen Cryptosporidium. Interdiscip. Perspect. Infect. Dis. 2010, 2010, 753512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacciò, S.M.; Putignani, L. Epidemiology of Human Cryptosporidiosis. In Cryptosporidium: Parasite and Disease; Cacciò, S.M., Widmer, G., Eds.; Springer: Vienna, Austria, 2014; pp. 43–79. ISBN 978-3-7091-1561-9. [Google Scholar]
- Checkley, W.; White, A.C.; Jaganath, D.; Arrowood, M.J.; Chalmers, R.M.; Chen, X.-M.; Fayer, R.; Griffiths, J.K.; Guerrant, R.L.; Hedstrom, L.; et al. A Review of the Global Burden, Novel Diagnostics, Therapeutics, and Vaccine Targets for Cryptosporidium. Lancet Infect. Dis. 2015, 15, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, J.F.; Ayoub, A.; Ayers, M.S. Treatment of Diarrhea Caused by Cryptosporidium Parvum: A Prospective Randomized, Double-Blind, Placebo-Controlled Study of Nitazoxanide. J. Infect. Dis. 2001, 184, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amadi, B.; Mwiya, M.; Musuku, J.; Watuka, A.; Sianongo, S.; Ayoub, A.; Kelly, P. Effect of Nitazoxanide on Morbidity and Mortality in Zambian Children with Cryptosporidiosis: A Randomised Controlled Trial. Lancet Lond. Engl. 2002, 360, 1375–1380. [Google Scholar] [CrossRef]
- Bones, A.J.; Jossé, L.; More, C.; Miller, C.N.; Michaelis, M.; Tsaousis, A.D. Past and Future Trends of Cryptosporidium in Vitro Research. Exp. Parasitol. 2019, 196, 28–37. [Google Scholar] [CrossRef]
- O’ Leary, J.K.; Blake, L.; Corcoran, G.D.; Sleator, R.D.; Lucey, B. A Novel Genotyping Method for Cryptosporidium Hominis. Exp. Parasitol. 2021, 225, 108113. [Google Scholar] [CrossRef]
- King, P.; Tyler, K.M.; Hunter, P.R. Anthroponotic Transmission of Cryptosporidium Parvum Predominates in Countries with Poorer Sanitation: A Systematic Review and Meta-Analysis. Parasit. Vectors 2019, 12, 16. [Google Scholar] [CrossRef]
- Widmer, G.; Carmena, D.; Kváč, M.; Chalmers, R.M.; Kissinger, J.C.; Xiao, L.; Sateriale, A.; Striepen, B.; Laurent, F.; Lacroix-Lamandé, S.; et al. Update on Cryptosporidium Spp.: Highlights from the Seventh International Giardia and Cryptosporidium Conference. Parasite Paris Fr. 2020, 27, 14. [Google Scholar] [CrossRef] [Green Version]
- Koehler, A.V.; Whipp, M.J.; Haydon, S.R.; Gasser, R.B. Cryptosporidium Cuniculus--New Records in Human and Kangaroo in Australia. Parasit. Vectors 2014, 7, 492. [Google Scholar] [CrossRef]
- Feng, Y.; Ryan, U.M.; Xiao, L. Genetic Diversity and Population Structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef]
- Fayer, R. Taxonomy and Species Delimitation in Cryptosporidium. Exp. Parasitol. 2010, 124, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidium Species in Humans and Animals: Current Understanding and Research Needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyzzer, E.E. An Extracellular Coccidium, Cryptosporidium Muris (Gen. Et Sp. Nov.), of the Gastric Glands of the Common Mouse. J. Med. Res. 1910, 23, 487–510.3. [Google Scholar] [PubMed]
- Vetterling, J.M.; Jervis, H.R.; Merrill, T.G.; Sprinz, H. Cryptosporidium Wrairi Sp. n. from the Guinea Pig Cavia Porcellus, with an Emendation of the Genus. J. Protozool. 1971, 18, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Trout, J.M.; Xiao, L.; Morgan, U.M.; Lal, A.A.; Dubey, J.P. CRYPTOSPORIDIUM CANIS N. SP. FROM DOMESTIC DOGS. J. Parasitol. 2001, 87, 1415–1422. [Google Scholar] [CrossRef]
- Fayer, R.; Santín, M.; Macarisin, D. Cryptosporidium Ubiquitum n. Sp. in Animals and Humans. Vet. Parasitol. 2010, 172, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Santín, M. Cryptosporidium Xiaoi n. Sp. (Apicomplexa: Cryptosporidiidae) in Sheep (Ovis Aries). Vet. Parasitol. 2009, 164, 192–200. [Google Scholar] [CrossRef]
- Li, J.; Guo, Y.; Roellig, D.M.; Li, N.; Feng, Y.; Xiao, L. Cryptosporidium Felis Differs from Other Cryptosporidium Spp. in Codon Usage. Microb. Genom. 2021, 7, 000711. [Google Scholar] [CrossRef]
- Ryan, U.; Zahedi, A.; Feng, Y.; Xiao, L. An Update on Zoonotic Cryptosporidium Species and Genotypes in Humans. Animals 2021, 11, 3307. [Google Scholar] [CrossRef]
- Levine, N.D. Taxonomy and Review of the Coccidian Genus Cryptosporidium (Protozoa, Apicomplexa). J. Protozool. 1984, 31, 94–98. [Google Scholar] [CrossRef]
- Alvarez-Pellitero, P.; Sitjà-Bobadilla, A. Cryptosporidium Molnari n. Sp. (Apicomplexa: Cryptosporidiidae) Infecting Two Marine Fish Species, Sparus aurata L. and Dicentrarchus labrax L. Int. J. Parasitol. 2002, 32, 1007–1021. [Google Scholar] [CrossRef]
- Alvarez-Pellitero, P.; Quiroga, M.; Sitjà-Bobadilla, A.; Redondo, M.; Palenzuela, O.; Padrós, F.; Vázquez, S.; Nieto, J. Cryptosporidium Scophthalmi n. Sp. (Apicomplexa: Cryptosporidiidae) from Cultured Turbot Scophthalmus Maximus. Light and Electron Microscope Description and Histopathological Study. Dis. Aquat. Organ. 2004, 62, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Kváč, M.; Kestřánová, M.; Pinková, M.; Květoňová, D.; Kalinová, J.; Wagnerová, P.; Kotková, M.; Vítovec, J.; Ditrich, O.; McEvoy, J.; et al. Cryptosporidium Scrofarum n. Sp. (Apicomplexa: Cryptosporidiidae) in Domestic Pigs (Sus Scrofa). Vet. Parasitol. 2013, 191, 218–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kváč, M.; Květoňová, D.; Sak, B.; Ditrich, O. Cryptosporidium Pig Genotype II in Immunocompetent Man. Emerg. Infect. Dis. 2009, 15, 982–983. [Google Scholar] [CrossRef] [PubMed]
- Current, W.L.; Upton, S.J.; Haynes, T.B. The Life Cycle of Cryptosporidium Baileyi n. Sp. (Apicomplexa, Cryptosporidiidae) Infecting Chickens. J. Protozool. 1986, 33, 289–296. [Google Scholar] [CrossRef]
- Ditrich, O.; Palkovič, L.; Štěrba, J.; Prokopič, J.; Loudová, J.; Giboda, M. The First Finding OfCryptosporidium Baileyi in Man. Parasitol. Res. 1991, 77, 44–47. [Google Scholar] [CrossRef]
- da Silva, D.C.; Homem, C.G.; Nakamura, A.A.; Teixeira, W.F.P.; Perri, S.H.V.; Meireles, M.V. Physical, Epidemiological, and Molecular Evaluation of Infection by Cryptosporidium Galli in Passeriformes. Parasitol. Res. 2010, 107, 271–277. [Google Scholar] [CrossRef]
- Jirků, M.; Valigurová, A.; Koudela, B.; Krízek, J.; Modrý, D.; Slapeta, J. New Species of Cryptosporidium Tyzzer, 1907 (Apicomplexa) from Amphibian Host: Morphology, Biology and Phylogeny. Folia Parasitol. 2008, 55, 81–94. [Google Scholar] [CrossRef] [Green Version]
- da Silva, D.C.; Paiva, P.R.S.O.; Nakamura, A.A.; Homem, C.G.; de Souza, M.S.; Grego, K.F.; Meireles, M.V. The Detection of Cryptosporidium Serpentis in Snake Fecal Samples by Real-Time PCR. Vet. Parasitol. 2014, 204, 134–138. [Google Scholar] [CrossRef]
- Pedraza-Díaz, S.; Ortega-Mora, L.M.; Carrión, B.A.; Navarro, V.; Gómez-Bautista, M. Molecular Characterisation of Cryptosporidium Isolates from Pet Reptiles. Vet. Parasitol. 2009, 160, 204–210. [Google Scholar] [CrossRef]
- Fayer, R.; Santín, M.; Trout, J.M. Cryptosporidium Ryanae n. Sp. (Apicomplexa: Cryptosporidiidae) in Cattle (Bos Taurus). Vet. Parasitol. 2008, 156, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Santín, M.; Xiao, L. Cryptosporidium Bovis n. Sp. (Apicomplexa: Cryptosporidiidae) in Cattle (Bos Taurus). J. Parasitol. 2005, 91, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Higuera, A.; Villamizar, X.; Herrera, G.; Giraldo, J.C.; Vasquez-A, L.R.; Urbano, P.; Villalobos, O.; Tovar, C.; Ramírez, J.D. Molecular Detection and Genotyping of Intestinal Protozoa from Different Biogeographical Regions of Colombia. PeerJ 2020, 8, e8554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, U.M.; Power, M.; Xiao, L. Cryptosporidium Fayeri n. Sp. (Apicomplexa: Cryptosporidiidae) from the Red Kangaroo (Macropus Rufus). J. Eukaryot. Microbiol. 2008, 55, 22–26. [Google Scholar] [CrossRef]
- Braima, K.; Zahedi, A.; Oskam, C.; Austen, J.; Egan, S.; Reid, S.; Ryan, U. Zoonotic Infection by Cryptosporidium Fayeri IVgA10G1T1R1 in a Western Australian Human. Zoonoses Public Health 2021, 68, 358–360. [Google Scholar] [CrossRef]
- Waldron, L.S.; Cheung-Kwok-Sang, C.; Power, M.L. Wildlife-Associated Cryptosporidium Fayeri in Human, Australia. Emerg. Infect. Dis. 2010, 16, 2006–2007. [Google Scholar] [CrossRef]
- Power, M.L.; Ryan, U.M. A New Species of Cryptosporidium (Apicomplexa: Cryptosporidiidae) from Eastern Grey Kangaroos (Macropus Giganteus). J. Parasitol. 2008, 94, 1114–1117. [Google Scholar] [CrossRef]
- Ren, X.; Zhao, J.; Zhang, L.; Ning, C.; Jian, F.; Wang, R.; Lv, C.; Wang, Q.; Arrowood, M.J.; Xiao, L. Cryptosporidium Tyzzeri n. Sp. (Apicomplexa: Cryptosporidiidae) in Domestic Mice (Mus Musculus). Exp. Parasitol. 2012, 130, 274–281. [Google Scholar] [CrossRef]
- Garcia-R, J.C.; Pita, A.B.; Velathanthiri, N.; French, N.P.; Hayman, D.T.S. Species and Genotypes Causing Human Cryptosporidiosis in New Zealand. Parasitol. Res. 2020, 119, 2317–2326. [Google Scholar] [CrossRef]
- Elwin, K.; Hadfield, S.J.; Robinson, G.; Crouch, N.D.; Chalmers, R.M. Cryptosporidium Viatorum n. Sp. (Apicomplexa: Cryptosporidiidae) among Travellers Returning to Great Britain from the Indian Subcontinent, 2007–2011. Int. J. Parasitol. 2012, 42, 675–682. [Google Scholar] [CrossRef]
- Ong, C.S.L.; Eisler, D.L.; Alikhani, A.; Fung, V.W.K.; Tomblin, J.; Bowie, W.R.; Isaac-Renton, J.L. Novel Cryptosporidium Genotypes in Sporadic Cryptosporidiosis Cases: First Report of Human Infections with a Cervine Genotype. Emerg. Infect. Dis. 2002, 8, 263–268. [Google Scholar] [CrossRef]
- Holubová, N.; Zikmundová, V.; Limpouchová, Z.; Sak, B.; Konečný, R.; Hlásková, L.; Rajský, D.; Kopacz, Z.; McEvoy, J.; Kváč, M. Cryptosporidium Proventriculi Sp. n. (Apicomplexa: Cryptosporidiidae) in Psittaciformes Birds. Eur. J. Protistol. 2019, 69, 70–87. [Google Scholar] [CrossRef]
- Holubová, N.; Tůmová, L.; Sak, B.; Hejzlarová, A.; Konečný, R.; McEvoy, J.; Kváč, M. Description of Cryptosporidium Ornithophilus n. Sp. (Apicomplexa: Cryptosporidiidae) in Farmed Ostriches. Parasit. Vectors 2020, 13, 340. [Google Scholar] [CrossRef] [PubMed]
- Ježková, J.; Prediger, J.; Holubová, N.; Sak, B.; Konečný, R.; Feng, Y.; Xiao, L.; Rost, M.; McEvoy, J.; Kváč, M. Cryptosporidium Ratti n. Sp. (Apicomplexa: Cryptosporidiidae) and Genetic Diversity of Cryptosporidium Spp. in Brown Rats (Rattus Norvegicus) in the Czech Republic. Parasitology 2021, 148, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Kváč, M.; Hofmannová, L.; Hlásková, L.; Květoňová, D.; Vítovec, J.; McEvoy, J.; Sak, B. Cryptosporidium Erinacei n. Sp. (Apicomplexa: Cryptosporidiidae) in Hedgehogs. Vet. Parasitol. 2014, 201, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Prediger, J.; Ježková, J.; Holubová, N.; Sak, B.; Konečný, R.; Rost, M.; McEvoy, J.; Rajský, D.; Kváč, M. Cryptosporidium Sciurinum n. Sp. (Apicomplexa: Cryptosporidiidae) in Eurasian Red Squirrels (Sciurus Vulgaris). Microorganisms 2021, 9, 2050. [Google Scholar] [CrossRef]
- Ježková, J.; Limpouchová, Z.; Prediger, J.; Holubová, N.; Sak, B.; Konečný, R.; Květoňová, D.; Hlásková, L.; Rost, M.; McEvoy, J.; et al. Cryptosporidium Myocastoris n. Sp. (Apicomplexa: Cryptosporidiidae), the Species Adapted to the Nutria (Myocastor Coypus). Microorganisms 2021, 9, 813. [Google Scholar] [CrossRef]
- Jezkova, J.; Horcickova, M.; Hlaskova, L.; Sak, B.; Kvetonova, D.; Novak, J.; Hofmannova, L.; McEvoy, J.; Kvac, M. Cryptosporidium Testudinis Sp. n., Cryptosporidium Ducismarci Traversa, 2010 and Cryptosporidium Tortoise Genotype III (Apicomplexa: Cryptosporidiidae) in Tortoises. Folia Parasitol. 2016, 63, 35. [Google Scholar] [CrossRef] [Green Version]
- Holubová, N.; Sak, B.; Horčičková, M.; Hlásková, L.; Květoňová, D.; Menchaca, S.; McEvoy, J.; Kváč, M. Cryptosporidium Avium n. Sp. (Apicomplexa: Cryptosporidiidae) in Birds. Parasitol. Res. 2016, 115, 2243–2251. [Google Scholar] [CrossRef]
- Horčičková, M.; Čondlová, Š.; Holubová, N.; Sak, B.; Květoňová, D.; Hlásková, L.; Konečný, R.; Sedláček, F.; Clark, M.; Giddings, C.; et al. Diversity of Cryptosporidium in Common Voles and Description of Cryptosporidium Alticolis Sp. n. and Cryptosporidium Microti Sp. n. (Apicomplexa: Cryptosporidiidae). Parasitology 2019, 146, 220–233. [Google Scholar] [CrossRef]
- Zahedi, A.; Bolland, S.J.; Oskam, C.L.; Ryan, U. Cryptosporidium Abrahamseni n. Sp. (Apicomplexa: Cryptosporidiiae) from Red-Eye Tetra (Moenkhausia Sanctaefilomenae). Exp. Parasitol. 2021, 223, 108089. [Google Scholar] [CrossRef] [PubMed]
- Bolland, S.J.; Zahedi, A.; Oskam, C.; Murphy, B.; Ryan, U. Cryptosporidium Bollandi n. Sp. (Apicomplexa: Cryptosporidiiae) from Angelfish (Pterophyllum Scalare) and Oscar Fish (Astronotus Ocellatus). Exp. Parasitol. 2020, 217, 107956. [Google Scholar] [CrossRef] [PubMed]
- Čondlová, Š.; Horčičková, M.; Sak, B.; Květoňová, D.; Hlásková, L.; Konečný, R.; Stanko, M.; McEvoy, J.; Kváč, M. Cryptosporidium Apodemi Sp. n. and Cryptosporidium Ditrichi Sp. n. (Apicomplexa: Cryptosporidiidae) in Apodemus spp. Eur. J. Protistol. 2018, 63, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Beser, J.; Bujila, I.; Wittesjö, B.; Lebbad, M. From Mice to Men: Three Cases of Human Infection with Cryptosporidium Ditrichi. Infect. Genet. Evol. 2020, 78, 104120. [Google Scholar] [CrossRef] [PubMed]
- Kváč, M.; Havrdová, N.; Hlásková, L.; Daňková, T.; Kanděra, J.; Ježková, J.; Vítovec, J.; Sak, B.; Ortega, Y.; Xiao, L.; et al. Cryptosporidium Proliferans n. Sp. (Apicomplexa: Cryptosporidiidae): Molecular and Biological Evidence of Cryptic Species within Gastric Cryptosporidium of Mammals. PLoS ONE 2016, 11, e0147090. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Karim, M.R.; Zhang, L. Detection of Human Intestinal Protozoan Parasites in Vegetables and Fruits: A Review. Parasit. Vectors 2020, 13, 380. [Google Scholar] [CrossRef]
- Hlavsa, M.C.; Aluko, S.K.; Miller, A.D.; Person, J.; Gerdes, M.E.; Lee, S.; Laco, J.P.; Hannapel, E.J.; Hill, V.R. Outbreaks Associated with Treated Recreational Water—United States, 2015–2019. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 733–738. [Google Scholar] [CrossRef]
- Boschi-Pinto, C. Estimating Child Mortality Due to Diarrhoea in Developing Countries. Bull. World Health Organ. 2008, 86, 710–717. [Google Scholar] [CrossRef]
- Xiao, L.; Ryan, U.M. Cryptosporidiosis: An Update in Molecular Epidemiology. Curr. Opin. Infect. Dis. 2004, 17, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Pumipuntu, N.; Piratae, S. Cryptosporidiosis: A Zoonotic Disease Concern. Vet. World 2018, 11, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Vanathy, K.; Parija, S.C.; Mandal, J.; Hamide, A.; Krishnamurthy, S. Cryptosporidiosis: A Mini Review. Trop. Parasitol. 2017, 7, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Shaik, J.S.; Grigg, M.E. Genomics and Molecular Epidemiology of Cryptosporidium Species. Acta Trop. 2018, 184, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sponseller, J.K.; Griffiths, J.K.; Tzipori, S. The Evolution of Respiratory Cryptosporidiosis: Evidence for Transmission by Inhalation. Clin. Microbiol. Rev. 2014, 27, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogan, J.E. Disinfection Techniques for Cryptosporidium. J. Dairy Vet. Sci. 2018, 7, 001–003. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; LeJeune, J.; Alsdorf, D.; Lu, B.; Shum, C.K.; Liang, S. Global Distribution of Outbreaks of Water-Associated Infectious Diseases. PLoS Negl. Trop. Dis. 2012, 6, e1483. [Google Scholar] [CrossRef] [Green Version]
- O’ Leary, J.K.; Blake, L.; Corcoran, G.D.; Sleator, R.D.; Lucey, B. Increased Diversity and Novel Subtypes among Clinical Cryptosporidium Parvum and Cryptosporidium Hominis Isolates in Southern Ireland. Exp. Parasitol. 2020, 218, 107967. [Google Scholar] [CrossRef]
- Duranti, A.; Cacciò, S.M.; Pozio, E.; Di Egidio, A.; De Curtis, M.; Battisti, A.; Scaramozzino, P. Risk Factors Associated with Cryptosporidium Parvum Infection in Cattle. Zoonoses Public Health 2009, 56, 176–182. [Google Scholar] [CrossRef]
- Di Piazza, F.; Di Benedetto, M.A.; Maida, C.M.; Glorioso, S.; Adamo, G.; Mazzola, T.; Firenze, A. A Study on Occupational Exposure of Sicilian Farmers to Giardia and Cryptosporidium. J. Prev. Med. Hyg. 2013, 54, 212–217. [Google Scholar]
- Mor, S.M.; Tumwine, J.K.; Ndeezi, G.; Srinivasan, M.G.; Kaddu-Mulindwa, D.H.; Tzipori, S.; Griffiths, J.K. Respiratory Cryptosporidiosis in HIV-Seronegative Children in Uganda: Potential for Respiratory Transmission. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2010, 50, 1366–1372. [Google Scholar] [CrossRef] [Green Version]
- Waldron, L.S.; Dimeski, B.; Beggs, P.J.; Ferrari, B.C.; Power, M.L. Molecular Epidemiology, Spatiotemporal Analysis, and Ecology of Sporadic Human Cryptosporidiosis in Australia. Appl. Environ. Microbiol. 2011, 77, 7757–7765. [Google Scholar] [CrossRef] [Green Version]
- Waldron, L.S.; Ferrari, B.C.; Power, M.L. Glycoprotein 60 Diversity in C. Hominis and C. Parvum Causing Human Cryptosporidiosis in NSW, Australia. Exp. Parasitol. 2009, 122, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, A.; Monis, P.; Aucote, S.; King, B.; Paparini, A.; Jian, F.; Yang, R.; Oskam, C.; Ball, A.; Robertson, I.; et al. Zoonotic Cryptosporidium Species in Animals Inhabiting Sydney Water Catchments. PLoS ONE 2016, 11, e0168169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razakandrainibe, R.; Diawara, E.H.I.; Costa, D.; Le Goff, L.; Lemeteil, D.; Ballet, J.J.; Gargala, G.; Favennec, L. Common Occurrence of Cryptosporidium Hominis in Asymptomatic and Symptomatic Calves in France. PLoS Negl. Trop. Dis. 2018, 12, e0006355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varcasia, A.; Capelli, G.; Ruiu, A.; Ladu, M.; Scala, A.; Bjorkman, C. Prevalence of Neospora Caninum Infection in Sardinian Dairy Farms (Italy) Detected by Iscom ELISA on Tank Bulk Milk. Parasitol. Res. 2006, 98, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, R.M.; Robinson, G.; Elwin, K.; Hadfield, S.J.; Xiao, L.; Ryan, U.; Modha, D.; Mallaghan, C. Cryptosporidium Sp. Rabbit Genotype, a Newly Identified Human Pathogen. Emerg. Infect. Dis. 2009, 15, 829–830. [Google Scholar] [CrossRef]
- Lv, C.; Zhang, L.; Wang, R.; Jian, F.; Zhang, S.; Ning, C.; Wang, H.; Feng, C.; Wang, X.; Ren, X.; et al. Cryptosporidium Spp. in Wild, Laboratory, and Pet Rodents in China: Prevalence and Molecular Characterization. Appl. Environ. Microbiol. 2009, 75, 7692–7699. [Google Scholar] [CrossRef] [Green Version]
- Bodager, J.R.; Parsons, M.B.; Wright, P.C.; Rasambainarivo, F.; Roellig, D.; Xiao, L.; Gillespie, T.R. Complex Epidemiology and Zoonotic Potential for Cryptosporidium Suis in Rural Madagascar. Vet. Parasitol. 2015, 207, 140–143. [Google Scholar] [CrossRef]
- Montecino-Latorre, D.; Li, X.; Xiao, C.; Atwill, E.R. Elevation and Vegetation Determine Cryptosporidium Oocyst Shedding by Yellow-Bellied Marmots (Marmota Flaviventris) in the Sierra Nevada Mountains. Int. J. Parasitol. Parasites Wildl. 2015, 4, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, A.; Paparini, A.; Jian, F.; Robertson, I.; Ryan, U. Public Health Significance of Zoonotic Cryptosporidium Species in Wildlife: Critical Insights into Better Drinking Water Management. Int. J. Parasitol. Parasites Wildl. 2016, 5, 88–109. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Xiao, L.; Alderisio, K.; Elwin, K.; Cebelinski, E.; Chalmers, R.; Santin, M.; Fayer, R.; Kvac, M.; Ryan, U.; et al. Subtyping Cryptosporidium Ubiquitum, a Zoonotic Pathogen Emerging in Humans. Emerg. Infect. Dis. 2014, 20, 217–224. [Google Scholar] [CrossRef]
- Dyachenko, V.; Kuhnert, Y.; Schmaeschke, R.; Etzold, M.; Pantchev, N.; Daugschies, A. Occurrence and Molecular Characterization of Cryptosporidium Spp. Genotypes in European Hedgehogs (Erinaceus europaeus L.) in Germany. Parasitology 2010, 137, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Power, M.L.; Cheung-Kwok-Sang, C.; Slade, M.; Williamson, S. Cryptosporidium Fayeri: Diversity within the GP60 Locus of Isolates from Different Marsupial Hosts. Exp. Parasitol. 2009, 121, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, A.; Monis, P.; Gofton, A.W.; Oskam, C.L.; Ball, A.; Bath, A.; Bartkow, M.; Robertson, I.; Ryan, U. Cryptosporidium Species and Subtypes in Animals Inhabiting Drinking Water Catchments in Three States across Australia. Water Res. 2018, 134, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Jex, A.R.; Smith, H.V.; Monis, P.T.; Campbell, B.E.; Gasser, R.B. Cryptosporidium—Biotechnological Advances in the Detection, Diagnosis and Analysis of Genetic Variation. Biotechnol. Adv. 2008, 26, 304–317. [Google Scholar] [CrossRef]
- Klotz, C.; Aebischer, T.; Seeber, F. Stem Cell-Derived Cell Cultures and Organoids for Protozoan Parasite Propagation and Studying Host-Parasite Interaction. Int. J. Med. Microbiol. IJMM 2012, 302, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Guo, F.; Zhu, G. Cryptosporidium Lactate Dehydrogenase Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Target for Developing Therapeutics. PLoS Pathog. 2015, 11, e1005250. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P. Treatment and Prevention of Cryptosporidiosis: What Options Are There for a Country like Zambia? Parasitology 2011, 138, 1488–1491. [Google Scholar] [CrossRef]
- Diawara, E.H.; François, A.; Stachulski, A.V.; Razakandrainibe, R.; Costa, D.; Favennec, L.; Rossignol, J.-F.; Gargala, G. Systemic Efficacy on Cryptosporidium Parvum Infection of Aminoxanide (RM-5061), a New Amino-Acid Ester Thiazolide Prodrug of Tizoxanide. Parasitology 2021, 148, 975–984. [Google Scholar] [CrossRef]
- Hussien, S.M.M.; Abdella, O.H.; Abu-Hashim, A.H.; Aboshiesha, G.A.; Taha, M.A.A.; El-Shemy, A.S.; El-Bader, M.M. Comparative Study between the Effect of Nitazoxanide and Paromomycine in Treatment of Cryptosporidiosis in Hospitalized Children. J. Egypt. Soc. Parasitol. 2013, 43, 463–470. [Google Scholar]
- Huang, M.-Z.; Li, J.; Guan, L.; Li, D.-Q.; Nie, X.-M.; Gui, R.; Chen, X. Therapeutic Effects of Acetylspiramycin and Garlicin on Cryptosporidiosis among Drug Users. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Diptyanusa, A.; Sari, I.P. Treatment of Human Intestinal Cryptosporidiosis: A Review of Published Clinical Trials. Int. J. Parasitol. Drugs Drug Resist. 2021, 17, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Nachipo, P.; Hermann, D.; Quinnan, G.; Gordon, M.A.; Van Voorhis, W.C.; Iroh Tam, P.-Y. Evaluating the Safety, Tolerability, Pharmacokinetics and Efficacy of Clofazimine in Cryptosporidiosis (CRYPTOFAZ): Study Protocol for a Randomized Controlled Trial. Trials 2018, 19, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, S.U.; Mi, R.; Zhou, S.; Gong, H.; Ullah, M.; Huang, Y.; Han, X.; Chen, Z. Advances in Therapeutic and Vaccine Targets for Cryptosporidium: Challenges and Possible Mitigation Strategies. Acta Trop. 2021, 226, 106273. [Google Scholar] [CrossRef] [PubMed]
- Vinayak, S.; Pawlowic, M.C.; Sateriale, A.; Brooks, C.F.; Studstill, C.J.; Bar-Peled, Y.; Cipriano, M.J.; Striepen, B. Genetic Modification of the Diarrhoeal Pathogen Cryptosporidium Parvum. Nature 2015, 523, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Ashigbie, P.G.; Shepherd, S.; Steiner, K.L.; Amadi, B.; Aziz, N.; Manjunatha, U.H.; Spector, J.M.; Diagana, T.T.; Kelly, P. Use-Case Scenarios for an Anti-Cryptosporidium Therapeutic. PLoS Negl. Trop. Dis. 2021, 15, e0009057. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, K.N.C.; Sowmyanarayanan, T.V.; Paul, A.; Babji, S.; Ajjampur, S.S.R.; Priyadarshini, S.; Sarkar, R.; Balasubramanian, K.A.; Wanke, C.A.; Ward, H.D.; et al. Immune Response and Intestinal Permeability in Children with Acute Gastroenteritis Treated with Lactobacillus Rhamnosus GG: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2014, 58, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Pickerd, N.; Tuthill, D. Resolution of Cryptosporidiosis with Probiotic Treatment. Postgrad. Med. J. 2004, 80, 112–113. [Google Scholar] [CrossRef] [Green Version]
- Sebastián Domingo, J.J. Review of the Role of Probiotics in Gastrointestinal Diseases in Adults. Gastroenterol. Hepatol. 2017, 40, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Partida-Rodríguez, O.; Serrano-Vázquez, A.; Nieves-Ramírez, M.E.; Moran, P.; Rojas, L.; Portillo, T.; González, E.; Hernández, E.; Finlay, B.B.; Ximenez, C. Human Intestinal Microbiota: Interaction Between Parasites and the Host Immune Response. Arch. Med. Res. 2017, 48, 690–700. [Google Scholar] [CrossRef]
- Ichikawa-Seki, M.; Motooka, D.; Kinami, A.; Murakoshi, F.; Takahashi, Y.; Aita, J.; Hayashi, K.; Tashibu, A.; Nakamura, S.; Iida, T.; et al. Specific Increase of Fusobacterium in the Faecal Microbiota of Neonatal Calves Infected with Cryptosporidium Parvum. Sci. Rep. 2019, 9, 12517. [Google Scholar] [CrossRef] [Green Version]
- Carey, M.A.; Medlock, G.L.; Alam, M.; Kabir, M.; Uddin, M.J.; Nayak, U.; Papin, J.; Faruque, A.S.G.; Haque, R.; Petri, W.A.; et al. Megasphaera in the Stool Microbiota Is Negatively Associated With Diarrheal Cryptosporidiosis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 73, e1242–e1251. [Google Scholar] [CrossRef] [PubMed]
- Brosschot, T.P.; Reynolds, L.A. The Impact of a Helminth-Modified Microbiome on Host Immunity. Mucosal Immunol. 2018, 11, 1039–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bednarska, M.; Jankowska, I.; Pawelas, A.; Piwczyńska, K.; Bajer, A.; Wolska-Kuśnierz, B.; Wielopolska, M.; Welc-Falęciak, R. Prevalence of Cryptosporidium, Blastocystis, and Other Opportunistic Infections in Patients with Primary and Acquired Immunodeficiency. Parasitol. Res. 2018, 117, 2869–2879. [Google Scholar] [CrossRef] [Green Version]
- Mohteshamuddin, K.; Hamdan, L.; AlKaabi, A.B.; Barigye, R. Cryptosporidium Parvum and Other Enteric Pathogens in Scouring Neonatal Dairy Calves from the Al Ain Region, United Arab Emirates. Vet. Parasitol. Reg. Stud. Rep. 2020, 21, 100435. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Yoon, K.-J. An Overview of Calf Diarrhea—Infectious Etiology, Diagnosis, and Intervention. J. Vet. Sci. 2014, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Garro, C.J.; Morici, G.E.; Tomazic, M.L.; Vilte, D.; Encinas, M.; Vega, C.; Bok, M.; Parreño, V.; Schnittger, L. Occurrence of Cryptosporidium and Other Enteropathogens and Their Association with Diarrhea in Dairy Calves of Buenos Aires Province, Argentina. Vet. Parasitol. Reg. Stud. Rep. 2021, 24, 100567. [Google Scholar] [CrossRef]
- Charania, R.; Wade, B.E.; McNair, N.N.; Mead, J.R. Changes in the Microbiome of Cryptosporidium-Infected Mice Correlate to Differences in Susceptibility and Infection Levels. Microorganisms 2020, 8, 879. [Google Scholar] [CrossRef]
- Mammeri, M.; Obregón, D.A.; Chevillot, A.; Polack, B.; Julien, C.; Pollet, T.; Cabezas-Cruz, A.; Adjou, K.T. Cryptosporidium Parvum Infection Depletes Butyrate Producer Bacteria in Goat Kid Microbiome. Front. Microbiol. 2020, 11, 548737. [Google Scholar] [CrossRef]
- Pane, S.; Ristori, M.V.; Gardini, S.; Russo, A.; Del Chierico, F.; Putignani, L. Clinical Parasitology and Parasitome Maps as Old and New Tools to Improve Clinical Microbiomics. Pathogens 2021, 10, 1550. [Google Scholar] [CrossRef]
- Karanis, P. The Truth about in Vitro Culture of Cryptosporidium Species. Parasitology 2018, 145, 855–864. [Google Scholar] [CrossRef]
- Karanis, P.; Aldeyarbi, H.M. Evolution of Cryptosporidium in Vitro Culture. Int. J. Parasitol. 2011, 41, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Bhalchandra, S.; Lamisere, H.; Ward, H. Intestinal Organoid/Enteroid-Based Models for Cryptosporidium. Curr. Opin. Microbiol. 2020, 58, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Chieppa, M.; Rescigno, M.; Huang, A.Y.C.; Germain, R.N. Dynamic Imaging of Dendritic Cell Extension into the Small Bowel Lumen in Response to Epithelial Cell TLR Engagement. J. Exp. Med. 2006, 203, 2841–2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, I.; Dutta, D.; Schaefer, D.A.; Iakobachvili, N.; Artegiani, B.; Sachs, N.; Boonekamp, K.E.; Bowden, G.; Hendrickx, A.P.A.; Willems, R.J.L.; et al. Modelling Cryptosporidium Infection in Human Small Intestinal and Lung Organoids. Nat. Microbiol. 2018, 3, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Luka, G.; Samiei, E.; Tasnim, N.; Dalili, A.; Najjaran, H.; Hoorfar, M. Comprehensive Review of Conventional and State-of-the-Art Detection Methods of Cryptosporidium. J. Hazard. Mater. 2022, 421, 126714. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, U.H.; Chao, A.T.; Leong, F.J.; Diagana, T.T. Cryptosporidiosis Drug Discovery: Opportunities and Challenges. ACS Infect. Dis. 2016, 2, 530–537. [Google Scholar] [CrossRef]
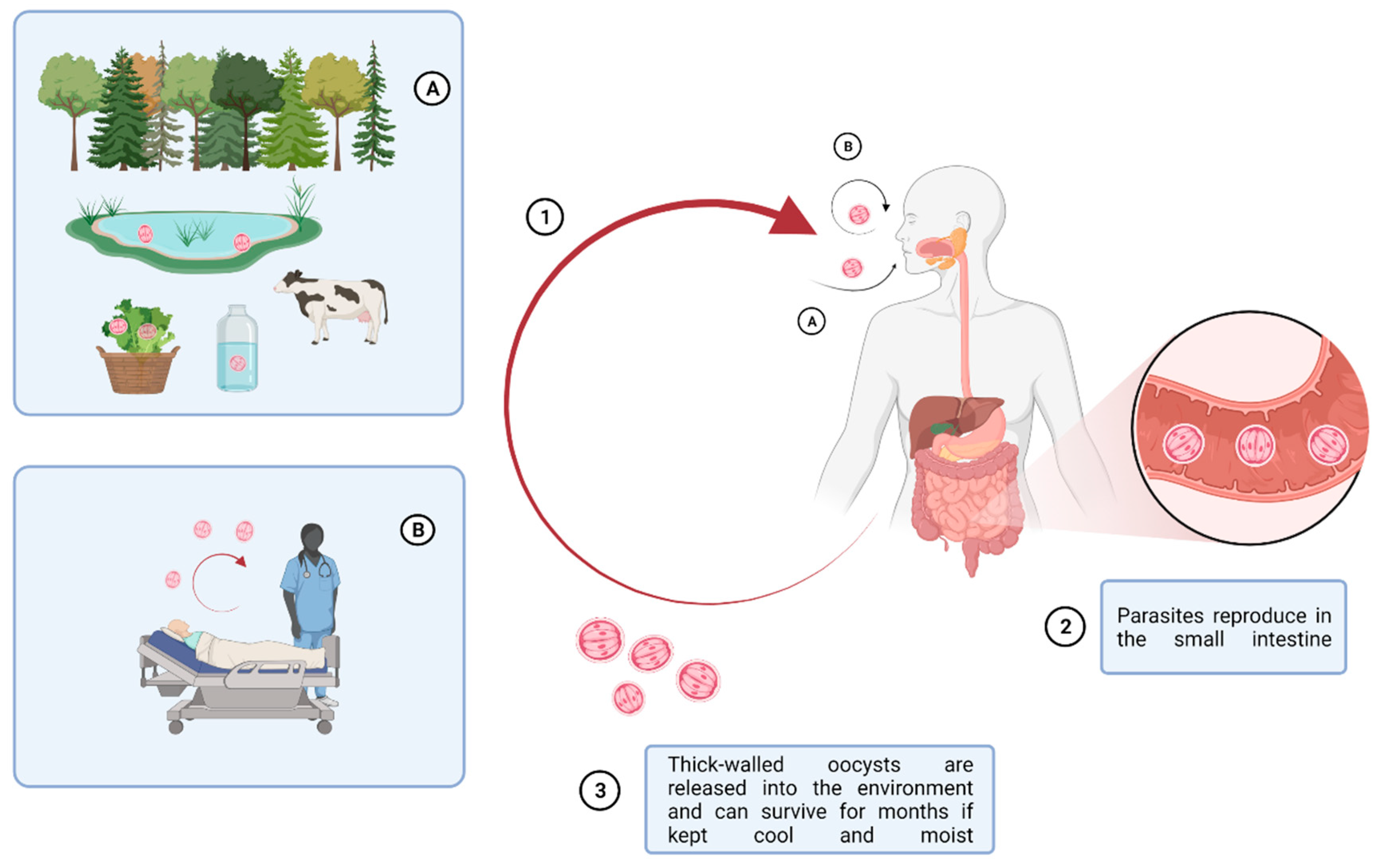
Figure 1.
Multiple factors affecting the routes of Cryptosporidium spp. infection cycle. Zoonotic Cryptosporidium parasites are transmitted from livestock through long-lived oocysts in their faces, which can contaminate the environment, water, and food, producing a source of infection to people. Besides water- and food-borne transmissions, inhalation of oocysts has been described as another mode of transmission. (1) oocyst ingestion by (A) fecal-oral route and (B) inhalation; (2) parasite reproduction in the small intestine; (3) release of oocysts in the environment. This picture was created with BioRender.com accessed on 20 May 2019.
Figure 1.
Multiple factors affecting the routes of Cryptosporidium spp. infection cycle. Zoonotic Cryptosporidium parasites are transmitted from livestock through long-lived oocysts in their faces, which can contaminate the environment, water, and food, producing a source of infection to people. Besides water- and food-borne transmissions, inhalation of oocysts has been described as another mode of transmission. (1) oocyst ingestion by (A) fecal-oral route and (B) inhalation; (2) parasite reproduction in the small intestine; (3) release of oocysts in the environment. This picture was created with BioRender.com accessed on 20 May 2019.

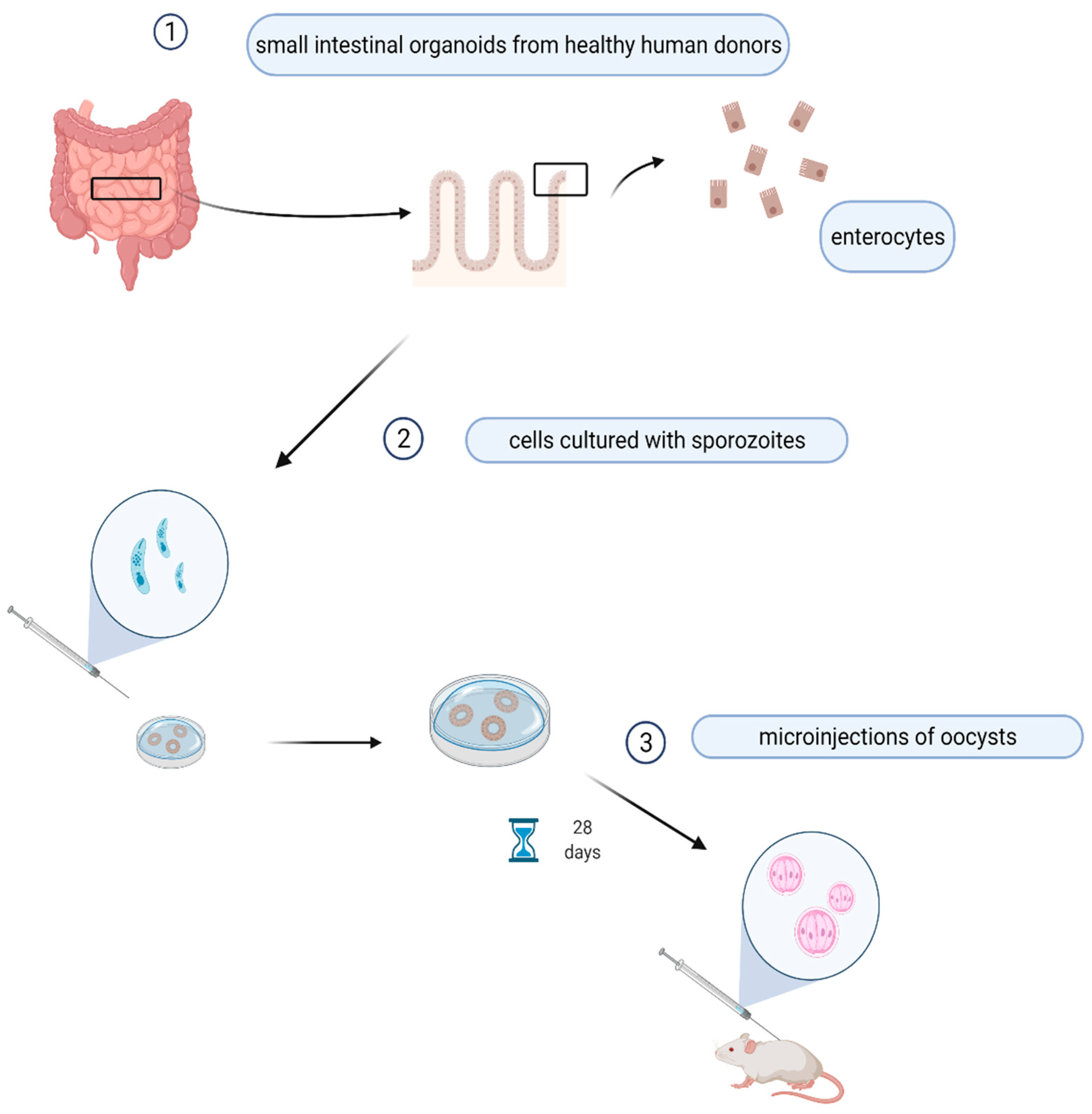
Figure 2.
Prototype of organoids’ simulating in vivo growth of Cryptosporidium spp. Enterocytes from patient’s intestine are isolated and cultured with sporozoites for 28 days, then the organoides are injected in the mouse. This picture was created with BioRender.com accessed on 20 May 2019.
Figure 2.
Prototype of organoids’ simulating in vivo growth of Cryptosporidium spp. Enterocytes from patient’s intestine are isolated and cultured with sporozoites for 28 days, then the organoides are injected in the mouse. This picture was created with BioRender.com accessed on 20 May 2019.


{kind=link}
{kind=link}
Table 1.
Cryptosporidium species identification, confirmed by polymerase chain reaction (PCR) and sequence analyses in animals and in humans.
Table 1.
Cryptosporidium species identification, confirmed by polymerase chain reaction (PCR) and sequence analyses in animals and in humans.
| Species Name | Major Host(s) | Human Host | Author(s) |
|---|---|---|---|
| Cryptosporidium cuniculus | rabbits | yes | [11] |
| Cryptosporidium hominis | equine | yes | [12,13] |
| Cryptosporidium parvum | sheep, cattle | yes | [13,14] |
| Cryptosporidium muris | rodents and farm animals | yes | [13,15] |
| Cryptosporidium andersoni | cattle | yes | [13,16,17] |
| Cryptosporidium suis | pigs | yes | [13,18,19] |
| Cryptosporidium wrairi | guinea pigs | yes | [13,16] |
| Cryptosporidium meleagridis | birds | yes | [13] |
| Cryptosporidium canis | dogs | yes | [13,17] |
| Cryptosporidium ubiquitum | ruminants and rodents | yes | [18] |
| Cryptosporidium xiaoi | sheep and goats | yes | [18,19] |
| Cryptosporidium felis | cats | yes | [20,21] |
| Cryptosporidium nasorum | fish | not | [22] |
| Cryptosporidium molnari | fish | not | [23] |
| Cryptosporidium scophthalmi | fish | not | [24] |
| Cryptosporidium scrofarum | pigs | yes | [25,26] |
| Cryptosporidium baileyi | chicken | yes | [27,28] |
| Cryptosporidium galli | chicken | not | [29] |
| Cryptosporidium fragile | amphibia | not | [30] |
| Cryptosporidium serpentis | snakes | not | [31] |
| Cryptosporidium varanii | pet reptiles | not | [32] |
| Cryptosporidium ryanae | cattle | not | [33] |
| Cryptosporidium bovis | cattle | yes | [34,35] |
| Cryptosporidium fayeri | marsupials | yes | [36,37,38] |
| Cryptosporidium macropodum | marsupials | not | [39] |
| Cryptosporidium tyzzeri | mice | yes | [40,41] |
| Cryptosporidium viatorum | rodents | yes | [42] |
| Cryptosporidium occultus | rodents | yes | [43] |
| Cryptosporidium proventriculi | birds | not | [44] |
| Cryptosporidium ornithophilus | ostrich | not | [45] |
| Cryptosporidium ratti | rats | not | [46] |
| Cryptosporidium erinacei | hedgehogs | yes | [41,47] |
| Cryptosporidium sciurinum | red squirrels | not | [48] |
| Cryptosporidium myocastoris | nutria | not | [49] |
| Cryptosporidium testudinis | tortoises | not | [50] |
| Cryptosporidium avium | birds | not | [51] |
| Cryptosporidium alticolis | common voles | not | [52] |
| Cryptosporidium microti | common voles | not | [52] |
| Cryptosporidium abrahamseni | fish | not | [53] |
| Cryptosporidium bollandi | fish | not | [54] |
| Cryptosporidium apodemi | rats | not | [55] |
| Cryptosporidium ditrichi | rodents | yes | [55,56] |
| Cryptosporidium ducismarci | tortoises | not | [50] |
| Cryptosporidium proliferans | rodents | not | [57] |
Table 2.
Use of drugs for the treatment of human cryptosporidiosis.
| Treatment | Clinical Improvement | Protocols | Author(s) |
|---|---|---|---|
| Nitazoxanide | inhibits oocyst excretion | clinical trials | [90] |
| Paromomycin | no clinical benefits | clinical trials | [91] |
| Spiramycin | oocyst reduction | in vitro and in vivo studies | [92] |
| Macrolides (Azithromycin, Erythromycin, Roxithromycin) | no clear clinical benefits | in vitro and in vivo studies | [93] |
| Clofazimine | oocyst reduction | clinical trials | [94] |
| Benzoxaboroles, Pyrazolopyridine | oocyst reduction | in vivo studies | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pane, S.; Putignani, L. Cryptosporidium: Still Open Scenarios. Pathogens 2022, 11, 515. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050515
AMA Style
Pane S, Putignani L. Cryptosporidium: Still Open Scenarios. Pathogens. 2022; 11(5):515. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050515
Chicago/Turabian StylePane, Stefania, and Lorenza Putignani. 2022. "Cryptosporidium: Still Open Scenarios" Pathogens 11, no. 5: 515. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050515
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.