
Molecular Survey of Vector-Borne Pathogens in Ticks, Sheep Keds, and Domestic Animals from Ngawa, Southwest China

 and
and
Abstract
:1. Introduction
2. Results
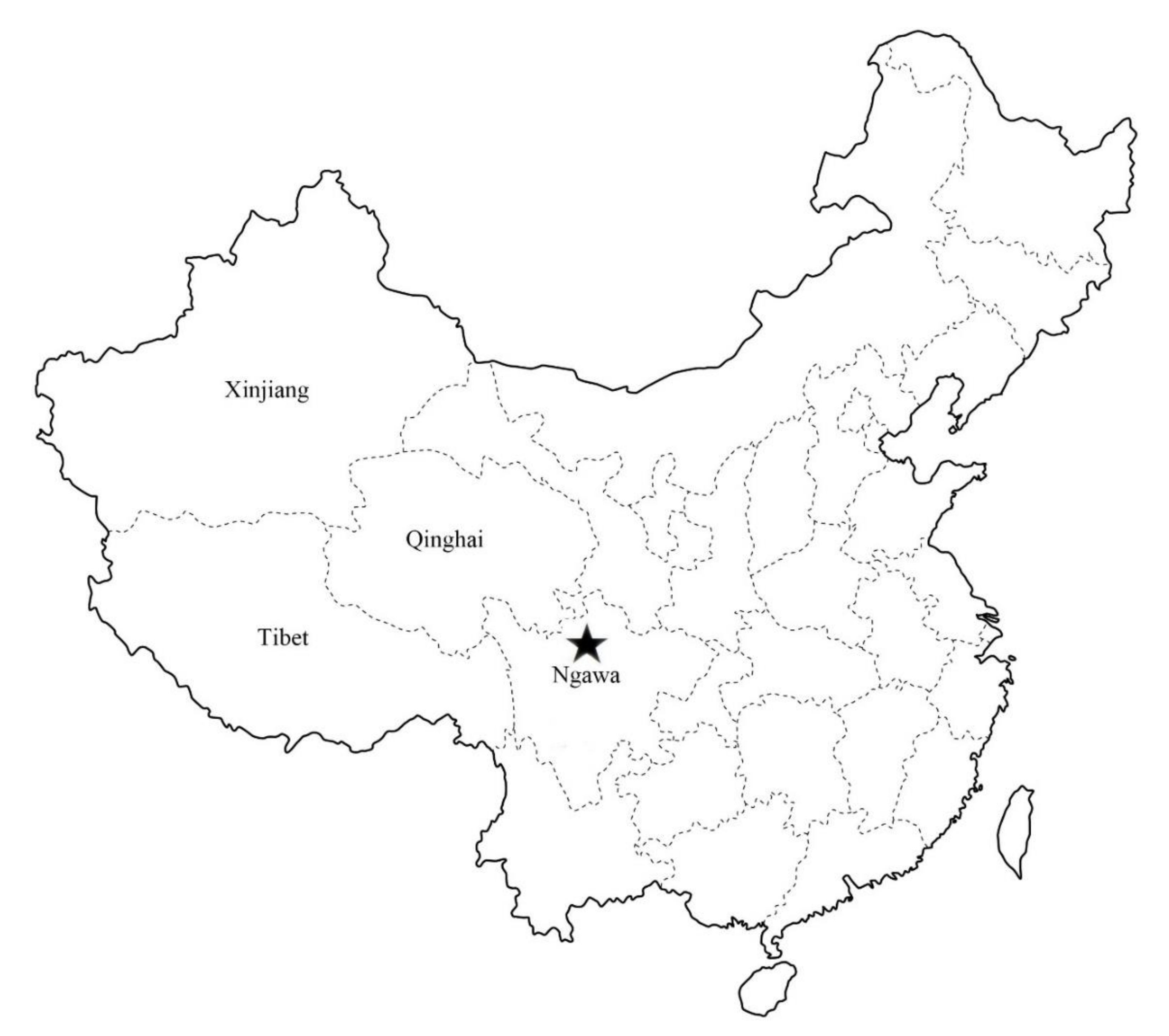
2.1. Sample Collection
2.2. PCR Detection
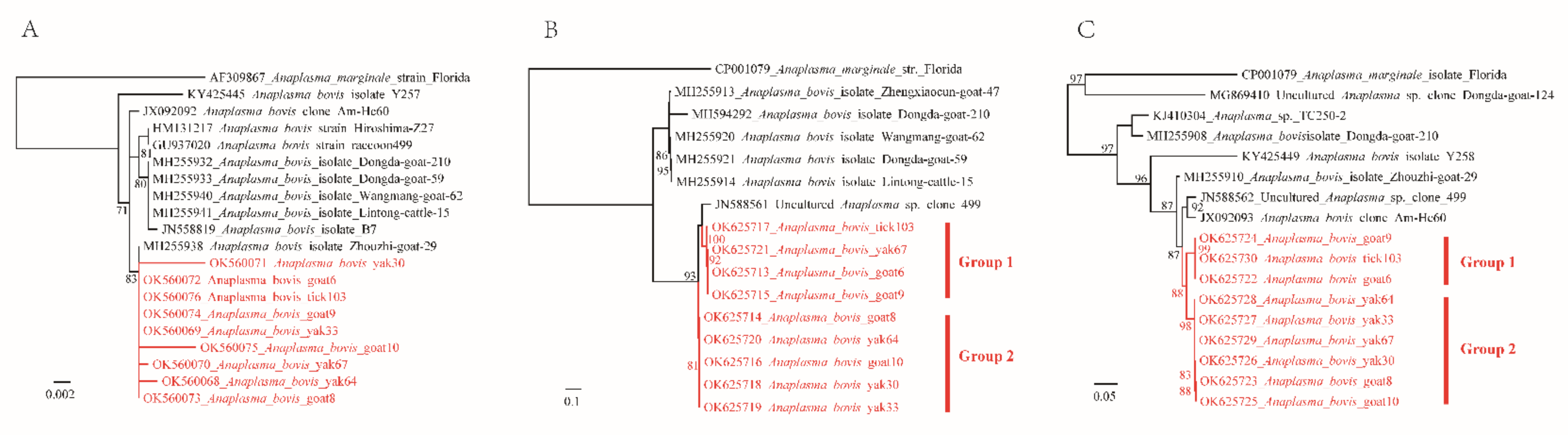
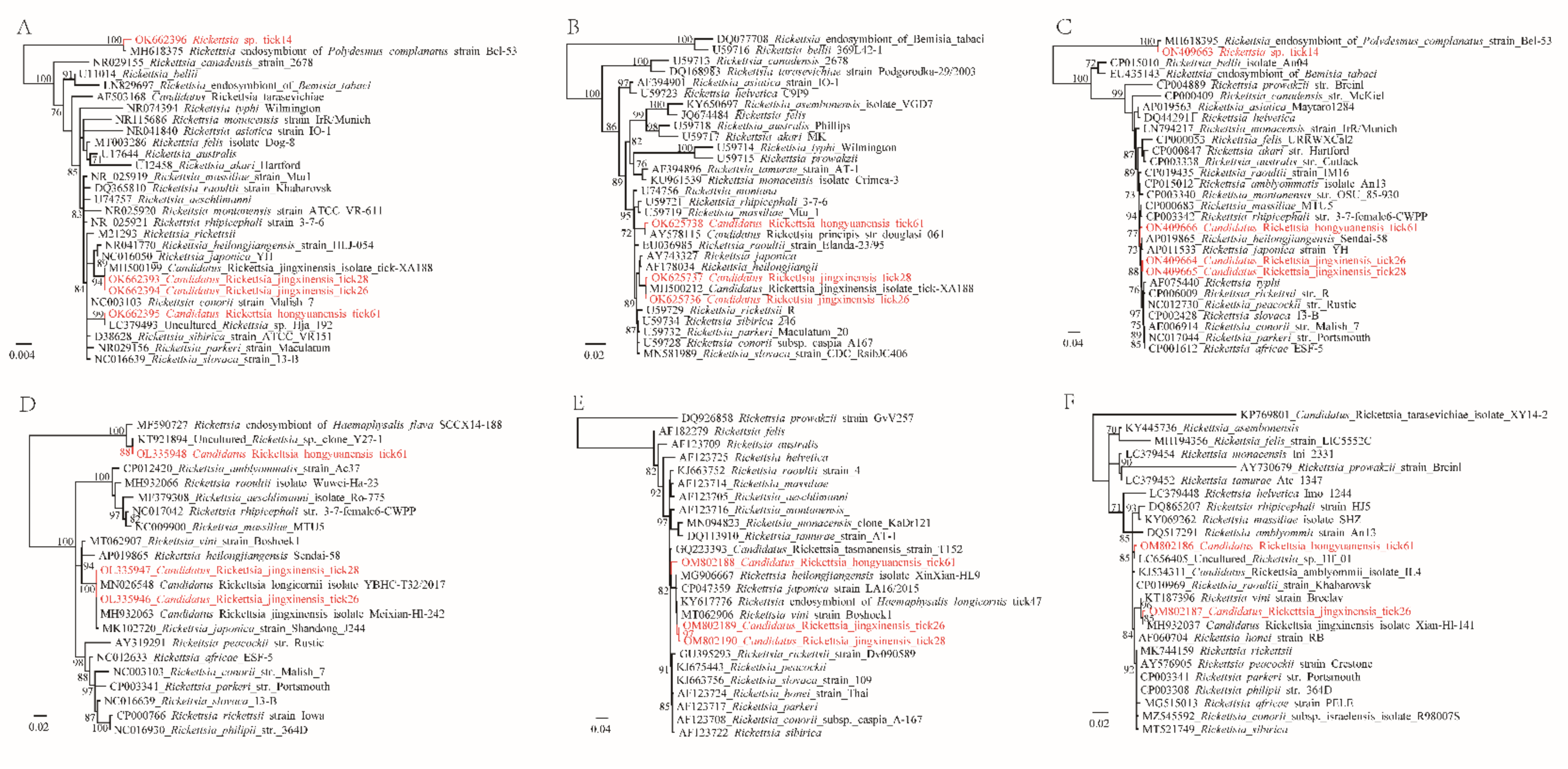
2.3. Amplification of Key Genes and Genetic and Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Processing
4.2. DNA Extraction
4.3. Species Identification of Ticks and Keds
4.4. PCR Detection and Amplification of Key Genes
4.5. Genetic and Phylogenetic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smetanová, K.; Schwarzová, K.; Kocianová, E. Detection of Anaplasma phagocytophilum, Coxiella burnetii, Rickettsia spp., and Borrelia burgdorferi s. l. in ticks, and wild-living animals in western and middle Slovakia. Ann. N. Y. Acad. Sci. 2006, 1078, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Todd, C.S.; Mansoor, G.F.; Buhler, C.; Rahimi, H.; Zekria, R.; Fernandez, S.; Mikhail, A.F.; Scott, P.T.; Yingst, S.L. Prevalence of zoonotic and vector-borne infections among Afghan National Army Recruits in Afghanistan. Vector Borne Zoonotic Dis. 2016, 16, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Minetti, C.; Pilotte, N.; Zulch, M.; Canelas, T.; Tettevi, E.J.; Veriegh, F.B.D.; Osei-Atweneboana, M.Y.; Williams, S.A.; Reimer, L.J. Field evaluation of DNA detection of human filarial and malaria parasites using mosquito excreta/feces. PLoS Negl. Trop. Dis. 2020, 14, e0008175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.; Li, F.; Liao, Y.; Shen, J.J.; Xu, J.M.; Chen, Y.Z.; Li, J.H.; Holmes, E.C.; Zhang, Y.Z. Epidemiology and diversity of Rickettsiales bacteria in humans and animals in Jiangsu and Jiangxi provinces, China. Sci. Rep. 2019, 9, 13176. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Lin, H.; Xu, X.; Ren, Q.; Aizezi, M.; Luo, J.; Luo, Y.; Ma, Z.; Chen, Z.; Tan, Y.; et al. Coxiella burnetii is widespread in ticks (Ixodidae) in the Xinjiang areas of China. BMC Vet. Res. 2020, 16, 317. [Google Scholar] [CrossRef]
- Ge, Y.; Yin, H.; Rikihisa, Y.; Pan, W.; Yin, H. Molecular Detection of Tick-Borne Rickettsiales in Goats and Sheep from Southeastern China. Vector Borne Zoonotic Dis. 2016, 16, 309–316. [Google Scholar] [CrossRef]
- Wang, H.; Yang, J.; Mukhtar, M.U.; Liu, Z.; Zhang, M.; Wang, X. Molecular detection and identification of tick-borne bacteria and protozoans in goats and wild Siberian roe deer (Capreolus pygargus) from Heilongjiang Province, northeastern China. Parasit Vectors 2019, 12, 296. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.H.; Chung, L.H.; Chien, C.H.; Tung, Y.J.; Wei, H.Y.; Yen, T.Y.; Shu, P.Y.; Wang, H.C. Human granulocytic anaplasmosis in Kinmen, an offshore island of Taiwan. PLoS Negl. Trop. Dis. 2019, 13, e0007728. [Google Scholar] [CrossRef]
- Woldehiwet, Z. Immune evasion and immunosuppression by Anaplasma phagocytophilum, the causative agent of tick-borne fever of ruminants and human granulocytic anaplasmosis. Vet. J. 2008, 175, 37–44. [Google Scholar] [CrossRef]
- Kazar, J. Coxiella burnetii infection. Ann. N. Y. Acad. Sci. 2005, 1063, 105–114. [Google Scholar] [CrossRef]
- Cho, H.C.; Hwang, S.; Kim, E.M.; Park, Y.J.; Shin, S.U.; Jang, D.H.; Chae, J.S.; Choi, K.S. Prevalence and molecular characterization of Coxiella burnetii in cattle, goats, and horses in the Republic of Korea. Vector Borne Zoonotic Dis. 2021, 21, 502–508. [Google Scholar] [CrossRef]
- González-Barrio, D.; Hagen, F.; Tilburg, J.J.; Ruiz-Fons, F. Coxiella burnetii genotypes in Iberian wildlife. Microb. Ecol. 2016, 72, 890–897. [Google Scholar] [CrossRef]
- Tokarevich, N.K.; Panferova, Y.A.; Freylikhman, O.A.; Blinova, O.V.; Medvedev, S.G.; Mironov, S.V.; Grigoryeva, L.A.; Tretyakov, K.A.; Dimova, T.; Zaharieva, M.M.; et al. Coxiella burnetii in ticks and wild birds. Ticks Tick-Borne Dis. 2019, 10, 377–385. [Google Scholar] [CrossRef]
- Marenzoni, M.L.; Stefanetti, V.; Papa, P.; Casagrande Proietti, P.; Bietta, A.; Coletti, M.; Passamonti, F.; Henning, K. Is the horse a reservoir or an indicator of Coxiella burnetii infection? Systematic review and biomolecular investigation. Vet. Microbiol. 2013, 167, 662–669. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Han, R.; Liu, G.; Shi, Y.; Luo, J.; Yin, H. Molecular survey and characterization of a novel Anaplasma species closely related to Anaplasma capra in ticks, northwestern China. Parasit Vectors 2016, 9, 603. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Yuan, D.; Guo, L.; Hou, W.; Mo, X.; Yin, J.; Yang, A.; Li, R. Molecular detection of Bartonella in ixodid ticks collected from yaks and plateau pikas (Ochotona curzoniae) in Shiqu County, China. BMC Vet. Res. 2020, 16, 235. [Google Scholar] [CrossRef]
- Hao, L.; Yuan, D.; Li, S.; Jia, T.; Guo, L.; Hou, W.; Lu, Z.; Mo, X.; Yin, J.; Yang, A.; et al. Detection of Theileria spp. in ticks, sheep keds (Melophagus ovinus), and livestock in the eastern Tibetan Plateau, China. Parasitol Res. 2020, 119, 2641–2648. [Google Scholar] [CrossRef]
- Thu, M.J.; Qiu, Y.; Matsuno, K.; Kajihara, M.; Mori-Kajihara, A.; Omori, R.; Monma, N.; Chiba, K.; Seto, J.; Gokuden, M.; et al. Diversity of spotted fever group rickettsiae and their association with host ticks in Japan. Sci. Rep. 2019, 9, 1500. [Google Scholar] [CrossRef] [Green Version]
- Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. Genome sequence of the endosymbiont Rickettsia peacockii and comparison with virulent Rickettsia rickettsii: Identification of virulence factors. PLoS ONE 2009, 4, e8361. [Google Scholar] [CrossRef]
- Mediannikov, O.; Sidelnikov, Y.; Ivanov, L.; Fournier, P.E.; Tarasevich, I.; Raoult, D. Far eastern tick-borne rickettsiosis: Identification of two new cases and tick vector. Ann. N. Y. Acad. Sci. 2006, 1078, 80–88. [Google Scholar] [CrossRef]
- Yang, J.; Tian., Z.; Liu, Z.; Niu, Q.; Han, R.; Li, Y.; Guan, G.; Liu, J.; Liu, G.; Luo, J.; et al. Novel spotted fever group rickettsiae in Haemaphysalis qinghaiensis ticks from Gansu, Northwest China. Parasit Vectors 2016, 9, 146. [Google Scholar] [CrossRef] [Green Version]
- Izzard, L.; Graves, S.; Cox, E.; Fenwick, S.; Unsworth, N.; Stenos, J. Novel rickettsia in ticks, Tasmania, Australia. Emerg Infect Dis. 2009, 15, 1654–1656. [Google Scholar] [CrossRef]
- Ogata, H.; Audic, S.; Renesto-Audiffren, P.; Fournier, P.E.; Barbe, V.; Samson, D.; Roux, V.; Cossart, P.; Weissenbach, J.; Claverie, J.M.; et al. Mechanisms of evolution in Rickettsia conorii and R. prowazekii. Science 2001, 293, 2093–2098. [Google Scholar] [CrossRef]
- El Karkouri, K.; Mediannikov, O.; Robert, C.; Raoult, D.; Fournier, P.E. Genome Sequence of the Tick-Borne Pathogen Rickettsia raoultii. Genome Announc. 2016, 4, e00157-16. [Google Scholar] [CrossRef] [Green Version]
- Akter, A.; Ooka, T.; Gotoh, Y.; Yamamoto, S.; Fujita, H.; Terasoma, F.; Kida, K.; Taira, M.; Nakadouzono, F.; Gokuden, M.; et al. Extremely Low Genomic Diversity of Rickettsia japonica Distributed in Japan. Genome Biol. Evol. 2017, 9, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 4, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Fournier, P.E.; Dumler, J.S.; Greub, G.; Zhang, J.; Wu, Y.; Raoult, D. Gene sequence-based criteria for identification of new rickettsia isolates and description of Rickettsia heilongjiangensis sp. nov. J. Clin. Microbiol. 2003, 41, 5456–5465. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Stanojević, M.; Stamenković, G.; Ilić, B.; Paunović, M.; Lu, M.; Pešić, B.; Đurić Maslovara, I.; Siljic, M.; Cirkovic, V.; et al. Insight into diversity of bacteria belonging to the order Rickettsiales in 9 arthropods species collected in Serbia. Sci. Rep. 2019, 9, 18680. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Kontsedalov, S.; Skaljac, M.; Brumin, M.; Sobol, I.; Czosnek, H.; Vavre, F.; Fleury, F.; et al. The transmission efficiency of tomato yellow leaf curl virus by the whitefly Bemisia tabaci is correlated with the presence of a specific symbiotic bacterium species. J. Virol. 2010, 84, 9310–9317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.P.; Tie, W.F.; Meng, S.; Li, D.; Wang, J.L.; Du, L.Y.; Xie, G.C. Extensive genetic diversity of Anaplasma bovis in ruminants in Xi’an, China. Ticks Tick Borne Dis. 2020, 11, 101477. [Google Scholar] [CrossRef] [PubMed]
- Panthawong, A.; Grieco, J.P.; Ngoen-Klan, R.; Chao, C.C.; Chareonviriyaphap, T. Detection of Anaplasma spp. and Bartonella spp. from wild-caught rodents and their ectoparasites in Nakhon Ratchasima Province, Thailand. J. Vector Ecol. 2020, 45, 241–253. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Guan, G.; Xie, J.; Luo, J.; Wang, S.; Wang, S.; Yin, H. First molecular survey and identification of Anaplasma spp. in white yaks (Bos grunniens) in China. Parasitology 2016, 143, 686–691. [Google Scholar] [CrossRef]
- Von Fricken, M.E.; Qurollo, B.A.; Boldbaatar, B.; Wang, Y.W.; Jiang, R.R.; Lkhagvatseren, S.; Koehler, J.W.; Moore, T.C.; Nymadawa, P.; Anderson, B.D.; et al. Genetic diversity of Anaplasma and Ehrlichia bacteria found in Dermacentor and Ixodes ticks in Mongolia. Ticks Tick Borne Dis. 2020, 11, 101316. [Google Scholar] [CrossRef]
- Liu, H.; Li, Q.; Zhang, X.; Li, Z.; Wang, Z.; Song, M.; Wei, F.; Wang, S.; Liu, Q. Characterization of rickettsiae in ticks in northeastern China. Parasit Vectors 2016, 9, 498. [Google Scholar] [CrossRef] [Green Version]
- Bang, M.S.; Kim, C.M.; Pyun, S.H.; Kim, D.M.; Yun, N.R. Molecular investigation of tick-borne pathogens in ticks removed from tick-bitten humans in the southwestern region of the Republic of Korea. PLoS ONE 2021, 16, e0252992. [Google Scholar] [CrossRef]
- Guo, W.P.; Wang, Y.H.; Lu, Q.; Xu, G.; Luo, Y.; Ni, X.; Zhou, E.M. Molecular detection of spotted fever group rickettsiae in hard ticks, northern China. Transbound Emerg. Dis. 2019, 66, 1587–1596. [Google Scholar] [CrossRef]
- Liu, H.; Liang, X.; Wang, H.; Sun, X.; Bai, X.; Hu, B.; Shi, N.; Wang, N.; Zhang, X.; Huang, L.; et al. Molecular evidence of the spotted fever group Rickettsiae in ticks from Yunnan Province, Southwest China. Exp. Appl. Acarol. 2020, 80, 339–348. [Google Scholar] [CrossRef]
- Takhampunya, R.; Korkusol, A.; Pongpichit, C.; Yodin, K.; Rungrojn, A.; Chanarat, N.; Promsathaporn, S.; Monkanna, T.; Thaloengsok, S.; Tippayachai, B.; et al. Metagenomic approach to characterizing disease epidemiology in a disease-endemic environment in Northern Thailand. Front. Microbiol. 2019, 10, 319. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Guo, W.B.; Pan, Y.S.; Jiang, B.G.; Du, C.H.; Que, T.C.; Zhan, L.; Wu, J.H.; Yu, M.H.; Cui, X.M.; et al. Detection of novel spotted fever group Rickettsiae (Rickettsiales: Rickettsiaceae) in ticks (Acari: Ixodidae) in Southwestern China. J. Med. Entomol. 2021, 58, 1363–1369. [Google Scholar] [CrossRef]
- Brenner, A.E.; Muñoz-Leal, S.; Sachan, M.; Labruna, M.B.; Raghavan, R. Coxiella burnetii and related tick endosymbionts evolved from pathogenic ancestors. Genome Biol. Evol. 2021, 13, evab108. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Lalzar, I.; Klasson, L. Distinctive genome reduction rates revealed by genomic analyses of two Coxiella-like endosymbionts in ticks. Genome Biol. Evol. 2015, 7, 1779–1796. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.K.; Owens, L. Infectivity, transmission and 16S rRNA sequencing of a rickettsia, Coxiella cheraxi sp. nov., from the freshwater crayfish Cherax quadricarinatus. Dis. Aquat. Organ. 2000, 41, 115–122. [Google Scholar] [CrossRef]
- Huang, B.; Shen, J. Classific Atlas of Parasites for Livestock and Poultry in China; China Agricultural Science and Technology Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Lu, M.; Tian, J.; Pan, X.; Qin, X.; Wang, W.; Chen, J.; Guo, W.; Li, K. Identification of Rickettsia spp., Anaplasma spp., and an Ehrlichia canis-like agent in Rhipicephalus microplus from Southwest and South-Central China. Ticks Tick Borne Dis. 2022, 13, 101884. [Google Scholar] [CrossRef]
- Duron, O.; Noël, V.; McCoy, K.D.; Bonazzi, M.; Sidi-Boumedine, K.; Morel, O.; Vavre, F.; Zenner, L.; Jourdain, E.; Durand, P.; et al. The recent evolution of a maternally-inherited endosymbiont of ticks led to the emergence of the Q fever pathogen, Coxiella burnetii. PLoS Pathog. 2015, 11, e1004892. [Google Scholar] [CrossRef] [Green Version]
- Borsoi, A.B.P.; Bitencourth, K.; de Oliveira, S.V.; Amorim, M.; Gazêta, G.S. Human Parasitism by Amblyomma parkeri Ticks Infected with Candidatus Rickettsia paranaensis, Brazil. Emerg. Infect Dis. 2019, 25, 2339–2341. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Delsuc, F.; Dufayard, J.F.; Gascuel, O. Estimating maximum likelihood phylogenies with PhyML. Methods Mol. Biol. 2009, 537, 113–137. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Haemaphysalis qinghaiensis | Melophagus ovinus | Bos mutus | Capra hircus |
|---|---|---|---|---|
| Anaplasma bovis | 8.83% (37/419) | 0.00% (0/106) | 45.71% (64/140) | 58.93% (33/56) |
| Candidatus Rickettsia jingxinensis | 0.48% (2/419) | 0.00% (0/106) | 0.00% (0/140) | 0.00% (0/56) |
| Candidatus Rickettsia hongyuanensis | 0.24% (1/419) | 0.00% (0/106) | 0.00% (0/140) | 0.00% (0/56) |
| Rickettsia sp. tick14 | 0.24% (1/419) | 0.00% (0/106) | 0.00% (0/140) | 0.00% (0/56) |
| Coxiella spp. | 3.34% (14/419) | 1.89% (2/106) | 0.71% (1/140) | 0.00% (0/56) |
| Coxiella sp. tick8 | 0.24% (1/419) | 0.00% (0/106) | 0.00% (0/140) | 0.00% (0/56) |
| Coxiella-like bacterium | 0.00% (0/419) | 0.00% (0/106) | 0.00% (0/140) | 1.79% (1/56) |
| Total | 13.37% (56/419) | 1.89% (2/106) | 46.43% (65/140) | 60.71% (34/56) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, M.; Tian, J.; Zhao, H.; Jiang, H.; Qin, X.; Wang, W.; Li, K. Molecular Survey of Vector-Borne Pathogens in Ticks, Sheep Keds, and Domestic Animals from Ngawa, Southwest China. Pathogens 2022, 11, 606. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050606
Lu M, Tian J, Zhao H, Jiang H, Qin X, Wang W, Li K. Molecular Survey of Vector-Borne Pathogens in Ticks, Sheep Keds, and Domestic Animals from Ngawa, Southwest China. Pathogens. 2022; 11(5):606. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050606
Chicago/Turabian StyleLu, Miao, Junhua Tian, Hongqing Zhao, Hai Jiang, Xincheng Qin, Wen Wang, and Kun Li. 2022. "Molecular Survey of Vector-Borne Pathogens in Ticks, Sheep Keds, and Domestic Animals from Ngawa, Southwest China" Pathogens 11, no. 5: 606. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050606