
The Influence of Contracaecum larvae (Nematoda, Anisakidae) Parasitism on the Population of Prussian carp (Carassius gibelio) in Lake Sakadaš, Croatia

 ,
,
Abstract
:1. Introduction
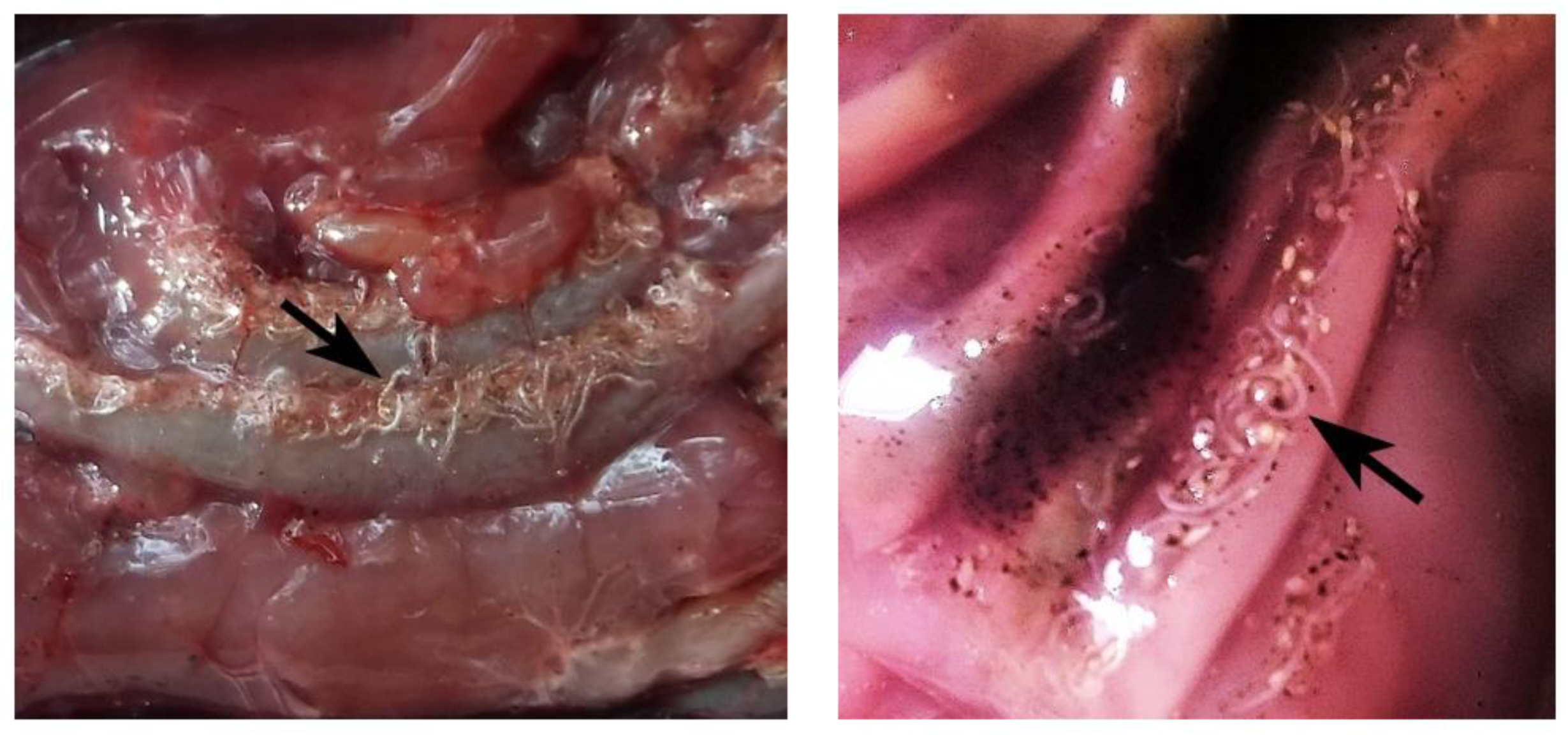
2. Results
3. Discussion
4. Materials and Methods
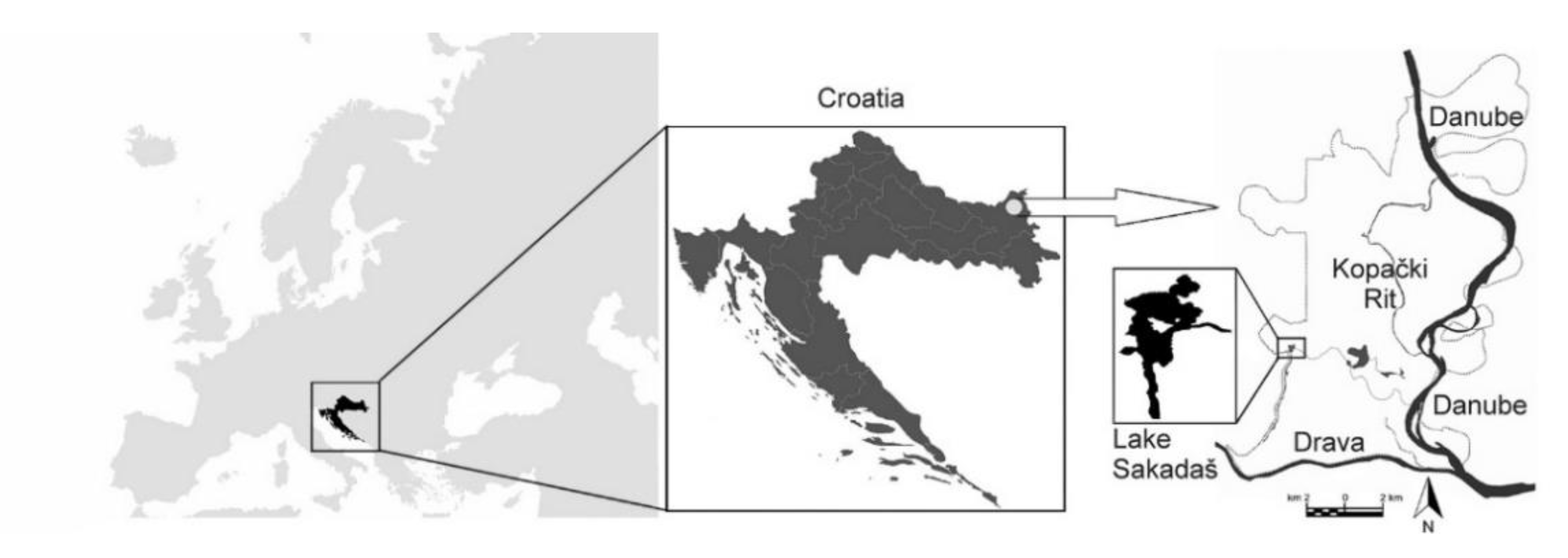
4.1. Study Area
4.2. Prussian carp (Carassius gibelio) Sampling and Parasite Inspection
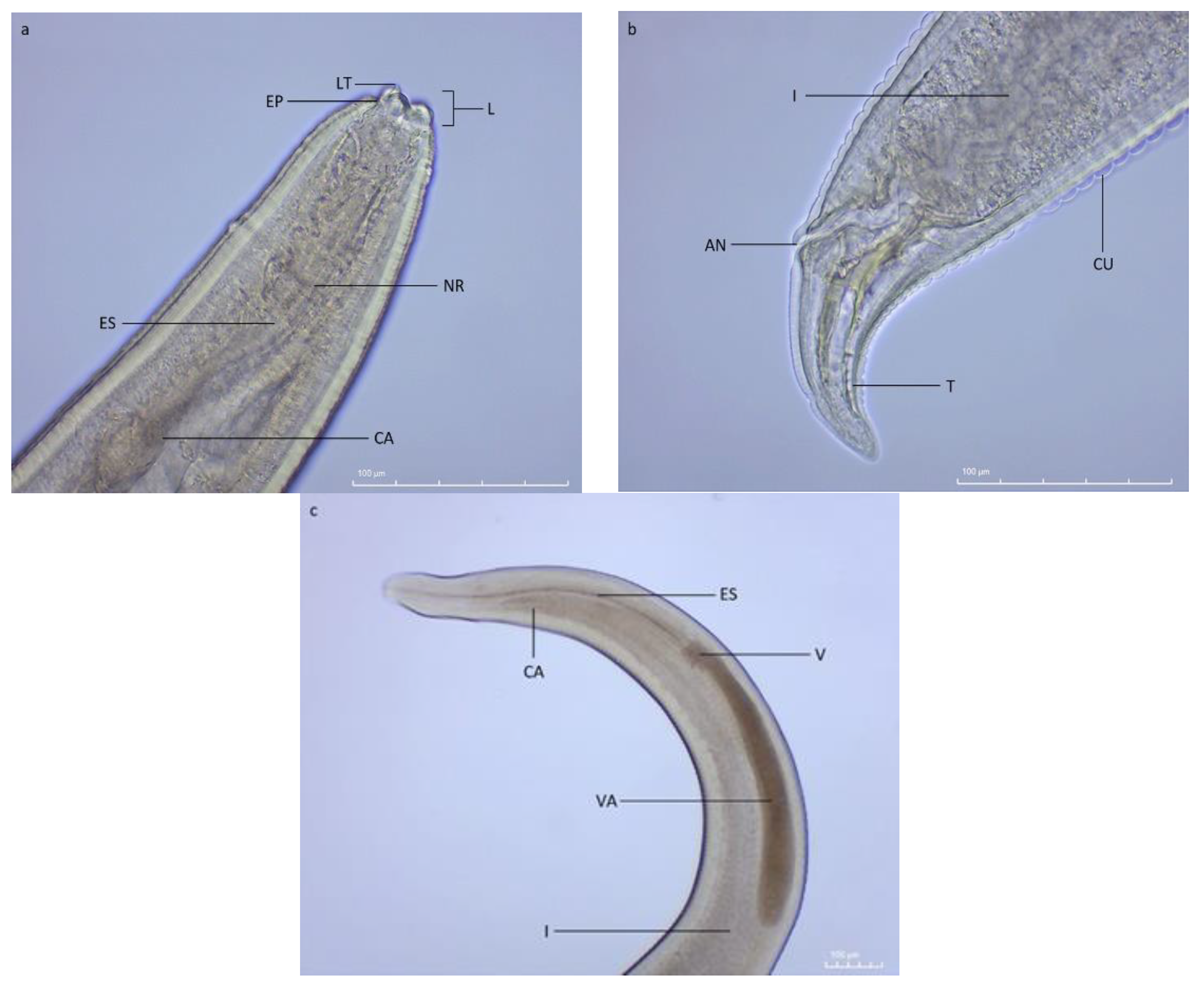
4.3. Parasite Identification and Measurements
4.4. Parasite-Host Relationship
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to Linnaeus: How Many Parasites? How Many Hosts? PNAS 2008, 105 (Suppl. 1), 11482–11489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sures, B.; Nachev, M.; Pahl, M.; Grabner, D.; Selbach, C. Parasites as Drivers of Key Processes in Aquatic Ecosystems: Facts and Future Directions. Exp. Parasitol. 2017, 180, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Timi, J.T.; MacKenzie, K. Parasites in Fisheries and Mariculture. Parasitology 2015, 142, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, K.; Buchmann, K.; Székely, C. Phylum Nematoda. In Fish Diseases and Disorders. Volume 1: Protozoan and Metazoan Infections; Woo, P.T.K., Ed.; CABI: Wallingford, UK, 2006; Volume 1, pp. 417–443. [Google Scholar] [CrossRef]
- Olivero-Verbel, J.; Baldiris-Avila, R.; Guette-Fernandez, J.; Benavides-Alvarez, A.; Mercado-Camargo, J.; Arroyo-Salgado, B. Contracaecum sp. infection in Hoplias Malabaricus (Moncholo) from Rivers and Marshes of Colombia. Vet. Parasitol. 2006, 140, 90–97. [Google Scholar] [CrossRef]
- Maceda-Veiga, A.; Green, A.J.; Poulin, R.; Lagrue, C. Body Condition Peaks at Intermediate Parasite Loads in the Common Bully Gobiomorphus Cotidianus. PLoS ONE 2016, 11, e0168992. [Google Scholar] [CrossRef] [Green Version]
- Farooq, T.; Khan, I.; Tak, I. urR.; Dar, S.A.; Yousuf, A.R. Endoparasites of Some Economically Important Food Fishes of River Jhelum, Kashmir (India). J. Parasit. Dis. 2016, 40, 860–871. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.C. Nematode Parasites of Vertebrates: Their Development and Transmission; CABI: Wallingford, UK, 2000. [Google Scholar] [CrossRef]
- Fagerholm, H.P. Systematic Implications of Male Caudal Morphology in Ascaridoid Nematode Parasites. Syst. Parasitol. 1991, 19, 215–229. [Google Scholar] [CrossRef]
- Nadler, S.A.; D’Amelio, S.; Dailey, M.D.; Paggi, L.; Siu, S.; Sakanari, J.A. Molecular Phylogenetics and Diagnosis of Anisakis, Pseudoterranova, and Contracaecum from Northern Pacific Marine Mammals. J. Parasitol. 2005, 91, 1413–1429. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, S. Parasite Loss or Parasite Gain? Story of Contracaecum Nematodes in Antipodean Waters. Parasite Epidemiol. Control 2019, 4, e00087. [Google Scholar] [CrossRef]
- Nemys (Ed.) Nemys: World Database of Nematodes. Contracaecum Railliet & Henry, 1912. Accessed through: World Register of Marine Species. 2022. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=22849 (accessed on 7 April 2022).
- Al-Zubaidy, A.B. Prevalence and Densities of Contracaecum sp. Larvae in Liza Abu (Heckel, 1843) from Different Iraqi Water Bodies. J. King Abdulaziz Univ. Mar. Sci. 2009, 20, 3–17. [Google Scholar] [CrossRef]
- Markogliese, D.J. The role of zooplankton in the transmission of helminth parasites to fish. Rev. Fish Biol. Fish. 1995, 5, 336–371. [Google Scholar] [CrossRef]
- Martins, M.L.; Onaka, E.M.; Fenerick, J., Jr. Larval Contracaecum sp. (Nematoda: Anisakidae) in Hoplias malabaricus and Hoplerythrinus unitaeniatus (Osteichthyes: Erythrinidae) of economic importance in occidental marshlands of Maranhão, Brazil. Vet. Parasitol. 2005, 127, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Moravec, F. Experimental studies on the development of Contracaecum rudolphii (Nematoda: Anisakidae) in copepod and fish paratenic hosts. Folia Parasitol. 2009, 56, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barson, M. The Occurrence of Contracaecum sp. Larvae (Nematoda: Anisakidae) in the Catfish Clarias gariepinus (Burchell) from Lake Chivero, Zimbabwe. Onderstepoort J. Vet. Res. 2004, 71, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Molnár, K.; Székely, C.; Baska, F.; Müller, T.; Zuo, S.; Kania, P.W.; Nowak, B.; Buchmann, K. Differential Survival of 3rd Stage Larvae of Contracaecum rudolphii Type B Infecting Common Bream (Abramis brama) and Common Carp (Cyprinus carpio). Parasitol. Res. 2019, 118, 2811–2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcogliese, D.J.; Cone, D.K. Food Webs: A Plea for Parasites. Trends Ecol. Evol. 1997, 12, 320–325. [Google Scholar] [CrossRef]
- Yanong, R.P.E. Nematode (Roundworm) Infections in Fish. Univ. Florida 2002, 1–9. [Google Scholar] [CrossRef]
- Parker, G.A.; Ball, M.A.; Chubb, J.C. Evolution of Complex Life Cycles in Trophically Transmitted Helminths. I. Host Incorporation and Trophic Ascent. J. Evol. Biol. 2015, 28, 267–291. [Google Scholar] [CrossRef] [Green Version]
- Amato, J.F.R.; Monteiro, C.M.; Amato, S.B. Contracaecum rudolphii Hartwich (Nematoda, Anisakidae) from the Neotropical Cormorant, Phalacrocorax brasilianus (Aves, Phalacrocoracidae) in Southern Brazil. Rev. Bras. Zool. 2006, 23, 1284–1289. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, S.; Butcher, A.R. First Report of Human Anisakidosis in Australia. Med. J. Aust. 2011, 194, 199–200. [Google Scholar] [CrossRef]
- Schaum, E.; Müller, W. Heterocheilidiasis (Case Report). Dtsch. Med. Wochenschr. 1967, 92, 2230–2234. [Google Scholar] [CrossRef] [PubMed]
- Adroher-Auroux, F.J.; Benítez-Rodríguez, R. Anisakiasis and Anisakis: An Underdiagnosed Emerging Disease and Its Main Etiological Agents. Res. Vet. Sci. 2020, 132, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Kassai, T.; Cordero Del Campillo, M.; Euzeby, J.; Gaafar, S.; Hiepe, T.; Himonas, C.A. Standardized Nomenclature of Animal Parasitic Diseases (SNOAPAD). Vet. Parasitol. 1988, 29, 299–326. [Google Scholar] [CrossRef]
- Shibata, K.; Yoshida, Y.; Miyaoka, Y.; Emoto, S.; Kawai, T.; Kobayashi, S.; Ogasawara, K.; Taketomi, A. Intestinal Anisakiasis with Severe Intestinal Ischemia Caused by Extraluminal Live Larvae: A Case Report. Surg. Case Rep. 2020, 6, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Hirosawa, T.; Sakamoto, T.; Shimizu, T. Gastric Anisakiasis. Am. J. Med. Sci. 2020, 360, 318–319. [Google Scholar] [CrossRef] [PubMed]
- Decruyenaere, P.; Van de Maele, B.; Hulstaert, E.; Van Vlierberghe, H.; Decruyenaere, J.; Lapeere, H. IgE-Mediated Gastroallergic Anisakiasis with Eosinophilic Oesophagitis: A Case Report. Acta Clin. Belgica Int. J. Clin. Lab. Med. 2022, 77, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular Epidemiology of Anisakis and Anisakiasis: An Ecological and Evolutionary Road Map, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; Volume 99. [Google Scholar] [CrossRef]
- Mladineo, I. Anisakiasis in Europe: Emerging, Neglected, Misdiagnosed, or All of the Above? Vet. Stanica 2019, 50, 397–405. [Google Scholar]
- Fiorenza, E.A.; Wendt, C.A.; Dobkowski, K.A.; King, T.L.; Pappaionou, M.; Rabinowitz, P.; Samhouri, J.F.; Wood, C.L. It’s a Wormy World: Meta-Analysis Reveals Several Decades of Change in the Global Abundance of the Parasitic Nematodes Anisakis spp. and Pseudoterranova spp. in Marine Fishes and Invertebrates. Glob. Chang. Biol. 2020, 26, 2854–2866. [Google Scholar] [CrossRef]
- Mladineo, I.; Popović, M.; Drmić-Hofman, I.; Poljak, V. A Case Report of Anisakis pegreffii (Nematoda, Anisakidae) Identified from Archival Paraffin Sections of a Croatian Patient. BMC Infect. Dis. 2016, 16, 42. [Google Scholar] [CrossRef] [Green Version]
- Tomljanović, T.; Fabijanić, N.; Treer, T.; Matulić, D.; Piria, M.; Šprem, N.; Aničić, I.; Safner, R. Utjecaj Invazivne Vrste Riba Babuške (Carassius gibelio) Na Sastav Ihtiocenoze u Republici Hrvatskoj. In 47th Croatian and 7th International Symposium on Agriculture; Pospišil, M., Ed.; University of Zagreb, Faculty of Agriculture: Opatija, Croatia, 2012; pp. 634–636. [Google Scholar]
- Japoshvili, B.; Mumladze, L.; Murvanidze, L. The Population of Carassius gibelio (Bloch, 1782) and Its Parasites in Madatapa Lake (South Georgia). Iran. J. Fish. Sci. 2017, 16, 793–799. [Google Scholar]
- Ruppert, J.L.W.; Docherty, C.; Neufeld, K.; Hamilton, K.; MacPherson, L.; Poesch, M.S. Native Freshwater Species Get out of the Way: Prussian Carp (Carassius gibelio) Impacts Both Fish and Benthic Invertebrate Communities in North America. R. Soc. Open Sci. 2017, 4, 170400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrakovčić, M.; Brigić, A.; Buj, I.; Ćaleta, M.; Mustafić, P.; Zanella, D. Crvena knjiga slatkovodnih riba Hrvatske; Mrakovčić, M., Brigić, A., Eds.; Ministarstvo kulture, Državni zavod za zaštitu prirode, Republika Hrvatska: Zagreb, Croatia, 2006. [Google Scholar]
- Daghigh Roohi, J.; Sattari, M.; Nezamabadi, H.; Ghorbanpour, N. Occurrence and Intensity of Parasites in Prussian Carp, Carassius gibelio from Anzali Wetland, Southwest Caspian Sea. Iran. J. Fish. Sci. 2014, 13, 276–288. [Google Scholar]
- Demir, S.; Karakişi, H. Metazoan Parasite Fauna of the Prussian Carp, Carassius gibelio (Bloch, 1782) (Cyprinidae), from Marmara Lake, Turkey. Acta Zool. Bulg. 2016, 68, 265–268. [Google Scholar]
- Kucher, H.; Stock, T.M.; Das, M.K. Parasites of Prussian Carp (Carassius gibelio): An Invasive Species in Alberta, Canada. Can. J. Zool. 2019, 97, 849–853. [Google Scholar] [CrossRef] [Green Version]
- İnnal, D.; Stavrescu-Bedivan, M.; Özturk, M.O.; Özmen, Ö. First Record of Contracaecum rudolphii Hartwich, 1964 in Carassius gibelio (Bloch, 1782) From Turkey. Aquat. Sci. Eng. 2019, 35, 1–5. [Google Scholar] [CrossRef]
- Zanella, D.; Mustafić, P.; Marčić, Z.; Ćaleta, M.; Buj, I.; Horvatić, S.; Karlović, R.; Ivić, L.; Onorato, L. Diversity of freshwater ichthyofauna in nature park Kopački rit. In Proceedings of the Kopački Rit. Past, Present, Future 2021, Kopačevo, Bilje, Croatia, 30 September–1 October 2021; pp. 242–245. [Google Scholar]
- Mikuska, J.; Mikuska, T.; Romulić, M. Vodič kroz Biološku Raznolikost Kopačkog Rita—PTICE; Matica hrvatska Osijek: Osijek, Croatia, 2002; p. 112. [Google Scholar]
- Damjanović, I.; Rožac, V. The initial status of the mammalian fauna of the Nature Park Kopački rit. Book of abstracts of the 10th symposium with international participation. In Proceedings of the Kopački Rit: Past, Present, Future 2021, Kopačevo, Bilje, Croatia, 30 September–1 October 2021; pp. 44–47. [Google Scholar]
- Damjanović, I.; Rožac, V. Protocol for monitoring the Eurasian otter (Lutra lutra L.) in Nature Park Kopački rit. Book of abstracts of the 10th symposium with international participation. In Proceedings of the Kopački Rit: Past, Present, Future 2021, Kopačevo, Bilje, Croatia, 30 September–1 October 2021; pp. 52–55. [Google Scholar]
- Ramsar. Information Sheet on Ramsar Wetlands—Nature Park Kopacki Rit (Kopački Rit). October 2007. Available online: http://www.wetlands.org/reports/ris/3HR002RIS2007.pdf (accessed on 1 February 2022).
- Benčina, L.; Rožac, V.; Bolšec, B. Plan Upravljanja Parkom Prirode KOPAČKI Rit. Tikveš. 2010. Available online: https://pp-kopacki-rit.hr/wp-content/uploads/2019/06/plan_upravljanja.pdf (accessed on 2 February 2022).
- Sommerwerk, N.; Bloesch, J.; Baumgartner, C.; Bittl, T.; Čerba, D.; Csányi, B.; Davideanu, G.; Dokulil, M.; Frank, G.; Grecu, I.; et al. The Danube River Basin. In Rivers of Europe, 2nd ed.; Tockner, K., Zarfl, C., Robinson, C.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 81–180. [Google Scholar] [CrossRef]
- Mikuška, A.; Mikuška, T. A Historical Review of Great Cormorant Breeding Population in Croatia from 1960 to 2013. Cormor. Res. Group Bull. 2015, 8, 42–47. [Google Scholar]
- Mikuška, T.; Grlica, I.D.; Grgić, M.; Tomik, A. Fauna Ptica Hrvatskog Dijela Rezervata Biosphere “Mura-Drava-Dunav”; Hrvatsko društvo za zaštitu ptica i prirode: Osijek, Croatia, 2015; p. 68. [Google Scholar]
- Kanarek, G.; Bohdanowicz, J. Larval Contracaecum sp. (Nematoda: Anisakidae) in the Great Cormorant [Phalacrocorax carbo (L., 1758)] from North-Eastern Poland: A Morphological and Morphometric Analysis. Vet. Parasitol. 2009, 166, 90–97. [Google Scholar] [CrossRef]
- Pakosta, T.; Vetešník, L.; Šimková, A. A Long Temporal Study of Parasitism in Asexual-Sexual Populations of Carassius gibelio: Does the Parasite Infection Support Coevolutionary Red Queen Dynamics? Biomed. Res. Int. 2018, 2018, 6983740. [Google Scholar] [CrossRef] [Green Version]
- Dziekonska-Rynko, J.; Rokicki, J.; Mierzejewska, K.; Wziątek, B.; Bielecki, A. Location and Development of Larvae of Contracaecum rudolphii Hartwich, 1964 (Nematoda: Anisakidae) in Experimentally Infected Asps Leuciscus aspius (Linnaeus, 1758) (Pisces: Cyprinidae). Oceanol. Hydrobiol. Stud. 2015, 42, 296–301. [Google Scholar] [CrossRef]
- Køie, M.; Fagerholm, H.P. The Life Cycle of Contracaecum osculatum (Rudolphi, 18021 Sensu Stricto (Nematoda, Ascaridoidea, Anisakidae) in View of Experimental Infections. Parasitol. Res. 1995, 81, 481–489. [Google Scholar] [CrossRef]
- Buchmann, K. Fish Immune Responses against Endoparasitic Nematodes—Experimental Models. J. Fish Dis. 2012, 35, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Boane, C.; Cruz, C.; Saraiva, A. Metazoan Parasites of Cyprinus carpio L. (Cyprinidae) from Mozambique. Aquaculture 2008, 284, 59–61. [Google Scholar] [CrossRef]
- Younis, A.E.; Saad, A.I.; Rabei, J.M. The Occurrence of Contracaecum sp. Larvae (Nematoda: Anisakidae) in Four Teleostean Species from Lake Nasser, Egypt: Morphological and Molecular Studies. J. Basic Appl. Zool. 2017, 78, 9. [Google Scholar] [CrossRef]
- Llarena-Reino, M.; Piñeiro, C.; Antonio, J.; Outeriño, L.; Vello, C.; González, Á.F.; Pascual, S. Optimization of the Pepsin Digestion Method for Anisakids Inspection in the Fishing Industry. Vet. Parasitol. 2013, 191, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, S.; Suthar, J. A Revised Method of Examining Fish for Infection with Zoonotic Nematode Larvae. Int. J. Food Microbiol. 2016, 227, 13–16. [Google Scholar] [CrossRef]
- Gómez-Morales, M.A.; Castro, C.M.; Lalle, M.; Fernández, R.; Pezzotti, P.; Abollo, E.; Pozio, E. UV-Press Method versus Artificial Digestion Method to Detect Anisakidae L3 in Fish Fillets: Comparative Study and Suitability for the Industry. Fish. Res. 2018, 202, 22–28. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Allesina, S.; Arim, M.; Briggs, C.J.; De Leo, G.; Dobson, A.P.; Dunne, J.A.; Johnson, P.T.J.; Kuris, A.M.; Marcogliese, D.J.; et al. Parasites in Food Webs: The Ultimate Missing Links. Ecol. Lett. 2008, 11, 533–546. [Google Scholar] [CrossRef]
- Wilson, K.; Bjørnstad, O.N.; Dobson, A.P.; Merler, S.; Poglayen, G.; Randolf, S.E.; Read, A.F.; Skorping, A. Heterogeneities in Macroparasite Infections: Patterns and Processes. In The Ecology of Wildlife Diseases; Hudson, P.J., Rizzoli, A., Grenfell, H.H., Dobson, A.P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 6–44. [Google Scholar]
- Lagrue, C.; Poulin, R. The Scaling of Parasite Biomass with Host Biomass in Lake Ecosystems: Are Parasites Limited by Host Resources? Ecography 2016, 39, 507–514. [Google Scholar] [CrossRef]
- Jin, S.; Yan, X.; Zhang, H.; Fan, W. Weight–Length Relationships and Fulton’s Condition Factors of Skipjack Tuna (Katsuwonus pelamis) in the Western and Central Pacific Ocean. PeerJ 2015, 3, e758. [Google Scholar] [CrossRef] [Green Version]
- De Giosa, M.; Czerniejewski, P.; Rybczyk, A. Seasonal Changes in Condition Factor and Weight-Length Relationship of Invasive Carassius gibelio (Bloch, 1782) from Leszczynskie Lakeland, Poland. Adv. Zool. 2014, 2014, 678763. [Google Scholar] [CrossRef] [Green Version]
- Jisr, N.; Younes, G.; Sukhn, C.; El-Dakdouki, M.H. Length-Weight Relationships and Relative Condition Factor of Fish Inhabiting the Marine Area of the Eastern Mediterranean City, Tripoli-Lebanon. Egypt. J. Aquat. Res. 2018, 44, 299–305. [Google Scholar] [CrossRef]
- Pope, K.; Lochmann, S.; Young, M. Methods for Assessing Fish Populations. In Inland Fisheries Management in North America, 3rd edition; Quist, M.C., Hubert, W.A., Eds.; American Fisheries Society: Bethesda, MD, USA, 2010. [Google Scholar]
- Guidelli, G.; Tavechio, W.L.G.; Takemoto, R.M.; Pavanelli, G.C. Relative Condition Factor and Parasitism in Anostomid Fishes from the Floodplain of the Upper Paraná River, Brazil. Vet. Parasitol. 2011, 177, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulin, R. Variation in the Intraspecific Relationship between Fish Length and Intensity of Parasitic Infection: Biological and Statistical Causes. J. Fish Biol. 2000, 56, 123–137. [Google Scholar] [CrossRef]
- Iyaji, F.; Etim, L.; Eyo, J. Parasite Assemblages in Fish Hosts. Bio-Research 2010, 7, 561–570. [Google Scholar] [CrossRef]
- Treer, T.; Piria, M.; Šprem, N. The Relationship between Condition and Form Factors of Freshwater Fishes of Croatia. J. Appl. Ichthyol. 2009, 25, 608–610. [Google Scholar] [CrossRef]
- Treer, T.; Sperm, N.; Torcu Koç, H.; Sun, Y.; Piria, M. Length–Weight Relationships of Freshwater Fishes of Croatia. J. Appl. Ichthyol. 2008, 24, 626–628. [Google Scholar] [CrossRef]
- Stavrescu-Bedivan, M.; Scaeteanu, G.; Madjar, R.M.; Matei, P.; Tobă, G. Comparative Study of Length-Weight Relationship, Size Structure and Fulton’s Condition Factor for Prussian Carp from Different Romanian Aquatic Ecosystems. AgroLife Sci. J. 2015, 4, 132–139. [Google Scholar]
- Neff, B.D.; Cargnelli, L.M. Relationships between Condition Factors, Parasite Load and Paternity in Bluegill Sunfish, Lepomis macrochirus. Environ. Biol. Fishes 2004, 71, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Mashego, S.N.; Saayman, J.E. Observations on the Prevalence of Nematode Parasites of the Catfish, Clarias gariepinus (Burchell 1822), in Lebowa, South Africa. South African J. Wildl. Res. 1981, 11, 46–48. [Google Scholar]
- Rokicki, J.; Sołtysiak, Z.; Dziekońska-Rynko, J.; Borucińska, J. Pathology Associated with Contracaecum rudolphii (Nematoda: Anisakidae) Infection in the Great Cormorant Phalacrocorax carbo (L. 1758). Helminthologia 2011, 48, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Sato, T.; Shirai, W.; Kikuchi, S. Parasites and Related Pathological Lesions in the Gastrointestinal Tract of a Seal (Phoka vitulina Linnaeus). J. Vet. Med. Sci. 1998, 60, 1025–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, U. Landschaftsökologische Charakterisierung des Kopački Rit unter Besonderer Berücksichtigung von Flusslandschaftsformen sowie Deren Genese und Typologie. Ph.D. Thesis, Universität Wien, Wien, Austria, 2005. [Google Scholar]
- Aloo, P.A. A comparative study of helminth parasites from the fish Tilapia zillii and Oreochromis leucostictus in Lake Naivasha and Oloidien Bay, Kenya. Journal of Helminthology. J. Helminthol. 2002, 76, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kottelat, M.; Freyhof, J. (Eds.) Handbook of European Freshwater Fishes; Copeia: Cornol, Switzerland; Berlin, Germany, 2007; p. 646. [Google Scholar]
- Seinhorst, J.W. A Rapid Method for the Transfer of Nematodes from Fixative to Anhydrous Glycerin. Nematologica 1957, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Quang, N.X.; Chau, N.N.; Vanreusel, A. Nematode Morphometry and Biomass Patterns in Relation to Community Characteristics and Environmental Variables in the Mekong Delta. Raffles Bull. Zool. 2014, 62, 501–512. [Google Scholar]
- Anderson, R.C.; Chabaud, A.G.; Willmott, S. CIH Keys to the Nematode Parasites of Vertebrates. CAB International: Wallingford, UK, 1974. [Google Scholar]
- Lima, F.; Pozza, A.; Lehmann, P. Contracaecum spp. (Nematoda: Anisakidae) and Eustrongylides spp. (Nematoda: Dioctophymatidae) Nematode Larvae with Zoonotic Potencial Found in Two Fish Species from TramandaÍ River Basin, Southern Brazil. Bol. do Inst. Pesca 2019, 45, e495. [Google Scholar] [CrossRef]
- Cannon, L.R.G. Some Larval Ascaridoids from South-Eastern Queensland Marine Fishes. Int. J. Parasitol. 1977, 7, 233–243. [Google Scholar] [CrossRef]
- Andrassy, I. The determination of volume and weight of nematodes. Acta Zool. 1956, 2, 1–15. [Google Scholar]
- Treer, T.; Matulić, D.; Bogdanović, G.; Aničić, I.; Safner, R.; Piria, M.; Šprem, N.; Tomljanović, T. The condition of allochtonous fishes in the Mediterranean Vransko Lake. J. Appl. Ichthyol. 2010, 27, 965–967. [Google Scholar] [CrossRef]
- Le Cren, E.D. The Length-Weight Relationship and Seasonal Cycle in Gonad Weight and Condition in the Perch (Perca fluviatilis). Br. Ecol. Soc. 1951, 20, 201–219. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2020. Available online: https://www.R-project.org/ (accessed on 17 January 2022).
- Reiczigel, J.J.; Marozzi, M.; Fábián, I.; Rózsa, L. Biostatistics for Parasitologists—A Primer to Quantitative Parasitology. Trends Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et Al. Revisited. J. Parasitol. 1997, 83, 575. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Infected Fish a (N = 20) | Uninfected Fish a (N = 179) |
|---|---|---|
| TL ± SD (range) | 36.5 ± 2.0 (33.2–44.4) | 32.8 ± 7.6 (7.5–42.0) |
| SL ± SD (range) | 28.6 ± 1.8 (26.0–32.5) | 26.7 ± 6.3 (5.5–34.0) |
| Fish weight ± SD (range) | 803.0 ± 105.4 (660.0–1053.0) | 852.6 ± 324.8 (5.0–1410.0) |
| CF ± SD (range) | 1.64 ± 0.14 (1.32–1.93) | 2.08 ± 0.34 (0.98–3.02) |
| Sex | SL ± SD | TL ± SD | Fish Weight ± SD | Intensity ± SD | CF ± SD |
|---|---|---|---|---|---|
| M (N = 7) | 36.1 ± 1.4 | 28.4 ± 1.2 | 741.0 ± 51.0 | 414.1 ± 339.3 | 1.58 ± 0.15 |
| F (N = 13) | 36.7 ± 2.2 | 28.8 ± 2.1 | 836.0 ± 114.0 | 339.3 ± 319.2 | 1.68 ± 0.13 |
| TL | SL | Fish Weight | CF | |
|---|---|---|---|---|
| PresAbsN | 0.16 * | 0.09 | −0.05 | −0.37 ** |
| NoN | 0.12 | 0.07 | −0.03 | −0.27 ** |
| massN | 0.09 | 0.05 | −0.01 | −0.19 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuić, N.; Turković Čakalić, I.; Vlaičević, B.; Stojković Piperac, M.; Čerba, D. The Influence of Contracaecum larvae (Nematoda, Anisakidae) Parasitism on the Population of Prussian carp (Carassius gibelio) in Lake Sakadaš, Croatia. Pathogens 2022, 11, 600. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050600
Vuić N, Turković Čakalić I, Vlaičević B, Stojković Piperac M, Čerba D. The Influence of Contracaecum larvae (Nematoda, Anisakidae) Parasitism on the Population of Prussian carp (Carassius gibelio) in Lake Sakadaš, Croatia. Pathogens. 2022; 11(5):600. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050600
Chicago/Turabian StyleVuić, Nera, Ivana Turković Čakalić, Barbara Vlaičević, Milica Stojković Piperac, and Dubravka Čerba. 2022. "The Influence of Contracaecum larvae (Nematoda, Anisakidae) Parasitism on the Population of Prussian carp (Carassius gibelio) in Lake Sakadaš, Croatia" Pathogens 11, no. 5: 600. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050600