
Hemoglobin Endocytosis and Intracellular Trafficking: A Novel Way of Heme Acquisition by Leishmania


Kusuma School of Biological Sciences, Indian Institute of Technology, Hauz Khas, New Delhi 110016, India
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Pathogens 2022, 11(5), 585; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050585
Submission received: 21 March 2022
/
Revised: 10 May 2022
/
Accepted: 13 May 2022
/
Published: 16 May 2022
(This article belongs to the Special Issue Leishmania & Leishmaniasis)
Abstract
:Leishmania species are causative agents of human leishmaniasis, affecting 12 million people annually. Drugs available for leishmaniasis are toxic, and no vaccine is available. Thus, the major thrust is to identify new therapeutic targets. Leishmania is an auxotroph for heme and must acquire heme from the host for its survival. Thus, the major focus has been to understand the heme acquisition process by the parasites in the last few decades. It is conceivable that the parasite is possibly obtaining heme from host hemoprotein, as free heme is not available in the host. Current understanding indicates that Leishmania internalizes hemoglobin (Hb) through a specific receptor by a clathrin-mediated endocytic process and targets it to the parasite lysosomes via the Rab5 and Rab7 regulated endocytic pathway, where it is degraded to generate intracellular heme that is used by the parasite. Subsequently, intra-lysosomal heme is initially transported to the cytosol and is finally delivered to the mitochondria via different heme transporters. Studies using different null mutant parasites showed that these receptors and transporters are essential for the survival of the parasite. Thus, the heme acquisition process in Leishmania may be exploited for the development of novel therapeutics.

1. Introduction
Leishmaniasis is one of the most devastating tropical diseases, which is a threat to more than 0.7–1 million people in 100 endemic countries worldwide [1]. The disease is caused by 20 different species of the unicellular protozoan parasite, Leishmania [2]. The parasite has two life forms; one is the flagellated promastigote form, which inhabits the midgut of the phlebotomine sandfly, and the other is the non-motile amastigote form, which resides in a mammalian host. Leishmaniasis has been mainly classified into four clinical subtypes based on the symptoms: visceral leishmaniasis (kala-azar), cutaneous leishmaniasis, mucocutaneous leishmaniasis and post-kala-azar dermal leishmaniasis (PKDL). In cutaneous leishmaniasis, a skin ulcer forms at the site of an infected sandfly bite. It is mostly self-curable, but the time of healing varies depending on the Leishmania species and infected individuals. L. mexicana, L. amazonensis and L. tropica are the major causative agents of cutaneous leishmaniasis. Mucocutaneous leishmaniasis manifests as an infection in the mucosal membrane, which leads to progressive destructive ulcers in the mucosa, spreading from the mouth and nose to the larynx and pharynx. It is predominantly caused by L. braziliensis. Visceral leishmaniasis (VL) is the most severe form of the disease, which is caused by L. donovani, L. infantum and L. chagasi. It is a systemic disease that is deadly if left untreated [2,3]. For more than 70 years, the antimonial drugs meglumine antimoniate and sodium stibogluconate have been used for the treatment of leishmaniasis [4]. Antimonial drugs are toxic and have adverse reactions, including acute pancreatitis and cardiac arrhythmia [2]. Due to the high toxicity of antimonial drugs and the appearance of drug resistance, Amphotericin B, Miltefosine and Paromomycin are used as alternative anti-leishmaniasis drugs [5,6,7]. However, each of them has severe side effects, which include hypokalemia, nephrotoxicity and hepatic transaminitis [8,9]. In addition, there is no appropriate vaccine available for leishmaniasis [10]. Therefore, there is a need to develop better chemotherapeutic agents against leishmaniasis. Understanding the biology of Leishmania with respect to growth, differentiation and key intracellular processes may help in the identification of novel molecular targets for therapeutic intervention of leishmaniasis. One of the best ways to identify new drugs for the disease is to exploit the biochemical difference between the host and the parasites. Leishmania lacks a heme biosynthetic pathway and depends on an exogenous supply of heme for its survival, presumably from the host cells. Thus, understanding the heme acquisition processes by the parasite is an attractive area to identify new therapeutic targets against leishmaniasis.
2. Heme Acquisition: An Essential Process for Parasite
Heme is an essential cofactor for various biological processes such as electron transport chain, oxygen transport and storage (hemoglobin and myoglobin), metabolism of drugs, transcriptional regulation and signal transduction. It is an iron-containing porphyrin (protoporphyrin IX), in which most of the porphyrins contain iron as a central molecule. The central iron molecule can be oxidized into the ferric (Fe+3) or ferrous (Fe+2) oxidative state [11]. Different types of heme are present, such as heme A, heme B and heme C; the most common heme is heme B. Heme B is present in hemoglobin, myoglobin and other heme proteins. Heme is vital for most organisms such as archaea, bacteria and eukaryotes. In photosynthetic eukaryotes, heme biosynthesis initiates from the binding of glutamate to tRNAGlu. However, in heterotrophic eukaryotes, heme synthesis starts with the condensation of glycine with succinyl Co-A [12]. The rest of the seven steps in the heme biosynthetic pathway are evolutionarily conserved in most organisms. In contrast, the heme biosynthetic pathway is fully or partially absent in flagellated kinetoplastid parasites. For example, Leishmania can perform only the last three steps of the heme biosynthetic pathway in mitochondria, which involves coproporphyrinogen oxidase, protoporphyrinogen oxidase and ferrochelatase [13]. However, Trypanosoma lacks a complete heme biosynthetic pathway [14]. Therefore, these trypanosomatid parasites need to scavenge heme from their host for their survival.
3. Hemoglobin: Potential Source of Heme
As Leishmania is an auxotroph for heme, parasites require heme or pre-formed porphyrins for their growth from the extracellular milieu [15] and possibly rely on the host cell for the supply of heme [16]. However, free heme is toxic as it produces reactive oxygen species and damages the cell membrane [17]. Therefore, free heme/iron is almost not available in the host cells. Most of the heme in the cells is present in a complex with iron-binding proteins such as transferrin, lactoferrin, hemoglobin and plasma ferritin [18]. Incidentally, mammalian cells also import iron by endocytosis of iron-bound transferrin via the transferrin receptor [19], whereas hemoglobin is endocytosed via the CD163 receptor on macrophages [20]. Similarly, many bacterial [21,22] and fungal [23] pathogens have evolved various mechanisms to uptake host hemoproteins and extract heme from these molecules to meet their requirement. Presumably, Leishmania also salvages heme from macrophage hemoproteins such as Hb. This is well supported in the life cycle of Leishmania (Figure 1), as both amastigotes in the mammalian host and promastigotes in the insect vector have access to host Hb. The intracellular amastigote form of Leishmania resides in the macrophages, which take senescent RBC [24] and endocytose Hb by CD163 [20]. Interestingly, it was also shown that Leishmania infected macrophages phagocytose of more RBCs, possibly to acquire heme from the Hb present in erythrocytes [25]. Similarly, promastigote form in the insect gut has access to Hb from the lysed RBC in the sandfly gut [26]. This is supported by the fact that Leishmania can be grown in vitro in a serum-free culture medium containing blood lysate [27], and the blood lysate substituted with Hb in the culture medium renders them to grow even better [28]. In addition, the heme requirement by the parasite is met by the addition of heme directly into in vitro culture medium [29] for parasite growth. Since the free heme is not present in the bloodstream of the host and the fact that Hb can be used as a heme source at least in vitro, it is possible that Leishmania parasites facilitate the uptake of host cell hemoproteins to salvage heme by intracellular degradation of internalized hemoprotein. Thus, the presence of some proteins/receptors mediating the Hb endocytosis is crucial for the growth of the parasites. Therefore, it is necessary to understand the mechanism of endocytosis and intracellular trafficking of Hb to the appropriate destinations in the parasite. Here, we briefly summarized the intracellular trafficking processes in mammalian cells and compared similar processes in Leishmania to understand the regulation of the Hb trafficking in the parasite.
4. Endocytosis and Intracellular Trafficking in Mammalian Cells
Endocytosis is a process by which cells internalize various molecules from the extracellular milieu and play a key role in different processes such as nutrient uptake, antigen presentation and maintenance of cellular homeostasis. There are various modes of endocytosis, such as phagocytosis, macropinocytosis, clathrin-mediated endocytosis, caveolae-mediated endocytosis and clathrin or caveolae independent endocytosis [30]. After binding to the cell surface receptor, internalized cargoes are trafficked to the early endosomes, and subsequently, they are sorted to different intracellular destinations. The transfer of cargo from the donor membrane to the acceptor vesicle occurs through a coordinated series of vesicle fusion events specifically regulated by Rab GTPases and SNARE proteins [31]. Fusion between two particular compartments is regulated by a specific Rab along with cognate SNAREs to ensure proper targeting of the cargo to the appropriate intracellular destination.
The intracellular trafficking pathways can be broadly divided into endocytic pathways and secretory pathways. In the endocytic pathway, cells internalize material from the extracellular environment into the cell by binding with a specific cell surface receptor. Subsequently, ligands are internalized into early endosomes, where they are sorted to different destinations depending on the receptor system. Finally, ligands are trafficked to the lysosomes via late endosomes, where the cargo is degraded by the hydrolases present in the low pH of the lysosome. The degradation of the cargo in the lysosomes generates essential molecule, which is utilized by the cells depending on the type of ligand. The secretory pathway or exocytic pathway is involved in the trafficking of newly synthesized proteins, lipids and carbohydrates to various locations such as plasma membrane, endo-lysosomal system, mitochondria and the secretion of molecules outside the cell. It is a highly organized process that includes various steps involving post-translational modifications, packaging and targeting of newly synthesized proteins to their specific locations. Most of the newly synthesized proteins that are synthesized in the rough endoplasmic reticulum or in the cytosolic ribosomes follow the conventional secretory pathway via the trafficking of proteins through the ER-Golgi network to its target location. Any deviation from the classical/conventional pathway is termed as a non-conventional or uncanonical secretory pathway.
Rab GTPases are master regulators of vesicle trafficking, as evident from studies in mammalian cells. They regulate vesicle motility and membrane fusion through the recruitment of specific SNARE proteins and various effector molecules [31]. There are about 70 Rab GTPases in humans that are localized on distinct membrane compartments (Figure 2) and regulate the vesicular trafficking in the appropriate direction [32]. In the endocytic pathway, Rab5 localizes on the early endosomes and mediates the transport from the cell surface to early endosomes [33,34]. In contrast, Rab7 localizes in the late endosomes and regulates the transport of cargo from the early to the late endosome/lysosomes [35,36]. Rab4 involves the fast recycling of surface proteins from early endosomes, and Rab11 regulates the slow recycling of cargoes from endocytic recycling compartment [34,37]. Rab9 and Rab24 are also localized on the late endosomes. Rab9 mediates the trafficking of lysosomal enzymes from TGN to the lysosome [32,35]. In the secretory pathway, Rab1 regulates the anterograde transport from ER to Golgi whereas Rab2 mediates the retrograde transport from the Golgi to the ER. In addition, intra-Golgi transport is regulated by several Rabs, namely Rab6, Rab12, Rab13, Rab33, etc. Conversely, Rab3 and Rab8 regulate the transport of secretory protein from the Golgi to the extracellular milieu [31,38].
SNAREs are another group of coiled-coil proteins that confer specificity to vesicular fusion events in association with Rab GTPases, and approximately 36 members of this family of proteins are reported in mammalian cells [39]. Similar to Rab GTPases, specific SNARE also localizes on distinct intracellular compartments (Figure 3). In the endocytic pathway, Syntaxin13 localizes on the early endosomes and regulates trafficking from the early endosomes to the recycling endosomes, whereas early endosomal Syntaxin8 controls trafficking between early compartments. In contrast, Syntaxin7 plays a major role in regulating trafficking from the endosomes to the lysosomes [40,41]. Thus, the internalization of the ligands and transport to the lysosomal compartments is regulated by several endocytic Rabs and SNAREs, whereas transport of the newly synthesized receptor to the cell surface is controlled by secretory Rabs and SNAREs. However, intracellular trafficking in unicellular parasites is not well documented, apart from a few reports in Trypanosomatid parasites. Thus, it is important to characterize the intracellular trafficking pathway in Leishmania to understand the regulation of Hb endocytosis in the parasite.
5. Intracellular Trafficking in Leishmania
Leishmania is a protozoan parasite, and intracellular trafficking in this parasite is not yet characterized. However, recent reports emerging from Trypanosoma indicate that some of the Rabs and SNAREs are also conserved in this parasite [42,43]. Simultaneously, we also identified and characterized several Rabs in Leishmania to understand the intracellular trafficking pathways in the parasite.
Leishmania expresses a wide range of surface proteins to form protective receptors for nutrient uptake. Leishmania forms a protective coat on its surface known as glycocalyx, which is composed of lipophosphoglycan (LPG), proteophosphoglycan, glycophosphatidyl inositol (GPI) anchored proteins, free GPI glycolipids and glycoinositol phospholipids [44,45]. Leishmania also secretes several virulence factors into the host cells, such as gp63 (a metalloprotease), cysteine protease and LPG, which modulate the host cell signaling machinery for the survival of the intracellular parasite [46,47]. Among them, Ldgp63 is the major virulence factor [48,49]. Both endocytosis and exocytosis occur through the flagellar pocket of Leishmania, but the regulation of endocytic and secretory pathways are not characterized in the parasite. We initiated the studies to determine the role of different Rab GTPases in the regulation of intracellular trafficking in Leishmania.
We showed that Leishmania has a well-developed clathrin-mediated endocytic pathway such as mammalian cells [50]. The endocytic ligand binds to the receptor and recruits clathrin via the adaptor to form coated vesicles in Leishmania. Finally, the coated vesicle is pinched off from the plasma membrane by dynamin homologs in the parasite (unpublished data). We cloned and expressed early endosomal Rab5 from Leishmania and showed that Rab5 regulates the homotypic fusion between early endosomes [51]. Subsequently, we found that Leishmania donovani has two isoforms of LdRab5 (LdRab5a and LdRab5b), and both of them are localized in the early endocytic compartment. LdRab5a regulates fluid-phase endocytosis, whereas LdRab5b controls receptor-mediated endocytosis in the parasite [52]. The function of LdRab5b is essential for the parasites as Rab5b knockout parasites are unable to survive in the macrophages, probably due to the unavailability of the heme from Hb [53]. We showed that Rab7 in Leishmania localizes in the late endosomes and mediates the transport of cargo from the early to the late/lysosomal compartment, such as mammalian cells [54]. Leishmania also has a well-conserved recycling pathway, which is mediated by Rab4 and Rab11 homologs in the parasite (unpublished data).
We also analyzed the secretory pathway in the parasite, as this route is important for the secretion of virulence factors and the transport of newly synthesized receptors to the cell surface. Leishmania has a well-conserved Rab1 homolog that localizes in the Golgi. Our results showed that overexpression of the dominant negative mutant of LdRab1 blocks the secretion of secretory acid phosphatase and Ldgp63 [55]. We also found that newly synthesized proteins such as gp63 exit from the ER through a well-defined COPII complex consisting of LdSar1 GTPase, LdSec23, LdSec24 LdSec31 and LdSec13. Ldgp63-containing COPII vesicle budding from the ER is inhibited by LdSar1:T34N, a dominant negative mutant of LdSar1, and consequently blocks Ldgp63 trafficking and secretion in Leishmania [56]. These results suggest that Leishmania has a conventional secretory pathway such as mammalian cells. Bioinformatics analysis predicts that Leishmania genomes have several putative Rabs (Table 1) and SNARE [57] domain-containing proteins (Table 2), which need to be functionally characterized to understand the intracellular trafficking pathways in the parasite.
6. Heme Acquisition Processes in Leishmania
Leishmania is an auxotroph for several essential nutrients such as heme, folate, etc. They need to import these essential molecules from the extracellular milieu or from the host cells for their survival. Thus, identification of the processes by which parasites meet the requirement of these essential nutrients is of paramount importance for the development of novel therapeutic strategies. However, free heme is not available in the host cells under normal physiological conditions, and it is only present as a complex with several iron-binding proteins such as hemoglobin, transferrin, lactoferrin, ferritin, etc. Therefore, it is possible that the parasite may acquire heme/iron-containing proteins by endocytosis via the expression of a specific receptor or transporter on the parasite surface. Subsequently, various receptors/transporters were identified in Leishmania, which mediate endocytosis of these molecules [58]. For example, an LDL receptor-like molecule was identified on the surface of Leishmania, possibly for scavenging cholesterol and lipids from LDL particles; however, the LDL endocytic process is not properly characterized in the parasite [59]. Trypanosomatid parasites are also unable to synthesize purines de novo and therefore must acquire these nutrients from their hosts. Consequently, purine transporters such as LdNT1 and LdNT2 in L. donovani [60,61] and NT3 and NT4 are identified in L. major [62]. Leishmania is also a folate auxotroph and thus depends on the uptake of folate from the environment. It was shown that Leishmania has more than one active folate transporter. Among these, FT5 is a very high-affinity folate transporter in the parasite mediating the imports of folates and related molecules under varying conditions [63]. These high-affinity active transporters enable the parasites to efficiently compete with their hosts for the acquisition of these nutrients. Thus, the parasite expresses a plethora of transporters and receptors and follows different ways to import essential nutrients from their hosts. However, the heme/iron uptake process in Leishmania was not well characterized until recently. The following sections focus on recently identified different mechanisms that are involved in heme/iron acquisition in Leishmania.
6.1. Transferrin: As a Source of Iron by Leishmania
It is well characterized that mammalian cells import iron via the transferrin (Tf) receptor [19]. Iron-loaded Tf (holo Tf) binds with specific high-affinity receptors (TfR) on the surface of the mammalian cells and subsequently internalizes into the early endosomes via the clathrin-mediated process. In the acidified endosomal compartment, the Tf-TfR complex releases ferric iron and Tf without Fe (apoTf) along with the bound receptor, which recycles back to the plasma membrane via endocytic recycling compartments [64]. Therefore, studies were initiated to understand the role of transferrin in the iron uptake by Leishmania. Interestingly, a 70 kDa transferrin binding protein was detected in L. infantum and L. major cell lysates, and a monoclonal antibody against this protein recognizes human TfR, indicating that the putative protein is potentially a transferrin receptor in Leishmania [65]. Subsequently, the protein is purified from Leishmania promastigotes using affinity chromatography on a transferrin-Sepharose column. Further characterization reveals that the Leishmania transferrin receptor is an integral membrane glycoprotein of a single 70-kDa polypeptide, unlike the disulfide-linked dimer of the mammalian TfR. This Tf binding protein is uniformly distributed on the surface of the parasite; however, the role of this protein in transferrin uptake by Leishmania is not properly documented [66]. However, it is possible that the intracellular amastigote form acquires iron from the human transferrin as it was shown that L. mexicana containing parasitophorous vacuoles (PV) in macrophages fuse with the transferrin receptor-positive early endosomal compartment 10 days after infection and blocks the TfR receptor recycling in the macrophages. Thus, Leishmania amastigotes presumably recruit TfR on PV and import iron via human Tf through the flagellar pocket [67,68]. However, transferrin endocytosis in Leishmania needs to be further characterized.
6.2. Lactoferrin: As a Source of Iron by Leishmania
Apart from transferrin, parasites may also obtain iron from lactoferrin, a member of the transferrin family of proteins that are found in most of the biological fluids in mammals. Surprisingly, lactoferrin binding protein is identified as glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in mammalian cells, which also binds transferrin [69,70]. It was shown that L. chagasi promastigotes are able to take up 59Fe-lactoferrin more efficiently than the 59Fe-transferrin. This uptake process is saturable and is inhibited by the addition of 10-fold excess of unlabeled ferrilactoferrin, suggesting the existence of a putative lactoferrin receptor on the surface of L. chagasi promastigotes. Moreover, the parasite is efficiently grown in vitro in iron-depleted serum containing minimal essential medium supplemented with lactoferrin, indicating the acquisition of iron by Leishmania from lactoferrin [71]. Subsequent studies showed that lactoferrin also binds to the similar 70-kDa protein in promastigote, which is initially identified as transferrin binding protein in Leishmania. Thus, this binding site is not specific to any ferroprotein. It is possible that promastigotes preferentially take up iron in a reduced form rather than an oxidized form, as it was shown that promastigotes have NADPH-dependent iron reductase activity, which is discussed in detail in the heme/iron transporters section. Thus, extracellular iron is possibly reduced prior to its internalization [72]. These results suggest that the parasite possibly utilizes various iron sources to survive in the diverse environments in the insect and the mammalian hosts.
6.3. Hemoglobin: As a Source of Heme by Leishmania
Hemoglobin is the major heme carrier protein in the mammalian cells, which is sequestered in RBCs. As aged RBCs are phagocytosed by macrophages and are degraded in the phagolysosomal compartment in macrophages where Leishmania resides, the parasite may acquire heme from the host Hb coming from degraded RBCs [73]. The first clue of Leishmania using Hb as a source of heme comes from the observation that parasites can be grown in vitro in a serum-free culture medium containing blood lysate [27] or Hb [28]. Therefore, it is possible that Leishmania may have developed a process to take up the Hb and salvage heme by intracellular degradation of internalized Hb. With this notion, we initiated studies to identify the Hb receptor (HbR) in Leishmania.
We showed that Hb specifically binds with a putative high-affinity binding site present in the flagellar pocket of L. donovani promastigotes with saturation kinetics. Subsequently, bound Hb is rapidly internalized into the parasites and is degraded in the lysosomal compartment and thereby releasing the heme from intracellular degradation of the Hb. The Hb binding with Leishmania is specifically competed by unlabeled Hb but not by globin or hemin or other heme- or iron-containing proteins indicating that the parasite receptor specifically recognizes Hb. Finally, a 46 kDa protein was identified as a Hb binding protein from the detergent-solubilized surface membrane preparation of Leishmania promastigotes using hemoglobin-agarose affinity chromatography [74]. In order to understand the role and nature of the 46-kDa protein in Hb endocytosis in the Leishmania, the 46-kDa protein was purified to homogeneity from Leishmania promastigote membrane. The purified protein specifically binds Hb and antibodies against the purified protein and blocks Hb endocytosis in promastigotes, demonstrating that the 46-kDa protein is indeed a Hb-receptor (HbR) in Leishmania. Consequently, the full-length HbR gene is cloned, and recombinant protein is purified. To our surprise, sequence analysis of the cloned HbR revealed the presence of hexokinase (HK) signature sequences, ATP-binding domain and PTS-II motif, indicating that the cloned protein may be a hexokinase from Leishmania. It is puzzling, but cell lysate prepared from HbR-overexpressing Leishmania promastigotes shows enhanced HK activity in comparison with the untransfected cells. Moreover, HbR-overexpressing cells bind and internalize Hb more efficiently than untransfected control parasites. In summary, these results demonstrate that HbR in Leishmania is a hexokinase [75]. However, these results also open up several intriguing areas for future studies. For example, how is hexokinase targeted to the flagellar pocket to bind Hb? It is possible that parasites may generate two species of hexokinase either by using different start sites or by alternative splicing? Subsequently, full-length protein is transported to glycosomes and acts as a hexokinase, whereas the truncated protein is targeted to the flagellar pocket to be a HbR as we found that PTS deleted protein is predominantly localized in the flagellar pocket (unpublished data). We are currently evaluating how it is happening within the parasite. We also tried to determine how newly synthesized HbR is transported to the flagellar pocket from the ER-Golgi network in Leishmania. Our results suggest that though Leishmania has a conventional Rab1 dependent secretory pathway, newly synthesized HbR traffic from the ER-Golgi network to the flagellar pocket follows a Rab1 independent, unconventional secretory pathway. Similarly, another open question is to determine whether hexokinase activity of HbR is needed for Hb endocytosis. However, we found that mutation in the hexokinase motif in the HbR completely abrogates the hexokinase activity of the protein, but hexokinase defective mutant HbR still binds Hb in vitro (unpublished data). It would be interesting to explore whether any other downstream accessory protein in Hb endocytosis machinery requires hexokinase activity of the HbR.
Further characterization reveals that the N-terminus of the HbR is the extracellular Hb-binding domain, and the C-terminus is the cytoplasmic signaling domain of HbR [51,75]. Interestingly, the HbR sequence is well conserved in different species and strains of L. amazonensis, L. infantum, L. donovani, L. major, L. turanica, L. gerbilli, L. tropica and L. enrietti, with 97 to 100% sequence identity indicating that other species of Leishmania possibly use HbR for heme acquisition [76]. In addition, immunolocalization studies demonstrate that the HbR is also present in intracellular amastigote form [77] and mediates Hb uptake in amastigotes [53]. More recently, a 40 amino acids sequence (SSEKMKQLTMYMIHEMVEGLEGRPSTVRMLPSFVYTSDPA) in the N-terminal of HbR was identified as a Hb-binding domain [78]. This peptide completely blocks the Hb uptake in both promastigote and amastigote forms of Leishmania and thereby inhibits the growth of the parasite. Thus, this mimicking peptide may be used as a potential therapeutic agent against Leishmania.
Subsequently, studies were carried out to understand how receptor-bound Hb is internalized and is trafficked to appropriate intracellular destination to generate intracellular heme from the degradation of internalized Hb. It is well established in mammalian cells that extracellular ligands are internalized either by fluid-phase or clathrin-mediated endocytosis [79,80]. Among these modes of endocytosis, the most well-studied process is clathrin-mediated endocytosis. In the clathrin-mediated endocytosis, the cytoplasmic domain of the receptor is generally activated upon ligand binding, which interacts with endocytic adaptors, which in turn bind to the clathrin to form coated pits [81]. Finally, Dynamin GTPase cleaves the coated pits from the plasma membrane and internalizes the cargo in coated vesicles [82]. In order to understand the modes of Hb internalization in Leishmania, the clathrin heavy chain from Leishmania (Ld-CHC) is cloned and overexpressed in the parasites. Interestingly, Ld-CHC is shown to localize in the flagellar pocket of Leishmania along with bound Hb. Kinetic analysis of intracellular trafficking of Hb in Ld-CHC overexpressing Leishmania promastigotes reveals that Hb is associated with Ld-CHC coated region at early time points of internalization, and subsequently, Ld-CHC dissociates from the Hb-containing vesicles, indicating that clathrin-coating and uncoating regulate Hb trafficking in Leishmania. In addition, overexpression of dominant negative mutant of clathrin heavy chain of Leishmania (GFP-Ld-CHC-Hub) completely blocks the Hb internalization and causes severe growth defect in the parasite, demonstrating that the Hb internalization by clathrin-mediated endocytosis is essential for Leishmania [50]. After high-affinity binding of Hb with the HbR in clathrin coated-pits, coated vesicles are cleaved by Dynamin homologs in Leishmania (unpublished data) and internalize into discrete intracellular vesicles, presumably early endosome-like compartments within 5 min, as visualized by the internalization of gold conjugated Hb [74]. This is the first demonstration that Hb trafficking in Leishmania is probably regulated by vesicle fusion, as shown in the higher eukaryotic cell.
As Rab GTPases are master regulators of the intracellular trafficking pathway by vesicle fusion [38], therefore endocytic Rabs such as Rab5 (LdRab5) and Rab7 (LdRab7) are cloned from Leishmania to understand the intracellular trafficking of Hb in the parasite. Subsequently, Hb-containing early endosomes are isolated from Leishmania promastigotes and determine the role of LdRab5 in homotypic fusion between endosomes in vitro to understand the trafficking of the Hb in Leishmania. It was shown that homotypic fusion between the early endosomes in Leishmania is regulated by LdRab5, whereas fusion between early endosomes with late endosomes is dependent on LdRab7 [51]. These findings suggest that the early step of Hb endocytosis is regulated by LdRab5, and transport of the Hb from the early endosome to the late endosomes/lysosome is mediated by LdRab7. In order to unequivocally prove the role of LdRab5 and LdRab7 in Hb trafficking in vivo, transgenic parasites are generated by overexpressing the LdRab5, LdRab7 or their dominant negative mutants. Incidentally, it was found that Leishmania has two isoforms of Rab5, namely LdRab5a and LdRab5b. LdRab5a is shown to regulate the fluid phase endocytosis, whereas LdRab5b controls the receptor-mediated endocytosis of the Hb in the parasite. Kinetic analysis of the Hb endocytosis using Alexa594 labeled Hb in promastigotes showed that Hb enters into LdRab5-positive early endosome within 5 min of internalization and is subsequently trafficked to the LysoTracker-positive lysosomes at about 45 min where Hb is degraded to generate intracellular heme as no Hb is detected at about 60 min after internalization. This process of lysosomal transport is accelerated in the transgenic parasites overexpressing LdRab5a:WT, whereas bound Hb is stuck in the flagellar pocket in the parasites overexpressing LdRab5b:N133I, a dominant negative mutant of LdRab5b [52]. Subsequently, LdRab5b-/- null-mutant parasites are generated, and these parasites show severe growth defects and are unable to internalize Hb. However, these cells are rescued by exogenous addition of hemin in the growth medium, demonstrating that Leishmania internalizes Hb in Rab5b dependent process and generates intracellular heme by lysosomal degradation for its survival [53]. Interestingly, LdRab5b-/- parasites can infect the macrophages but are unable to survive after 96 h of infection in comparison to the normal parasite. These results suggest an interesting possibility of using Rab5b null-mutant Leishmania as a new vaccine candidate, as these parasites can infect macrophages but are unable to survive in macrophages, and therefore, it may generate a protective immune response in an infected host. In order to understand the mechanism of trafficking of internalized Hb to lysosomes in Leishmania, similar studies were carried out using transgenic parasites overexpressing LdRab7 or its mutants. Interestingly, LdRab7 is shown to localize both on the early and the late endocytic compartments in Leishmania and thereby connecting both early and late events of Hb endocytosis. Overexpression of LdRab7:WT in Leishmania induces transport of internalized Hb to lysosomes, where it is rapidly degraded. In contrast, transgenic parasites overexpressing LdRab7:T21N, a GDP-locked mutant of Rab7, significantly inhibit the degradation of internalized Hb by blocking its transport to the lysosomes. Most importantly, parasites overexpressing LdRab7:T21N show growth defects possibly due to the nonavailability of heme from Hb degradation. This is supported by the fact that the addition of exogenous hemin recovers the growth of these cells almost to the control level. Thus, Rab7-mediated transport of internalized Hb to the lysosomes is required for the generation of intracellular heme from Hb degradation in Leishmania [54].
Thus, Leishmania devices a novel process to salvage heme from the Hb. In summary, Leishmania expresses a high-affinity specific receptor for Hb in the flagellar pocket, which internalizes bound Hb by clathrin-mediated endocytosis. Subsequently, the internalized Hb is transported to the lysosomes via early and late compartments, where it is degraded to generate intracellular heme, which is used by the parasite for its survival (Figure 4). Thus, blocking the Hb endocytosis by an appropriate inhibitor is an attractive area to explore for the development of new therapeutic molecules. In this notion, immunization with the HbR is found to induce sterile protection against the infection with virulent parasites in mice and hamsters, demonstrating that HbR is a novel vaccine candidate for visceral leishmaniasis [76]. However, it is also intriguing to understand how the heme released from the degradation of the Hb in the parasite lysosomes is trafficked to different intracellular destinations for its use. This pathway is not very well characterized but beginning to be elucidated. It emerges from recent studies that some of the heme transporters possibly regulate this process.
6.4. Heme/iron Transporters in Leishmania
The first heme-binding sites on the promastigote membrane were reported in Leishmania mexicana amazonensis [83]. It assumes to play a role in the transformation of promastigotes to amastigotes as it requires higher oxygen consumption. Analysis of L. amazonensis genome identified another Leishmania iron transporter 1 (LIT1), which is found to be present on the plasma membrane of iron-deprived promastigotes or intracellular amastigotes [84,85]. The LIT1 null mutant parasites show growth defects and are unable to form lesions in the parasite-infected animal model, indicating that this transporter plays an important role in the parasite survival. Subsequently, Leishmania ferric reductase 1 (LFR1) protein was identified, which is also expressed in the parasites in iron-depleted conditions and reduces the iron from the ferric (Fe3+) to the ferrous (Fe2+) state [86]. Moreover, the iron transport function of the LIT1 is dependent on LFR1, and both are overexpressed in amastigotes [87]. In addition, Leishmania infection triggers the production of heme oxygenase in macrophages, which catalyzes the intracellular heme to a ferrous state [88]. It is well established that ligand-bound transferrin receptor (TfR) is internalized via endocytosis and releases Fe3+ in acidified intracellular compartment [89]. Therefore, it is postulated that the Fe3+ form from the transferrin is first reduced to Fe2+ by LFR1, possibly in the acidified compartment in the macrophages, and then it is transported to the parasite’s cytosol by LIT1. Similarly, Nramp1, which is a cation efflux pump, can transport iron from the phagolysosome to the cytosol [90]. Iron from the cytosol is further transported to the appropriate intracellular destination for its use.
Similarly, several heme transporters were also identified in Leishmania. The first heme importer was identified as Leishmania Heme Response 1 (LHR1) in L. mexicana amazonensis based on the homology of C. elegans heme transporter HRG-4 [91]. It promotes the uptake of heme by the parasite in heme-deficient conditions and has proved to be indispensable as null mutant parasites are lethal. They also show severe defects in developing lesions in the parasite-infected mice [92]. Interestingly, LHR1 in L. major (LmHR1) is present in its acidic compartments and colocalizes with internalized Hb, indicating that LmHR1 possibly exports heme generated from Hb degradation in the lysosomes to the cytosol [93]. In addition, another heme transporter, LmFLVCRb, was recently identified in L. major promastigotes based on the homology of mammalian heme transporter [94]. Similar to LHR1, LmFLVCRb was also found to be essential for parasite survival. However, it is puzzling why Leishmania has two heme transporters, and both of them are essential. These open up several questions. Are they species-specific, or are they involved in the transport of the heme to different intracellular organelles? Thus, the heme derived from the Hb is transported to the cytosol by LHR1, which needs to reach the appropriate destination. Based on the role of ABC transporters such as ABCB6, which are involved in the transport of heme to the mitochondria in the mammalian cells, an ABC transporter LABCG5 with unusual topology was identified in Leishmania. This protein is not localized in the endocytic route in Leishmania and therefore may not participate in heme export from lysosomes. However, they can bind to the heme exported by the LHR1 [95]. Finally, another ABC transporter, LABCB3, was identified in Leishmania major, which is involved in the transport of cytosolic heme to the mitochondria and also plays a role in the mitochondrial heme biogenesis, possibly from host precursors [96]. Simultaneously, another mitochondrial iron importer, LMIT1, was identified in L. amazonensis based on the homology of mrs3 (yeast) and mitoferrin-1 (human) gene product. LMIT1 null mutant parasites are not viable due to strong defects in iron content and the function of mitochondria [97]. Thus, recent studies summarize how Leishmania is a heme auxotroph endocytose host hemoprotein and generates intracellular heme for their use (Figure 5).
7. Conclusions
There is now substantial evidence of how Leishmania meets the requirement of the heme through different receptors for hemoproteins and heme transporter. As heme is toxic and not freely available in the host cells, thus these parasites rely on host Hb to acquire heme. The other heme-binding proteins and transporters identified in Leishmania are possibly involved in the trafficking of the heme generated from Hb degradation to the appropriate destination. The most physiologically relevant process is to acquire heme from the host, the Hb, as it is abundantly available in the host. Thus, Leishmania internalizes host Hb through a specific high-affinity receptor (HbR) by clathrin-mediated endocytic process and subsequently transports it to the lysosomes via Rab5 and Rab7 dependent process, where it is degraded to generate intracellular heme. This process appears to be conserved in different species of Leishmania as well as in both promastigote and amastigote forms. The heme released from Hb degradation is then transported to the cytosol by different heme transporters such as LHR1 and LmFLVCRb. Finally, cytosolic heme is transported to the mitochondria by mitochondrial import proteins such as LABCB3 in Leishmania. This process of acquiring heme from the host Hb is also supported by the fact that visceral leishmaniasis patients are anemic and show increased erythrophagocytosis. In addition, the discovery of HbR as hexokinase opens up several interesting areas for future studies. Furthermore, apart from the fact that HbR is a novel vaccine candidate against leishmaniasis, it is also worth exploring Rab5b null mutant parasites as a vaccine candidate. Similarly, evaluation of the mimicking peptide corresponding to the Hb-binding domain is another area to explore for novel drug design against leishmaniasis. Thus, understanding the mechanism of Hb endocytosis and heme trafficking in Leishmania identifies several novel targets, which can be exploited for the development of therapeutic strategies against leishmaniasis.
Author Contributions
Conceptualization, supervision, review and editing, A.M., I.A. and R.B. Writing initial draft and editing, I.A. and R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available in the cited references.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Ld | Leishmania donovani; |
| Hb | Hemoglobin; |
| HbR | Hb-Receptor; |
| VL | Visceral Leishmaniasis; |
| CME | Clathrin mediated endocytosis. |
References
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Chappuis, F.; Sundar, S.; Hailu, A.; Ghalib, H.; Rijal, S.; Peeling, R.W.; Alvar, J.; Boelaert, M. Visceral leishmaniasis: What are the needs for diagnosis, treatment and control? Nat. Rev. Microbiol. 2007, 5, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.D.; Sousa, A.D.Q. Clinical Spectrum of Leishmaniasis. Clin. Infect. Dis. 1996, 22, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kedzierski, L. Leishmaniasis vaccine: Where are we today? J. Glob. Infect. Dis. 2010, 2, 177–185. [Google Scholar] [CrossRef] [PubMed]
- CBern, C.; Adler-Moore, J.; Berenguer, J.; Boelaert, M.; den Boer, M.; Davidson, R.N.; Figueras, C.; Gradoni, L.; Kafetzis, D.A.; Ritmeijer, K.; et al. Liposomal amphotericin B for the treatment of visceral leishmaniasis. Clin. Infect. Dis. 2006, 43, 917–924. [Google Scholar]
- Berman, J. Miltefosine to treat leishmaniasis. Expert Opin. Pharmacother. 2005, 6, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Boer, M.D.; Davidson, R.N. Treatment options for visceral leishmaniasis. Expert Rev. Anti-Infect. Ther. 2006, 4, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; More, D.K.; Singh, M.K.; Singh, V.P.; Sharma, S.; Makharia, A.; Kumar, P.C.K.; Murray, H.W. Failure of Pentavalent Antimony in Visceral Leishmaniasis in India: Report from the Center of the Indian Epidemic. Clin. Infect. Dis. 2000, 31, 1104–1107. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Victoria, F.J.; Sánchez-Cañete, M.P.; Seifert, K.; Croft, S.L.; Sundar, S.; Castanys, S.; Gamarro, F. Mechanisms of experimental resistance of Leishmania to miltefosine: Implications for clinical use. Drug Resist. Updat. 2006, 9, 26–39. [Google Scholar] [CrossRef]
- Zutshi, S.; Kumar, S.; Chauhan, P.; Bansode, Y.; Nair, A.; Roy, S.; Sarkar, A.; Saha, B. Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments. Vaccines 2019, 7, 156. [Google Scholar] [CrossRef] [Green Version]
- Tripodi, K.E.J.; Bravo, S.M.M.; Cricco, J.A. Role of Heme and Heme-Proteins in Trypanosomatid Essential Metabolic Pathways. Enzym. Res. 2011, 2011, 873230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dailey, H.A.; Dailey, T.A.; Gerdes, S.; Jahn, D.; Jahn, M.; O’Brian, M.R.; Warren, M.J. Prokaryotic Heme Biosynthesis: Multiple Pathways to a Common Essential Product. Microbiol. Mol. Biol. Rev. 2017, 81, e00048-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sah, J.; Ito, H.; Kolli, B.K.; Peterson, D.A.; Sassa, S.; Chang, K.-P. Genetic Rescue of Leishmania Deficiency in Porphyrin Biosynthesis Creates Mutants Suitable for Analysis of Cellular Events in Uroporphyria and for Photodynamic Therapy. J. Biol. Chem. 2002, 277, 14902–14909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kořený, L.; Lukes, J.; Oborník, M. Evolution of the haem synthetic pathway in kinetoplastid flagellates: An essential pathway that is not essential after all? Int. J. Parasitol. 2010, 40, 149–156. [Google Scholar] [CrossRef]
- Chang, K.P.; Chang, C.S.; Sassa, S. Heme biosynthesis in bacterium-protozoon symbioses: Enzymic defects in host hemoflagellates and complemental role of their intracellular symbiotes. Proc. Natl. Acad. Sci. USA 1975, 72, 2979–2983. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.S.; Chang, K.-P. Heme requirement and acquisition by extracellular and intracellular stages of Leishmania mexicana amazonensis. Mol. Biochem. Parasitol. 1985, 16, 267–276. [Google Scholar] [CrossRef]
- Solar, I.; Muller-Eberhard, U.; Shviro, Y.; Shaklai, N. Long-term intercalation of residual hemin in erythrocyte membranes distorts the cell. Biochim. Biophys. Acta 1991, 1062, 51–58. [Google Scholar] [CrossRef]
- Ong, S.T.; Shan Ho, J.Z.; Ho, B.; Ding, J.L. Iron-withholding strategy in innate immunity. Immunobiology 2006, 211, 295–314. [Google Scholar] [CrossRef]
- Anderson, G.J.; Vulpe, C.D. Mammalian iron transport. Cell. Mol. Life Sci. 2009, 66, 3241–3261. [Google Scholar] [CrossRef]
- Kristiansen, M.; Graversen, J.H.; Jacobsen, C.; Sonne, O.; Hoffman, H.-J.; Law, S.K.A.; Moestrup, S.K. Identification of the haemoglobin scavenger receptor. Nature 2001, 409, 198–201. [Google Scholar] [CrossRef]
- Wandersman, C.; Delepelaire, P. Bacterial Iron Sources: From Siderophores to Hemophores. Annu. Rev. Microbiol. 2004, 58, 611–647. [Google Scholar] [CrossRef] [PubMed]
- Contreras, H.; Chim, N.; Credali, A.; Goulding, C.W. Heme uptake in bacterial pathogens. Curr. Opin. Chem. Biol. 2014, 19, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornitzer, D.; Roy, U. Pathways of heme utilization in fungi. Biochim. Biophys. Acta 2020, 1867, 118817. [Google Scholar] [CrossRef] [PubMed]
- Knutson, M.D.; Oukka, M.; Koss, L.M.; Aydemir, F.; Wessling-Resnick, M. Iron release from macrophages after erythrophagocytosis is up-regulated by ferroportin 1 overexpression and down-regulated by hepcidin. Proc. Natl. Acad. Sci. USA 2005, 102, 1324–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, A.; Uchida, K.; Chambers, J.; Sato, K.; Hong, J.; Sanjoba, C.; Matsumoto, Y.; Yamagishi, J.; Goto, Y. Hemophagocytosis induced by Leishmania donovani infection is beneficial to parasite survival within macrophages. PLOS Neglected Trop. Dis. 2019, 13, e0007816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamhawi, S. Phlebotomine sand flies and Leishmania parasites: Friends or foes? Trends Parasitol. 2006, 22, 439–445. [Google Scholar] [CrossRef]
- Roy, J.C. Cultivation of various Leishmania parasites in solid medium. Ind. J. Med. Res. 1932, 20, 367. [Google Scholar]
- Gholamhosseinian, A.; Vassef, A. Superiority of Hemoglobin to Hemin for Cultivation of Leishmania tropica Promastigotes in Serum-Free Media. J. Protozool. 1988, 35, 446–449. [Google Scholar] [CrossRef]
- Ide, A.R.; da Silva, B.A.; Santos, A.L.d.; Vermelho, A.B.; Alviano, C.S.; Dutra, P.M.; Mdo, S.R. A new experimental culture medium for cultivation of Leishmania amazonensis: Its efficacy for the continuous in vitro growth and differentiation of infective promastigote forms. Parasitol. Res. 2010, 106, 1249–1252. [Google Scholar]
- Elkin, S.R.; Lakoduk, A.; Schmid, S.L. Endocytic pathways and endosomal trafficking: A primer. Wien. Med. Wochenschr. 2016, 166, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.R. Rab GTPases: Specifying and deciphering organelle identity and function. Trends Cell Biol. 2001, 11, 487–491. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Barbieri, A.M.; Funato, K.; Roberts, R.; Stahl, P.D. Sequential Actions of Rab5 and Rab7 Regulate Endocytosis in the Xenopus Oocyte. J. Cell Biol. 1997, 136, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Sönnichsen, B.; De Renzis, S.; Nielsen, E.; Rietdorf, J.; Zerial, M. Distinct Membrane Domains on Endosomes in the Recycling Pathway Visualized by Multicolor Imaging of Rab4, Rab5, and Rab11. J. Cell Biol. 2000, 149, 901–914. [Google Scholar] [CrossRef]
- Bucci, C.; Thomsen, P.; Nicoziani, P.; McCarthy, J.; van Deurs, B. Rab7: A Key to Lysosome Biogenesis. Mol. Biol. Cell 2000, 11, 467–480. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Funato, K.; Stahl, P.D. Rab7 Regulates Transport from Early to Late Endocytic Compartments in Xenopus Oocytes. J. Biol. Chem. 1997, 272, 13055–13059. [Google Scholar] [CrossRef] [Green Version]
- Sorkin, A.; von Zastrow, M. Endocytosis and signalling: Intertwining molecular networks. Nat. Rev. Mol. Cell Biol. 2009, 10, 609–622. [Google Scholar] [CrossRef] [Green Version]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef]
- Cai, H.; Reinisch, K.; Ferro-Novick, S. Coats, Tethers, Rabs, and SNAREs Work Together to Mediate the Intracellular Destination of a Transport Vesicle. Dev. Cell 2007, 12, 671–682. [Google Scholar] [CrossRef] [Green Version]
- Jahn, R.; Scheller, R.H. SNAREs—Engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef]
- Hong, W.; Lev, S. Tethering the assembly of SNARE complexes. Trends Cell Biol. 2013, 24, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Murungi, E.; Barlow, L.D.; Venkatesh, D.; Adung, V.O.; Dacks, J.B.; Field, M.C.; Christoffels, A. A comparative analysis of trypanosomatid SNARE proteins. Parasitol. Int. 2013, 63, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesh, D.; Zhang, N.; Zoltner, M.; Del Pino, R.C.; Field, M.C. Evolution of protein trafficking in kinetoplastid parasites: Complexity and pathogenesis. Traffic 2018, 19, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M. The structure, biosynthesis and functions of glycosylphosphatidylinositol anchors, and the contributions of trypanosome research. J. Cell Sci. 1999, 112, 2799–2809. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.J.; Ferguson, M. The structure, biosynthesis and function of glycosylated phosphatidylinositols in the parasitic protozoa and higher eukaryotes. Biochem. J. 1993, 294, 305–324. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.S.; Chang, K.P. Monoclonal antibody affinity purification of a Leishmania membrane glycoprotein and its inhibition of leishmania-macrophage binding. Proc. Natl. Acad. Sci. USA 1986, 83, 100–104. [Google Scholar] [CrossRef] [Green Version]
- JMcKerrow, H.; Sun, E.; Rosenthal, P.J.; Bouvier, J. The proteases and pathogenicity of parasitic protozoa. Annu. Rev. Microbiol. 1993, 47, 821–853. [Google Scholar] [CrossRef]
- Chaudhuri, G.; Chaudhuri, M.; Pan, A.; Chang, K.P. Surface acid proteinase (gp63) of Leishmania mexicana. A metalloenzyme capable of protecting liposome-encapsulated proteins from phagolysosomal degradation by macrophages. J. Biol. Chem. 1989, 264, 7483–7489. [Google Scholar] [CrossRef]
- McGwire, B.; Chang, K.-P. Genetic rescue of surface metalloproteinase (gp63)-deficiency in Leishmania amazonensis variants increases their infection of macrophages at the early phase. Mol. Biochem. Parasitol. 1994, 66, 345–347. [Google Scholar] [CrossRef]
- Agarwal, S.; Rastogi, R.; Gupta, D.; Patel, N.; Raje, M.; Mukhopadhyay, A. Clathrin-mediated hemoglobin endocytosis is essential for survival of Leishmania. Biochim. Biophys. Acta 2013, 1833, 1065–1077. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.B.; Tandon, R.; Krishnamurthy, G.; Vikram, R.; Sharma, N.; Basu, S.K.; Mukhopadhyay, A. Rab5-mediated endosome-endosome fusion regulates hemoglobin endocytosis in Leishmania donovani. EMBO J. 2003, 22, 5712–5722. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, R.; Verma, J.K.; Kapoor, A.; Langsley, G.; Mukhopadhyay, A. Rab5 Isoforms Specifically Regulate Different Modes of Endocytosis in Leishmania. J. Biol. Chem. 2016, 291, 14732–14746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, R.; Kapoor, A.; Verma, J.K.; Ansari, I.; Sood, C.; Kumar, K.; Mukhopadhyay, A. Rab5b function is essential to acquire heme from hemoglobin endocytosis for survival of Leishmania. Biochim. Biophys. Acta 2020, 1868, 118868. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Singh, S.B.; Basu, S.K.; Mukhopadhyay, A. Leishmania requires Rab7-mediated degradation of endocytosed hemoglobin for their growth. Proc. Natl. Acad. Sci. USA 2008, 105, 3980–3985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahl, S.; Parashar, S.; Malhotra, H.; Raje, M.; Mukhopadhyay, A. Functional Characterization of Monomeric GTPase Rab1 in the Secretory Pathway of Leishmania. J. Biol. Chem. 2015, 290, 29993–30005. [Google Scholar] [CrossRef] [Green Version]
- Parashar, S.; Mukhopadhyay, A. GTPase Sar1 regulates the trafficking and secretion of the virulence factor gp63 in Leishmania. J. Biol. Chem. 2017, 292, 12111–12125. [Google Scholar] [CrossRef] [Green Version]
- Besteiro, S.; Coombs, G.H.; Mottram, J.C. The SNARE protein family of Leishmania major. BMC Genom. 2006, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Landfear, S.M. Nutrient Transport and Pathogenesis in Selected Parasitic Protozoa. Eukaryot. Cell 2011, 10, 483–493. [Google Scholar] [CrossRef] [Green Version]
- De Cicco, N.N.; Pereira, M.G.; Corrêa, J.R.; Andrade-Neto, V.V.; Saraiva, F.B.; Chagas-Lima, A.C.; Gondim, K.C.; Torres-Santos, E.C.; Folly, E.; Saraiva, E.M.; et al. LDL uptake by Leishmania amazonensis: Involvement of membrane lipid microdomains. Exp. Parasitol. 2012, 130, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, G.; Carter, N.S.; Drew, M.E.; Beverley, S.M.; Sanchez, M.A.; Seyfang, A.; Ullman, B.; Landfear, S.M. Cloning of Leishmania nucleoside transporter genes by rescue of a transport-deficient mutant. Proc. Natl. Acad. Sci. USA 1998, 95, 9873–9878. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, G.; Ullman, B.; Landfear, S.M. Point mutations in a nucleoside transporter gene from Leishmania donovani confer drug resistance and alter substrate selectivity. Proc. Natl. Acad. Sci. USA 2001, 98, 6092–6097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, D.; Sanchez, M.A.; Koch, H.P.; Larsson, H.P.; Landfear, S.M. An Acid-activated Nucleobase Transporter from Leishmania major. J. Biol. Chem. 2009, 284, 16164–16169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, D.; Kündig, C.; Ouellette, M. A New Type of High Affinity Folic Acid Transporter in the Protozoan Parasite Leishmania and Deletion of Its Gene in Methotrexate-resistant Cells. J. Biol. Chem. 2002, 277, 29460–29467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkouvatsos, K.; Papanikolaou, G.; Pantopoulos, K. Regulation of iron transport and the role of transferrin. Biochim. Biophys. Acta 2012, 1820, 188–202. [Google Scholar] [CrossRef]
- Voyiatzaki, C.S.; Soteriadou, K.P. Evidence of transferrin binding sites on the surface of Leishmania promastigotes. J. Biol. Chem. 1990, 265, 22380–22385. [Google Scholar] [CrossRef]
- Voyiatzaki, C.; Soteriadou, K. Identification and isolation of the Leishmania transferrin receptor. J. Biol. Chem. 1992, 267, 9112–9117. [Google Scholar] [CrossRef]
- Russell, D.; Xu, S.; Chakraborty, P. Intracellular trafficking and the parasitophorous vacuole of Leishmania mexicana-infected macrophages. J. Cell Sci. 1992, 103, 1193–1210. [Google Scholar] [CrossRef]
- Borges, V.M.; Vannier-Santos, M.A.; De Souza, W. Subverted transferrin trafficking in Leishmania—Infected macrophages. Parasitol. Res. 1998, 84, 811–822. [Google Scholar] [CrossRef]
- Rawat, P.; Kumar, S.; Sheokand, N.; Raje, C.I.; Raje, M. The multifunctional glycolytic protein glyceraldehyde-3-phosphate dehydrogenase (GAPDH) is a novel macrophage lactoferrin receptor. Biochem. Cell Biol. 2012, 90, 329–338. [Google Scholar] [CrossRef]
- Raje, C.I.; Kumar, S.; Harle, A.; Nanda, J.S.; Raje, M. The Macrophage Cell Surface Glyceraldehyde-3-phosphate Dehydrogenase Is a Novel Transferrin Receptor. J. Biol. Chem. 2007, 282, 3252–3261. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.E.; Vorhies, R.W.; Andersen, K.A.; Britigan, B.E. Acquisition of iron from transferrin and lactoferrin by the protozoan Leishmania chagasi. Infect. Immun. 1994, 62, 3262–3269. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.E.; Lewis, T.S.; Miller, M.A.; McCormick, M.L.; Britigan, B.E. Leishmania chagasi: Uptake of iron bound to lactoferrin or transferrin requires an iron reductase. Exp. Parasitol. 2002, 100, 196–207. [Google Scholar] [CrossRef]
- Pantopoulos, K.; Porwal, S.K.; Tartakoff, A.; Devireddy, L. Mechanisms of Mammalian Iron Homeostasis. Biochemistry 2012, 51, 5705–5724. [Google Scholar] [CrossRef]
- Sengupta, S.; Tripathi, J.; Tandon, R.; Raje, M.; Roy, R.P.; Basu, S.K.; Mukhopadhyay, A. Hemoglobin Endocytosis in Leishmania Is Mediated through a 46-kDa Protein Located in the Flagellar Pocket. J. Biol. Chem. 1999, 274, 2758–2765. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, G.; Vikram, R.; Singh, S.B.; Patel, N.; Agarwal, S.; Mukhopadhyay, G.; Basu, S.K.; Mukhopadhyay, A. Hemoglobin Receptor in Leishmania Is a Hexokinase Located in the Flagellar Pocket. J. Biol. Chem. 2005, 280, 5884–5891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, R.; Gupta, D.; Rastogi, R.; Vikram, R.; Krishnamurthy, G.; Bimal, S.; Roy, S.; Mukhopadhyay, A. Vaccination with Leishmania Hemoglobin Receptor–Encoding DNA Protects Against Visceral Leishmaniasis. Sci. Transl. Med. 2013, 5, 202ra121. [Google Scholar] [CrossRef]
- Carvalho, S.; Cruz, T.; Santarém, N.; Castro, H.; Costa, V.; Tomás, A.M. Heme as a source of iron to Leishmania infantum amastigotes. Acta Trop. 2009, 109, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.; Verma, J.K.; Singh, V.; Krishnamurthy, G.; Sood, C.; Kapoor, A.; Kumar, K.; Ansari, I.; Mukhopadhyay, A. Identification and characterization of the hemoglobin-binding domain of hemoglobin receptor in Leishmania. FEBS Lett. 2020, 595, 548–558. [Google Scholar] [CrossRef]
- Kaksonen, M.; Roux, A. Mechanisms of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2018, 19, 313–326. [Google Scholar] [CrossRef]
- Mayor, S.; Pagano, R.E. Pathways of clathrin-independent endocytosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 603–612. [Google Scholar] [CrossRef]
- Mettlen, M.; Chen, P.-H.; Srinivasan, S.; Danuser, G.; Schmid, S.L. Regulation of Clathrin-Mediated Endocytosis. Annu. Rev. Biochem. 2018, 87, 871–896. [Google Scholar] [CrossRef] [PubMed]
- Morlot, S.; Roux, A. Mechanics of Dynamin-Mediated Membrane Fission. Annu. Rev. Biophys. 2013, 42, 629–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galbraith, R.; Mcelrath, M. Heme binding to Leishmania mexicana amazonensis. Mol. Biochem. Parasitol. 1988, 29, 47–53. [Google Scholar] [CrossRef]
- Huynh, C.; Sacks, D.L.; Andrews, N.W. A Leishmania amazonensis ZIP family iron transporter is essential for parasite replication within macrophage phagolysosomes. J. Exp. Med. 2006, 203, 2363–2375. [Google Scholar] [CrossRef]
- Jacques, I.; Andrews, N.W.; Huynh, C. Functional characterization of LIT1, the Leishmania amazonensis ferrous iron transporter. Mol. Biochem. Parasitol. 2010, 170, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flannery, A.R.; Huynh, C.; Mittra, B.; Mortara, R.; Andrews, N. LFR1 Ferric Iron Reductase of Leishmania amazonensis Is Essential for the Generation of Infective Parasite Forms. J. Biol. Chem. 2011, 286, 23266–23279. [Google Scholar] [CrossRef] [Green Version]
- Mittra, B.; Andrews, N.W. IRONy OF FATE: Role of iron-mediated ROS in Leishmania differentiation. Trends Parasitol. 2013, 29, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Luz, N.F.; Andrade, B.B.; Feijó, D.F.; Araújo-Santos, T.; Carvalho, G.Q.; Andrade, D.; Abánades, D.R.; Melo, E.V.; Silva, A.M.; Brodskyn, C.I.; et al. Heme Oxygenase-1 Promotes the Persistence of Leishmania chagasi Infection. J. Immunol. 2012, 188, 4460–4467. [Google Scholar] [CrossRef] [Green Version]
- Andrews, N.C. Iron homeostasis: Insights from genetics and animal models. Nat. Rev. Genet. 2000, 1, 208–217. [Google Scholar] [CrossRef]
- Blackwell, J.M.; Goswami, T.; Evans, C.A.W.; Sibthorpe, D.; Papo, N.; White, J.K.; Searle, S.; Miller, E.N.; Peacock, C.S.; Mohammed, H.; et al. SLC11A1 (formerly NRAMP1) and disease resistance. Microreview. Cell. Microbiol. 2001, 3, 773–784. [Google Scholar] [CrossRef]
- Huynh, C.; Yuan, X.; Miguel, D.C.; Renberg, R.L.; Protchenko, O.; Philpott, C.C.; Hamza, I.; Andrews, N.W. Heme Uptake by Leishmania amazonensis Is Mediated by the Transmembrane Protein LHR1. PLOS Pathog. 2012, 8, e1002795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miguel, D.C.; Flannery, A.R.; Mittra, B.; Andrews, N.W.; Barua, S.; Iyer, J.K.; Larabee, J.L.; Raisley, B.; Hughes, M.A.; Coggeshall, K.M.; et al. Heme Uptake Mediated by LHR1 Is Essential for Leishmania amazonensis Virulence. Infect. Immun. 2013, 81, 3620–3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabello-Donayre, M.; Malagarie-Cazenave, S.; Campos-Salinas, J.; Gálvez, F.J.; Rodríguez-Martínez, A.; Pineda-Molina, E.; Orrego, L.M.; García, M.M.; Sánchez-Cañete, M.P.; Estevez, A.; et al. Trypanosomatid parasites rescue heme from endocytosed hemoglobin through lysosomal HRG transporters. Mol. Microbiol. 2016, 101, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Cabello-Donayre, M.; Orrego, L.M.; Herráez, E.; Vargas, P.; Martínez-García, M.; Campos-Salinas, J.; Pérez-Victoria, I.; Vicente, B.; Marín, J.J.G.; Pérez-Victoria, J.M. Leishmania heme uptake involves LmFLVCRb, a novel porphyrin transporter essential for the parasite. Cell. Mol. Life Sci. 2019, 77, 1827–1845. [Google Scholar] [CrossRef] [PubMed]
- Campos-Salinas, J.; Cabello-Donayre, M.; García-Hernández, R.; Pérez-Victoria, I.; Castanys, S.; Gamarro, F.; Pérez-Victoria, J.M. A new ATP-binding cassette protein is involved in intracellular haem trafficking in Leishmania. Mol. Microbiol. 2011, 79, 1430–1444. [Google Scholar] [CrossRef] [PubMed]
- García, M.M.; Campos-Salinas, J.; Cabello-Donayre, M.; Pineda-Molina, E.; Gálvez, F.J.; Orrego, L.M.; Sánchez-Cañete, M.P.; Malagarie-Cazenave, S.; Koeller, D.M.; Pérez-Victoria, J.M. LmABCB3, an atypical mitochondrial ABC transporter essential for Leishmania major virulence, acts in heme and cytosolic iron/sulfur clusters biogenesis. Parasites Vectors 2016, 9, 7. [Google Scholar]
- Mittra, B.; Laranjeira-Silva, M.F.; De Menezes, J.P.B.; Jensen, J.; Michailowsky, V.; Andrews, N.W. A Trypanosomatid Iron Transporter that Regulates Mitochondrial Function Is Required for Leishmania amazonensis Virulence. PLoS Pathog. 2016, 12, e1005340. [Google Scholar] [CrossRef]
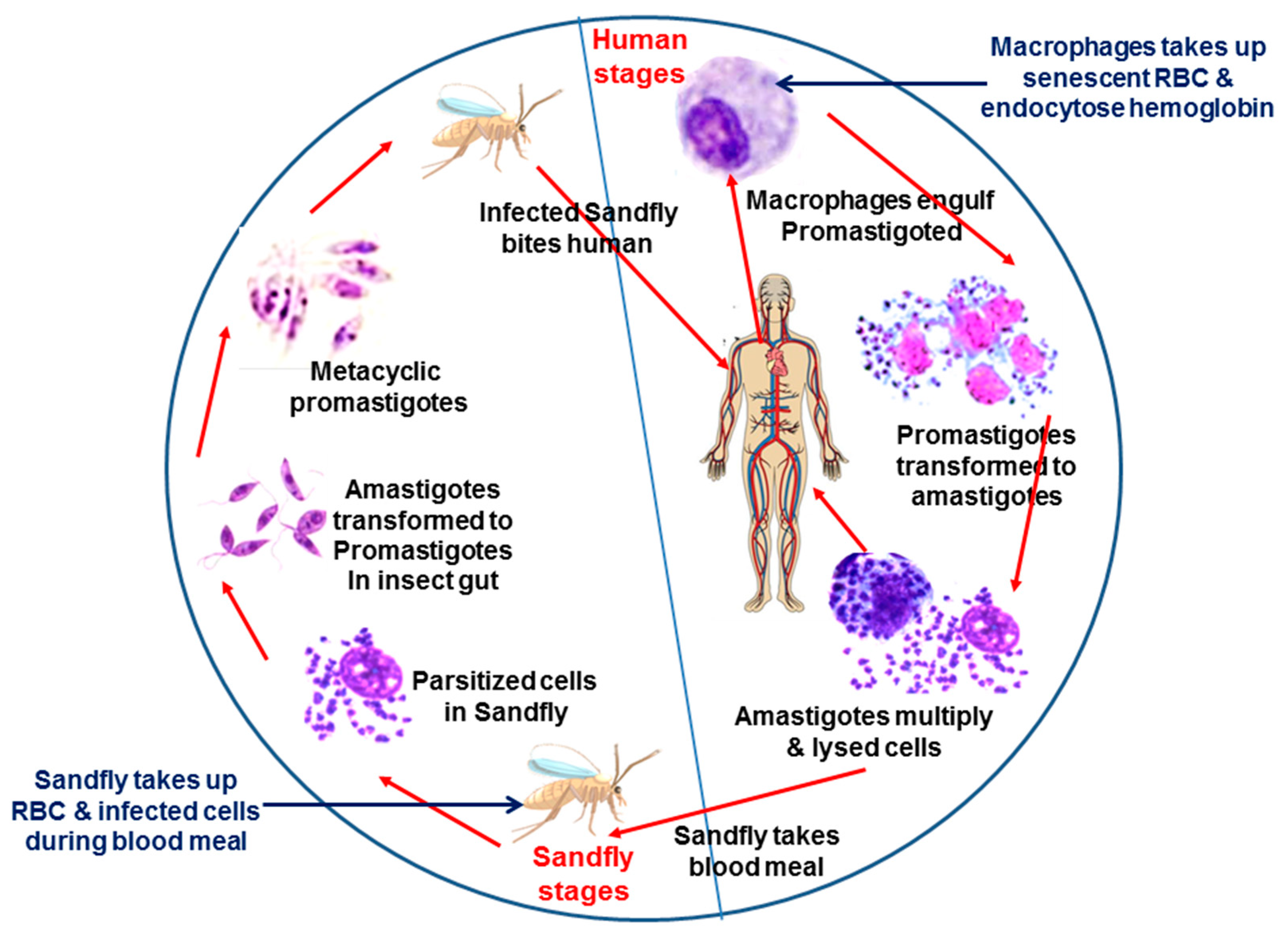
Figure 1.
Schematic representation shows the life cycle of Leishmania and source of hemoglobin.Leishmania has a digenetic life cycle. Transmission to humans occurs through the bite of a female Phlebotomine sandfly. In humans, promastigotes are taken up by the macrophages and transform into amastigotes in the parasitophorous vacuole (PV). Macrophages also ingest senescent RBCs, and lysed RBCs serve as source of hemoglobin for Leishmania. Amastigotes multiply inside the macrophages and eventually lyse cells. Parasites are released and further infect the surrounding macrophages leading to the manifestation of disease. Parasitized cells are ingested by the sandflies during the blood meal. Inside the midgut of the sandfly, amastigote transforms into motile procyclic promastigotes and colonizes their digestive tract. The insect also takes RBCs during the blood meal, which serves as a source of hemoglobin. Finally, they differentiate into infective metacyclic promastigote form and remain in the saliva of the sandfly. Parasites are transmitted to a new vertebrate host during their next blood meal.
Figure 1.
Schematic representation shows the life cycle of Leishmania and source of hemoglobin.Leishmania has a digenetic life cycle. Transmission to humans occurs through the bite of a female Phlebotomine sandfly. In humans, promastigotes are taken up by the macrophages and transform into amastigotes in the parasitophorous vacuole (PV). Macrophages also ingest senescent RBCs, and lysed RBCs serve as source of hemoglobin for Leishmania. Amastigotes multiply inside the macrophages and eventually lyse cells. Parasites are released and further infect the surrounding macrophages leading to the manifestation of disease. Parasitized cells are ingested by the sandflies during the blood meal. Inside the midgut of the sandfly, amastigote transforms into motile procyclic promastigotes and colonizes their digestive tract. The insect also takes RBCs during the blood meal, which serves as a source of hemoglobin. Finally, they differentiate into infective metacyclic promastigote form and remain in the saliva of the sandfly. Parasites are transmitted to a new vertebrate host during their next blood meal.

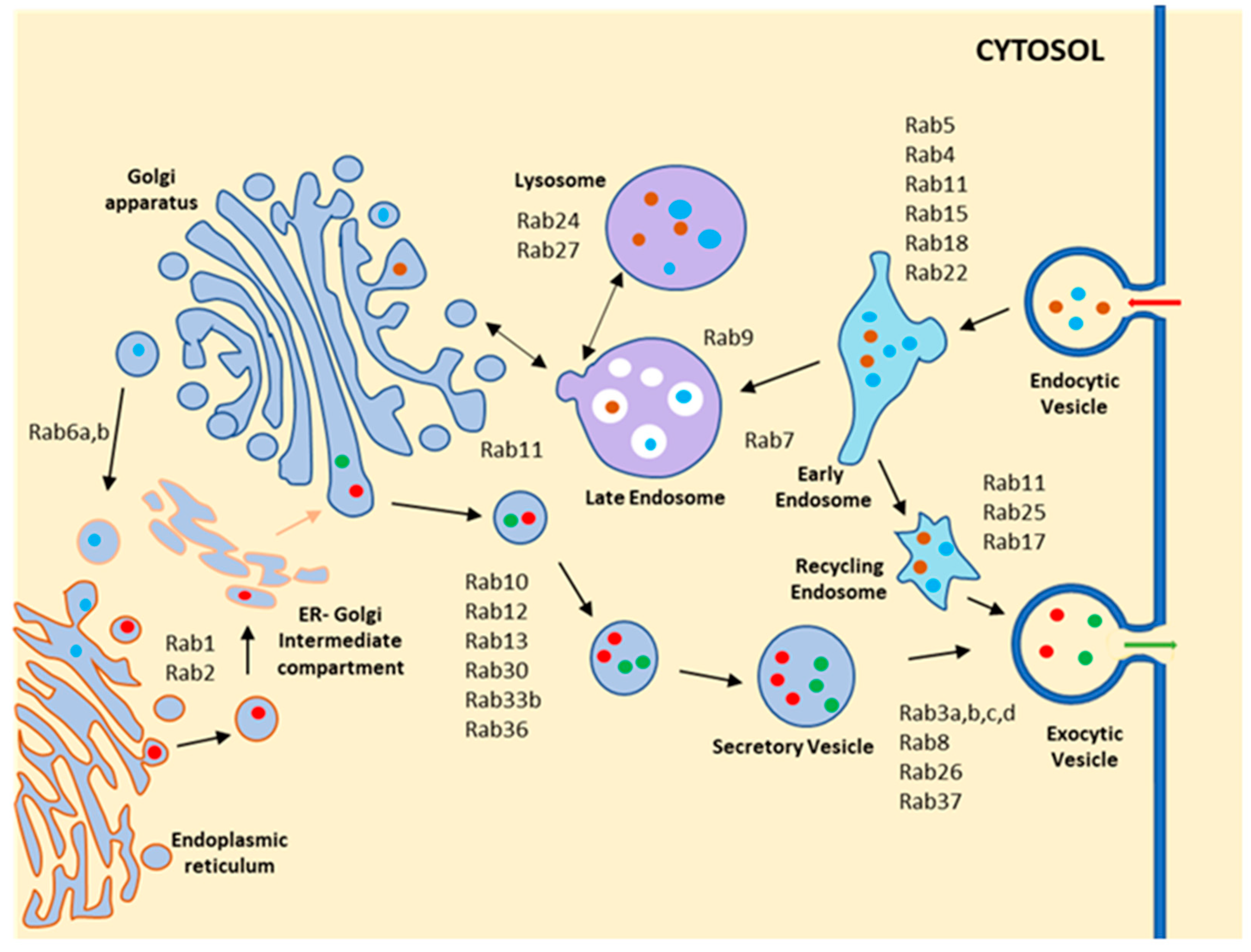
Figure 2.
Schematic diagram shows the localization and function of different Rab GTPases in mammalian cells. Rab GTPases are present in distinct membranous compartments and regulate transport of cargo between the various compartments. In the endocytic pathway, Rab5 involves trafficking of the cargo from the plasma membrane to the early endosome and localizes on the early endocytic compartment. Rab4 participates in the fast-recycling pathway from the early endosome, while Rab11 and Rab35 regulate the slow recycling pathway. Rab7 helps in the trafficking of cargo to the lysosome from the early endosome, and Rab9 regulates cargo transport from the lysosome to the TGN. In the secretory pathway, Rab1 localizes on the ER Golgi intermediate compartment and regulates anterograde trafficking from ER to Golgi, whereas Rab2 involves trafficking from Golgi to ER in retrograde. Rab6 controls intra-Golgi trafficking. Rab3, Rab27, Rab8, and Rab37 regulate the transport of different secretory vesicles via the exocytic pathway.
Figure 2.
Schematic diagram shows the localization and function of different Rab GTPases in mammalian cells. Rab GTPases are present in distinct membranous compartments and regulate transport of cargo between the various compartments. In the endocytic pathway, Rab5 involves trafficking of the cargo from the plasma membrane to the early endosome and localizes on the early endocytic compartment. Rab4 participates in the fast-recycling pathway from the early endosome, while Rab11 and Rab35 regulate the slow recycling pathway. Rab7 helps in the trafficking of cargo to the lysosome from the early endosome, and Rab9 regulates cargo transport from the lysosome to the TGN. In the secretory pathway, Rab1 localizes on the ER Golgi intermediate compartment and regulates anterograde trafficking from ER to Golgi, whereas Rab2 involves trafficking from Golgi to ER in retrograde. Rab6 controls intra-Golgi trafficking. Rab3, Rab27, Rab8, and Rab37 regulate the transport of different secretory vesicles via the exocytic pathway.

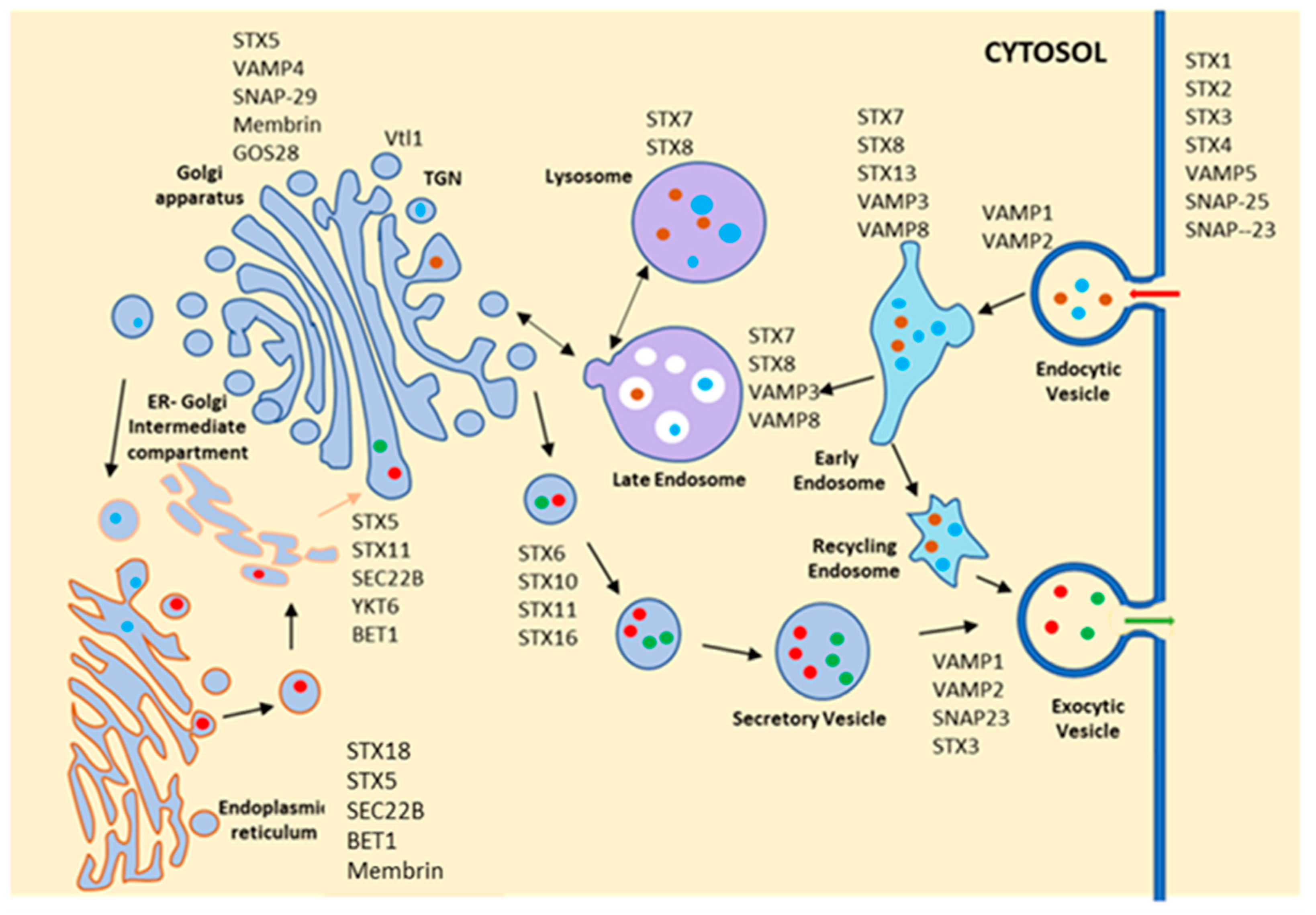
Figure 3.
Schematic diagram shows the localization and function of different SNAREs in mammalian cells. Similar to Rab GTPases, SNAREs are also localized on selective compartments and provide specificity in vesicle fusion steps. They are subdivided into v-SNARE and t-SNAREs highlighted as VAMP and STX, respectively, in the figure. STX (syntaxin) 1, STX2, STX3, STX4, VAMP5, SNAP-25 and SNAP-23 are localized at the plasma membrane and possibly regulate the fusion of endocytic as well as secretory vesicles. VAMP1, VAMP2 and VAMP3, along with STX7, STX8 and STX13 are found in different endocytic compartments and regulate the different steps in trafficking of endocytic cargo from plasma membrane to lysosomes. STX11, STX5 and SEC28b are located at the Golgi apparatus, whereas STX18, Sec22b, BET1 and Membrin are found at the endoplasmic reticulum, suggesting that traffic between ER and Golgi is regulated by these proteins along with VAMP4 and SNAP29. STX6, STX10, STX11 and STX16, along with VAMP1, VAMP2 and SNAP3, regulate the transport of different secretory vesicles from the Golgi.
Figure 3.
Schematic diagram shows the localization and function of different SNAREs in mammalian cells. Similar to Rab GTPases, SNAREs are also localized on selective compartments and provide specificity in vesicle fusion steps. They are subdivided into v-SNARE and t-SNAREs highlighted as VAMP and STX, respectively, in the figure. STX (syntaxin) 1, STX2, STX3, STX4, VAMP5, SNAP-25 and SNAP-23 are localized at the plasma membrane and possibly regulate the fusion of endocytic as well as secretory vesicles. VAMP1, VAMP2 and VAMP3, along with STX7, STX8 and STX13 are found in different endocytic compartments and regulate the different steps in trafficking of endocytic cargo from plasma membrane to lysosomes. STX11, STX5 and SEC28b are located at the Golgi apparatus, whereas STX18, Sec22b, BET1 and Membrin are found at the endoplasmic reticulum, suggesting that traffic between ER and Golgi is regulated by these proteins along with VAMP4 and SNAP29. STX6, STX10, STX11 and STX16, along with VAMP1, VAMP2 and SNAP3, regulate the transport of different secretory vesicles from the Golgi.

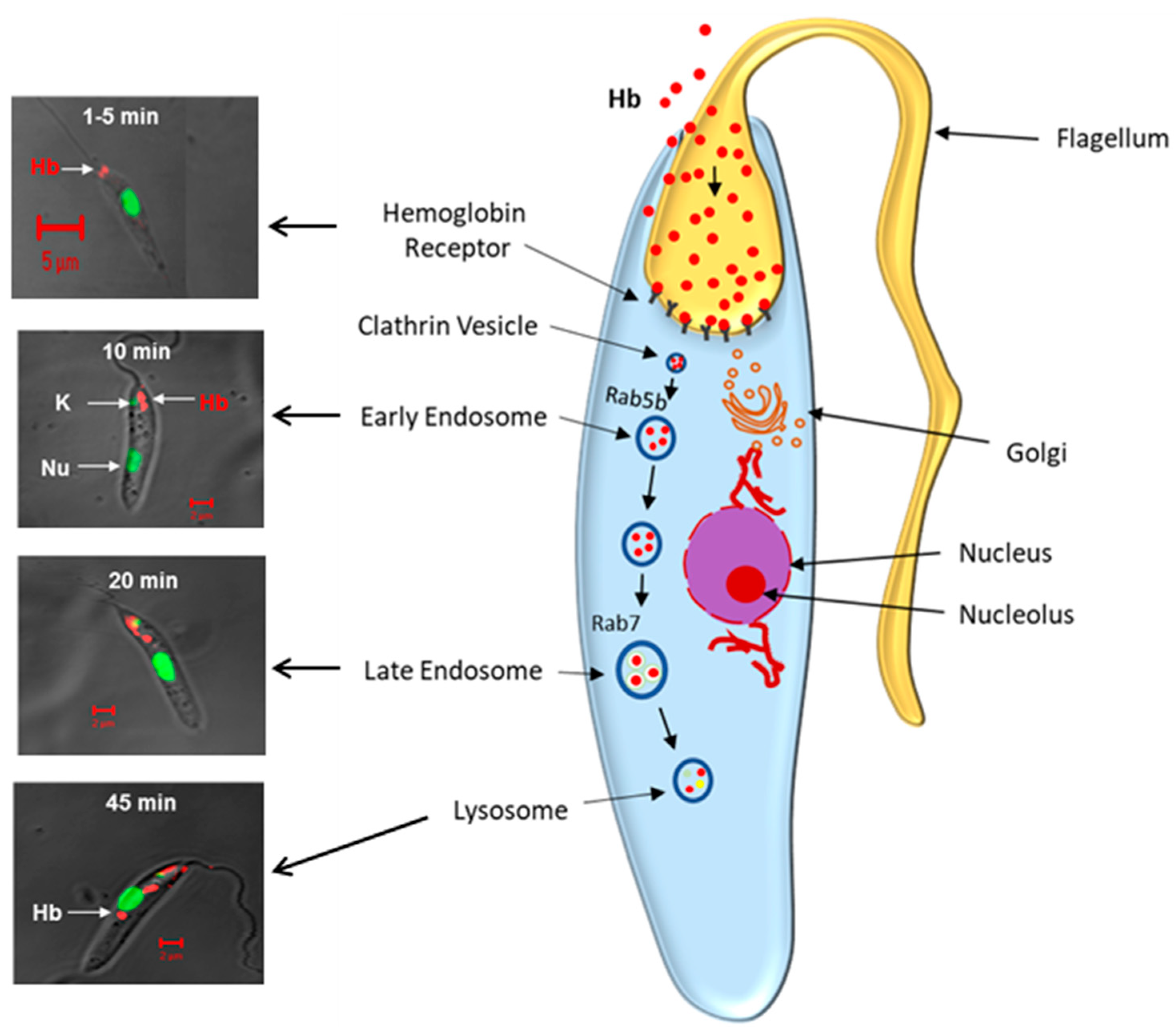
Figure 4.
Schematic representation demonstrates hemoglobin endocytosis in Leishmania donovani promastigotes. (Left Panel) shows that Alexa-Red labeled Hb bind to the Hb receptor on the flagellar pocket of Leishmania, which traffics the Rab5 positive early endosome by 10 min and finally reaches the lysosome via late endosome in about 45 min. Nucleus is marked with green. (Right Panel) depicts the schematic diagram of Hb trafficking from the flagellar pocket to the lysosomes via early and late endocytic compartments in Rab5b and Rab7 dependent processes in Leishmania promastigotes.
Figure 4.
Schematic representation demonstrates hemoglobin endocytosis in Leishmania donovani promastigotes. (Left Panel) shows that Alexa-Red labeled Hb bind to the Hb receptor on the flagellar pocket of Leishmania, which traffics the Rab5 positive early endosome by 10 min and finally reaches the lysosome via late endosome in about 45 min. Nucleus is marked with green. (Right Panel) depicts the schematic diagram of Hb trafficking from the flagellar pocket to the lysosomes via early and late endocytic compartments in Rab5b and Rab7 dependent processes in Leishmania promastigotes.

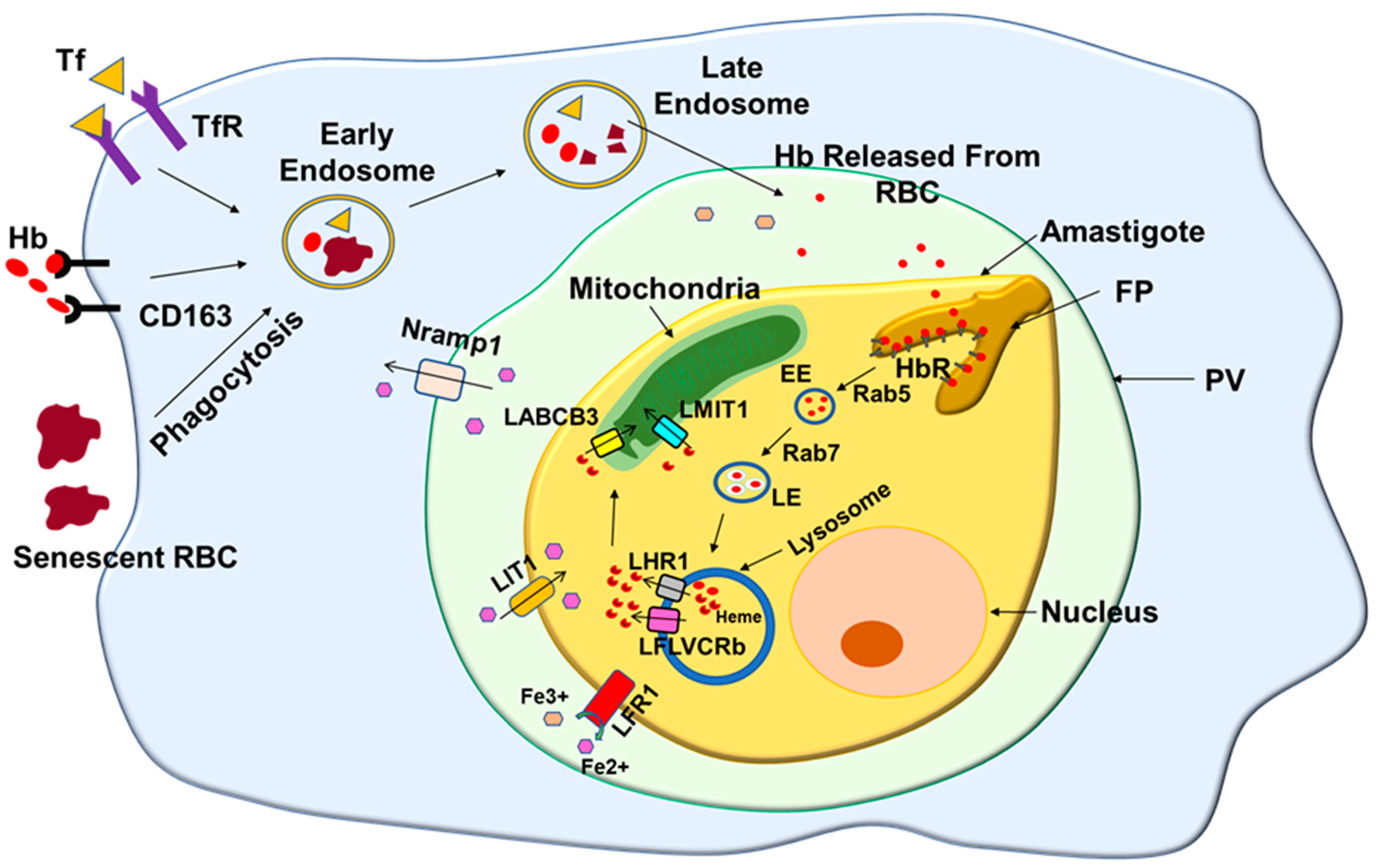
Figure 5.
Schematic diagram shows the heme acquisition process in Leishmania. Macrophages engulf senescent RBC and internalize hemoproteins such as transferrin (Tf) and hemoglobin (Hb) by specific receptor-mediated endocytic processes. These hemoproteins and RBCs are transported to the low pH compartment, where iron and hemoglobin are released from their respective hemoprotein. Leishmania residing in such compartment takes in the hemoglobin via their hemoglobin receptor (HbR). Subsequently, internalized Hb is transported to the parasite lysosomes, where it is degraded and releases heme. The released heme is transported to the parasite cytosol by heme transporter LHR1 and FLVCRb, which is finally transferred into mitochondria by mitochondrial heme importer LABCB3 and LMIT1. In addition, Fe3+ iron released by the degradation of transferrin in low acidity parasitophorous vacuole (PV) first reduces to Fe2+ by LFR1 and is imported by the parasite via its heme transporter LIT1.
Figure 5.
Schematic diagram shows the heme acquisition process in Leishmania. Macrophages engulf senescent RBC and internalize hemoproteins such as transferrin (Tf) and hemoglobin (Hb) by specific receptor-mediated endocytic processes. These hemoproteins and RBCs are transported to the low pH compartment, where iron and hemoglobin are released from their respective hemoprotein. Leishmania residing in such compartment takes in the hemoglobin via their hemoglobin receptor (HbR). Subsequently, internalized Hb is transported to the parasite lysosomes, where it is degraded and releases heme. The released heme is transported to the parasite cytosol by heme transporter LHR1 and FLVCRb, which is finally transferred into mitochondria by mitochondrial heme importer LABCB3 and LMIT1. In addition, Fe3+ iron released by the degradation of transferrin in low acidity parasitophorous vacuole (PV) first reduces to Fe2+ by LFR1 and is imported by the parasite via its heme transporter LIT1.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Putative Rab homologues in Leishmania.
| Accession Number | Putative Homologue | Location |
|---|---|---|
| KP308372 | Rab5a | Early endosome (Ref. [51]) |
| AY357217 | Rab5b | Early endosome (Ref. [52]) |
| EF507729 | Rab7 | Late endosome (Ref. [54]) |
| KT003639 | Rab1a | Golgi (Ref. [55]) |
| CBZ12137 | Rab1b | ? |
| KY484911 | Sar1 | ER (Ref. [56]) |
| CAJ08532 | Rab4 | ? |
| CAJ02862 | Rab11a | ? |
| CAJ08707 | Rab11b | ? |
| CAJ08729 | Rab2a | ? |
| AYU81761.1 | Rab2b | ? |
| CBZ11916 | Rab6 | ? |
| CAJ07017 | Rab14 | ? |
| CAJ06306 | Rab28 | ? |
| CAJ06472 | Rab18 | ? |
| XP003874984 | Rab23 | ? |
| CCM15425 | Rab21 | ? |
| XP001681766 | Rab20 | ? |
| XP003859234 | Rab31 | ? |
| TPP46114 | Rab35 | ? |
| CAJ08513 | ARF1 | ? |
| KAG5499224.1 | Rab10 | ? |
| TPP47221.1 | Rab39a | ? |
| KAG5475306.1 | Small GTPase | ? |
| KAG5503666.1 | Small GTPase | ? |
| XP001465392.1 | Small GTPase | ? |
| XP003860604.1 | Small GTPase | ? |
Note: To identify the Rab homolog from Leishmania, we performed a BLAST search by using respective mouse Rab sequence as a query, which identified putative Rabs in the parasite as mentioned in Table 1. ? denotes not characterized.
Table 2.
Putative SNARE homologs in Leishmania.
| Accession Number | Putative Homologue | Location |
|---|---|---|
| CAJ07097 | Syntaxin-7 | ? |
| CBZ12338 | Qa-SNARE | ? |
| CAJ08445 | Syntaxin 5 | ? |
| CAJ05451 | Syntaxin-1b isoform X2 | ? |
| CAJ05450 | Syntaxin-1b isoform x3 | ? |
| CAJ06328 | Syntaxin-16 isoform g | ? |
| CBZ12899 | Syntaxin-16 isoform d | ? |
| CAJ07029 | Qa-SNARE | ? |
| CAJ07013 | Qa-SNARE | ? |
| CAJ03503 | Qa-SNARE | ? |
| CAJ02128 | Qb-SNARE | ? |
| CAJ07086 | Qb-SNARE | ? |
| CAJ03721 | Qb-SNARE | ? |
| CAJ09368 | Qb-SNARE | ? |
| CAJ05002 | Qb-SNARE | ? |
| CAJ05003 | Qb-SNARE | ? |
| CAJ04850 | Syntaxin 6 | ? |
| CBZ12394 | BET1 | ? |
| CAJ04404 | Qc-SNARE | ? |
| CAJ04086 | Qc-SNARE | ? |
| CAJ07125 | SEC22 | ? |
| CAJ02181 | Qc-SNARE protein | ? |
| CBZ12839 | Synaptobrevin | ? |
| CAJ08743 | R-SNARE | ? |
| CAJ04725 | VAMP 4 | ? |
| CBZ12308 | VAMP 7 | ? |
| CAJ07162 | SNARE protein | ? |
Note: To identify the SNARE homolog from Leishmania, we performed a BLAST search by using respective mouse SNARE sequence as a query, which identified putative SNARE molecules in the parasite as mentioned in Table 2. ? denotes not characterized.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ansari, I.; Basak, R.; Mukhopadhyay, A. Hemoglobin Endocytosis and Intracellular Trafficking: A Novel Way of Heme Acquisition by Leishmania. Pathogens 2022, 11, 585. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050585
AMA Style
Ansari I, Basak R, Mukhopadhyay A. Hemoglobin Endocytosis and Intracellular Trafficking: A Novel Way of Heme Acquisition by Leishmania. Pathogens. 2022; 11(5):585. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050585
Chicago/Turabian StyleAnsari, Irshad, Rituparna Basak, and Amitabha Mukhopadhyay. 2022. "Hemoglobin Endocytosis and Intracellular Trafficking: A Novel Way of Heme Acquisition by Leishmania" Pathogens 11, no. 5: 585. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050585
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.