
An Alternative Culture Medium for Continuous In Vitro Propagation of the Human Pathogen Babesia duncani in Human Erythrocytes


Abstract
:1. Introduction
2. Results
2.1. DMEM-F12 Medium Supports the In Vitro growth of B. duncani WA-1 in Human RBCs
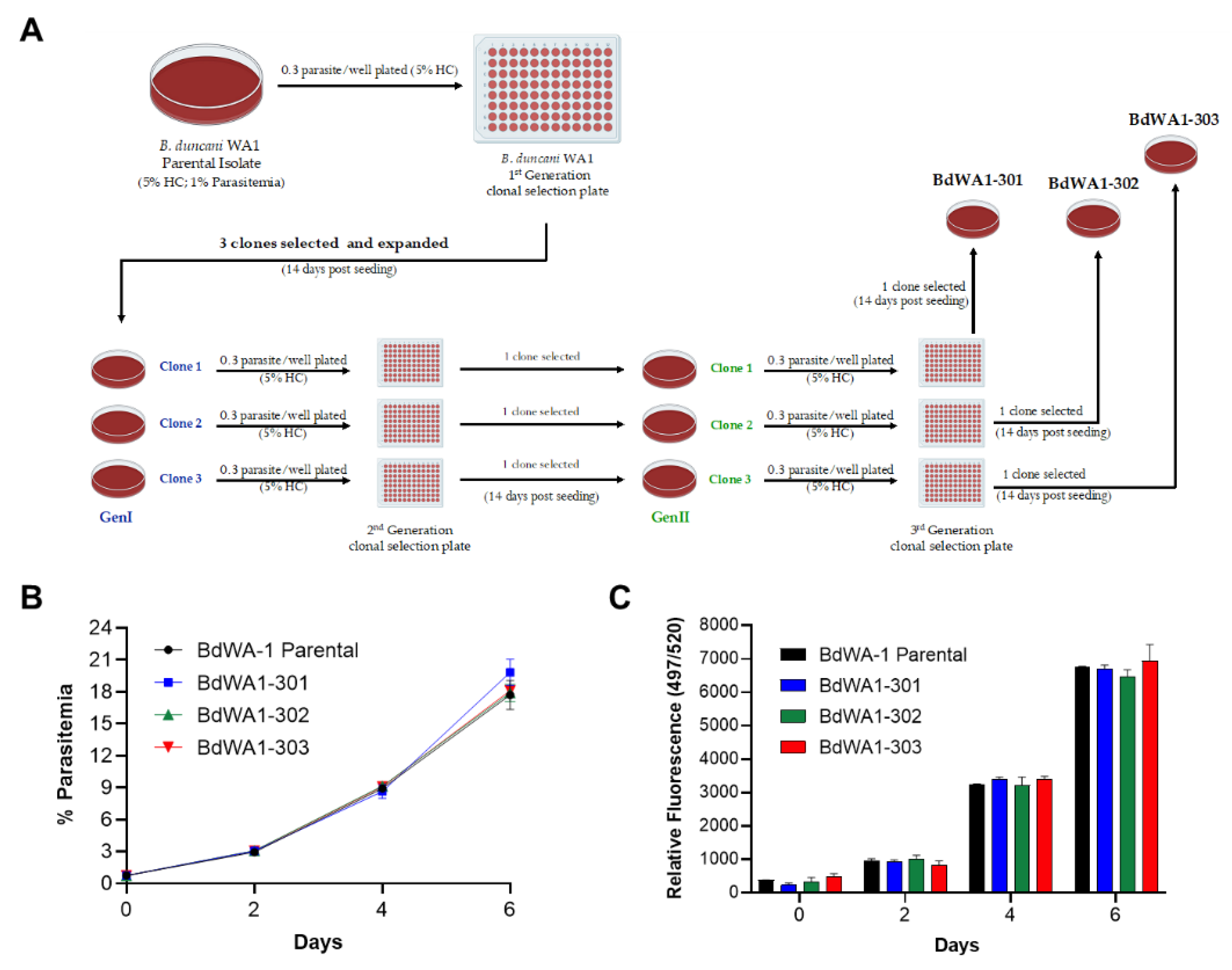
2.2. Derivation of New Lines of B. duncani WA-1 Clinical Isolate
2.3. New B. duncani Clones Are Virulent in Mice
3. Discussion
4. Materials and Methods
4.1. In Vitro Parasite Culture of B. duncani WA-1 in Different Growth Media
4.2. In Vitro Culture of B. duncani in Human RBCs
4.3. Cloning of B. duncani WA-1 Clinical Isolate
4.4. Comparison of In Vitro growth of B. duncani WA-1 and Clonal Lines BdWA1-301, BdWA1-302 and BdWA1-303
4.5. Immunofluorescence Assay
4.6. Ethics Statement
4.7. Virulence Assays in Mice
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paules, C.I.; Marston, H.D.; Bloom, M.E.; Fauci, A.S. Tickborne diseases-confronting a growing threat. N. Engl. J. Med. 2018, 379, 701–703. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.R.; Ben Mamoun, C. Babesiosis. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2018. [Google Scholar]
- Renard, I.; Ben Mamoun, C. Treatment of human babesiosis: Then and now. Pathogens 2021, 10, 1120. [Google Scholar] [CrossRef] [PubMed]
- Kjemtrup, A.M.; Conrad, P.A. Human babesiosis: An emerging tick-borne disease. Int. J. Parasitol. 2000, 30, 1323–1337. [Google Scholar] [CrossRef]
- Vannier, E.; Krause, P.J. Human babesiosis. N. Engl. J. Med. 2012, 366, 2397–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herwaldt, B.L.; Kjemtrup, A.M.; Conrad, P.A.; Barnes, R.C.; Wilson, M.; McCarthy, M.G.; Sayers, M.H.; Eberhard, M.L. Transfusion-transmitted babesiosis in Washington state: First reported case caused by a WA1-type parasite. J. Infect. Dis. 1997, 175, 1259–1262. [Google Scholar] [CrossRef]
- Kjemtrup, A.M.; Lee, B.; Fritz, C.L.; Evans, C.; Chervenak, M.; Conrad, P.A. Investigation of transfusion transmission of a WA1-type babesial parasite to a premature infant in California. Transfusion 2002, 42, 1482–1487. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Zintl, A.; Montero, E.; Hunfeld, K.P.; Gray, J. Human babesiosis in Europe. Pathogens 2021, 10, 1165. [Google Scholar] [CrossRef]
- Herwaldt, B.; Persing, D.H.; Precigout, E.A.; Goff, W.L.; Mathiesen, D.A.; Taylor, P.W.; Eberhard, M.L.; Gorenflot, A.F. A fatal case of babesiosis in Missouri: Identification of another piroplasm that infects humans. Ann. Intern. Med. 1996, 124, 643–650. [Google Scholar] [CrossRef]
- Jia, N.; Zheng, Y.C.; Jiang, J.F.; Jiang, R.R.; Jiang, B.G.; Wei, R.; Liu, H.B.; Huo, Q.B.; Sun, Y.; Chu, Y.L.; et al. Human babesiosis caused by a Babesia crassa-like pathogen: A case series. Clin. Infect. Dis. 2018, 67, 1110–1119. [Google Scholar] [CrossRef]
- Jiang, J.F.; Zheng, Y.C.; Jiang, R.R.; Li, H.; Huo, Q.B.; Jiang, B.G.; Sun, Y.; Jia, N.; Wang, Y.W.; Ma, L.; et al. Epidemiological, clinical, and laboratory characteristics of 48 cases of Babesia venatorum infection in China: A descriptive study. Lancet Infect. Dis. 2015, 15, 196–203. [Google Scholar] [CrossRef]
- Scott, J.D.; Sajid, M.S.; Pascoe, E.L.; Foley, J.E. Detection of Babesia odocoilei in humans with babesiosis symptoms. Diagnostics 2021, 11, 947. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.; Coray, E.; Ginsberg-Peltz, J.; Smith, L. A five-week-old twin with profound anemia: A case report of asymmetric congenital babesiosis. Cureus 2022, 14, e22774. [Google Scholar] [CrossRef] [PubMed]
- Cornillot, E.; Hadj-Kaddour, K.; Dassouli, A.; Noel, B.; Ranwez, V.; Vacherie, B.; Augagneur, Y.; Bres, V.; Duclos, A.; Randazzo, S.; et al. Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res. 2012, 40, 9102–9114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.C.; Cornillot, E.; McCracken, C.; Usmani-Brown, S.; Dwivedi, A.; Ifeonu, O.O.; Crabtree, J.; Gotia, H.T.; Virji, A.Z.; Reynes, C.; et al. Genome-wide diversity and gene expression profiling of Babesia microti isolates identify polymorphic genes that mediate host-pathogen interactions. Sci. Rep. 2016, 6, 35284. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Brasov, I.; Thekkiniath, J.; Kilian, N.; Lawres, L.; Gao, R.; DeBus, K.; He, L.; Yu, X.; Zhu, G.; et al. Establishment of a continuous in vitro culture of Babesia duncani in human erythrocytes reveals unusually high tolerance to recommended therapies. J. Biol. Chem. 2018, 293, 19974–19981. [Google Scholar] [CrossRef] [Green Version]
- Chiu, J.E.; Renard, I.; Pal, A.C.; Singh, P.; Vydyam, P.; Thekkiniath, J.; Kumar, M.; Gihaz, S.; Pou, S.; Winter, R.W.; et al. Effective therapy targeting cytochrome bc1 prevents Babesia erythrocytic development and protects from lethal infection. Antimicrob. Agents Chemother. 2021, 65, e0066221. [Google Scholar] [CrossRef]
- Pal, A.C.; Renard, I.; Singh, P.; Vydyam, P.; Chiu, J.E.; Pou, S.; Winter, R.W.; Dodean, R.; Frueh, L.; Nilsen, A.C.; et al. Babesiaduncani as a model organism to study the development, virulence and drug susceptibility of intraerythrocytic parasites in vitro and in vivo. J. Infect. Dis. 2022. [Google Scholar] [CrossRef]
- Camargo, E.P. Growth and differentiation in Trypanosoma cruzi. I. Origin of metacyclic trypanosomes in liquid media. Rev. Inst. Med. Trop. Sao Paulo 1964, 6, 93–100. [Google Scholar]
- Limoncu, M.E.; Balcioglu, I.C.; Yereli, K.; Ozbel, Y.; Ozbilgin, A. A new experimental in vitro culture medium for cultivation of Leishmania species. J. Clin. Microbiol. 1997, 35, 2430–2431. [Google Scholar] [CrossRef] [Green Version]
- White, S.M.; Constantin, P.E.; Claycomb, W.C. Cardiac physiology at the cellular level: Use of cultured hl-1 cardiomyocytes for studies of cardiac muscle cell structure and function. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H823–H829. [Google Scholar] [CrossRef] [Green Version]
- Quick, R.E.; Herwaldt, B.L.; Thomford, J.W.; Garnett, M.E.; Eberhard, M.L.; Wilson, M.; Spach, D.H.; Dickerson, J.W.; Telford, S.R., 3rd; Steingart, K.R.; et al. Babesiosis in Washington state: A new species of babesia? Ann. Intern. Med. 1993, 119, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Dao, A.H.; Eberhard, M.L. Pathology of acute fatal babesiosis in hamsters experimentally infected with the WA-1 strain of Babesia. Lab. Investig. 1996, 74, 853–859. [Google Scholar] [PubMed]
- McCormack, K.A.; Alhaboubi, A.; Pollard, D.A.; Fuller, L.; Holman, P.J. In vitro cultivation of Babesia duncani (Apicomplexa: Babesiidae), a zoonotic hemoprotozoan, using infected blood from Syrian hamsters (Mesocricetus auratus). Parasitol. Res. 2019, 118, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Claycomb, W.C.; Lanson, N.A., Jr.; Stallworth, B.S.; Egeland, D.B.; Delcarpio, J.B.; Bahinski, A.; Izzo, N.J., Jr. Hl-1 cells: A cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proc. Natl. Acad. Sci. USA 1998, 95, 2979–2984. [Google Scholar] [CrossRef] [Green Version]
- Downie, M.J.; Kirk, K.; Mamoun, C.B. Purine salvage pathways in the intraerythrocytic malaria parasite Plasmodium falciparum. Eukaryot. Cell 2008, 7, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Zuzarte-Luis, V.; Mota, M.M. Parasite sensing of host nutrients and environmental cues. Cell Host Microbe 2018, 23, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Augagneur, Y.; Jaubert, L.; Schiavoni, M.; Pachikara, N.; Garg, A.; Usmani-Brown, S.; Wesolowski, D.; Zeller, S.; Ghosal, A.; Cornillot, E.; et al. Identification and functional analysis of the primary pantothenate transporter, PfPAT, of the human malaria parasite Plasmodium falciparum. J. Biol. Chem. 2013, 288, 20558–20567. [Google Scholar] [CrossRef] [Green Version]
- Downie, M.J.; El Bissati, K.; Bobenchik, A.M.; Nic Lochlainn, L.; Amerik, A.; Zufferey, R.; Kirk, K.; Ben Mamoun, C. Pfnt2, a permease of the equilibrative nucleoside transporter family in the endoplasmic reticulum of Plasmodium falciparum. J. Biol. Chem. 2010, 285, 20827–20833. [Google Scholar] [CrossRef] [Green Version]
- El Bissati, K.; Downie, M.J.; Kim, S.K.; Horowitz, M.; Carter, N.; Ullman, B.; Ben Mamoun, C. Genetic evidence for the essential role of PfNT1 in the transport and utilization of xanthine, guanine, guanosine and adenine by Plasmodium falciparum. Mol. Biochem. Parasitol. 2008, 161, 130–139. [Google Scholar] [CrossRef] [Green Version]
- El Bissati, K.; Zufferey, R.; Witola, W.H.; Carter, N.S.; Ullman, B.; Ben Mamoun, C. The plasma membrane permease PfNT1 is essential for purine salvage in the human malaria parasite Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2006, 103, 9286–9291. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Complete Claycomb Medium | Complete DMEM-F12 Medium | Complete HL-1 Medium | |
|---|---|---|---|
| Base Medium | Claycomb Medium (Sigma: Cat. No.: 51800C) | DMEM-F12 Medium (Lonza: Cal. No.: BE04-687F/U1) | HL-1 Medium (LonzaTM: Discontinued) |
| Supplements | 20% Fetal Bovine Serum (Heat Inactivated) (Gibco, Cat. No.: 10438-026) | ||
| 1× of HT media supplement (50×) (Sigma, Cat. No.: H0137-10VL) | |||
| 1× of L-Glutamine (200 mM; 100×) (Gibco, Cat. No.: 25030-081) | |||
| 1× of Antimycotic-Antibiotic (100×) (Gibco, Cat. No.: 15240-062) | |||
| 1× of Gentamicin (10 mg/mL; 100×) (Gibco, Cat. No.: 15710-072) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.; Pal, A.C.; Mamoun, C.B. An Alternative Culture Medium for Continuous In Vitro Propagation of the Human Pathogen Babesia duncani in Human Erythrocytes. Pathogens 2022, 11, 599. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050599
Singh P, Pal AC, Mamoun CB. An Alternative Culture Medium for Continuous In Vitro Propagation of the Human Pathogen Babesia duncani in Human Erythrocytes. Pathogens. 2022; 11(5):599. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050599
Chicago/Turabian StyleSingh, Pallavi, Anasuya C. Pal, and Choukri Ben Mamoun. 2022. "An Alternative Culture Medium for Continuous In Vitro Propagation of the Human Pathogen Babesia duncani in Human Erythrocytes" Pathogens 11, no. 5: 599. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050599