Preparation of Chicken Anemia Virus (CAV) Virus-Like Particles and Chicken Interleukin-12 for Vaccine Development Using a Baculovirus Expression System


Abstract
:1. Introduction
2. Results
2.1. Expression of Recombinant Proteins
2.2. Virus-Like Particles (VLPs) Formed by Recombinant VP1 Protein
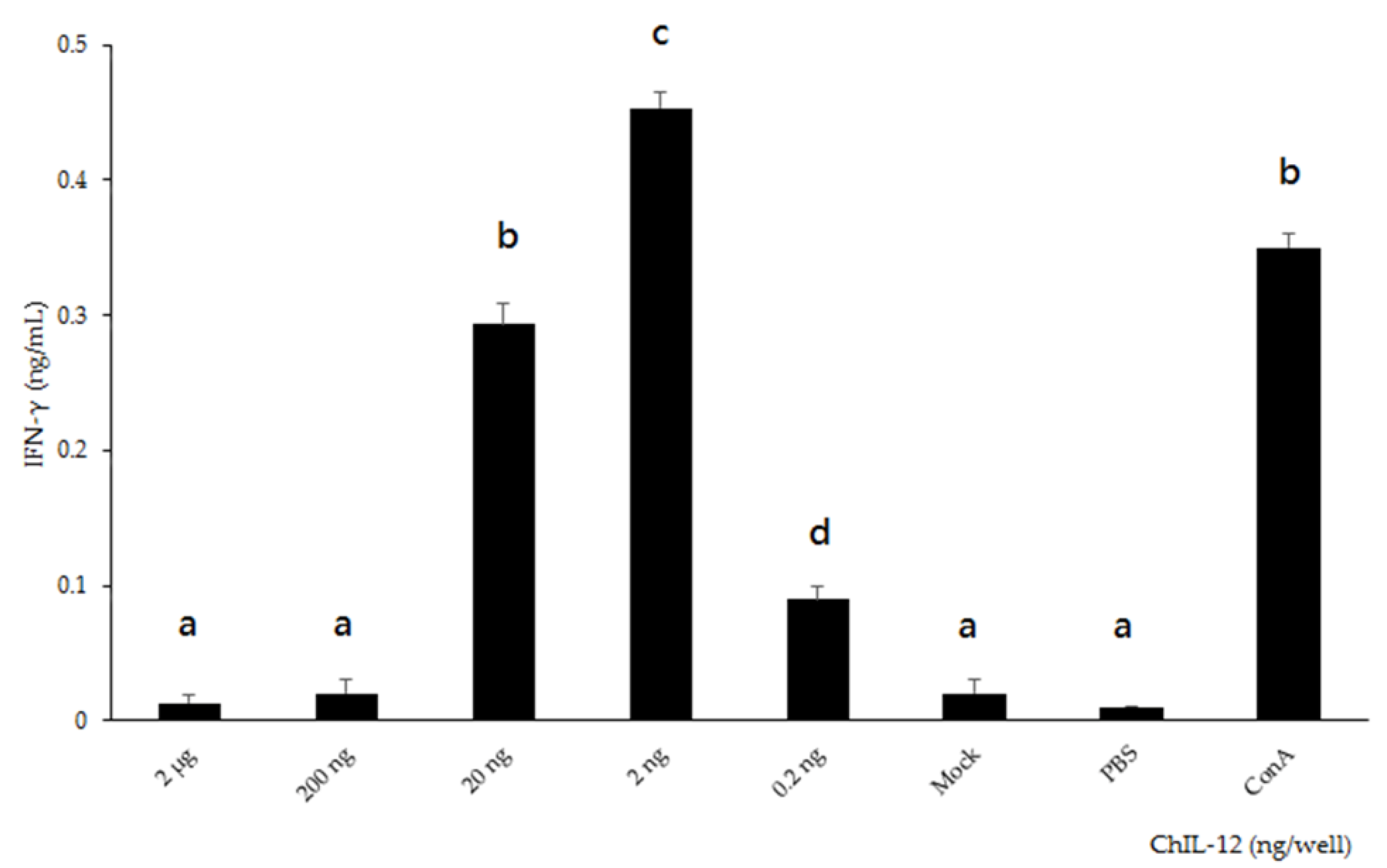
2.3. Bioactivity of rchIL-12
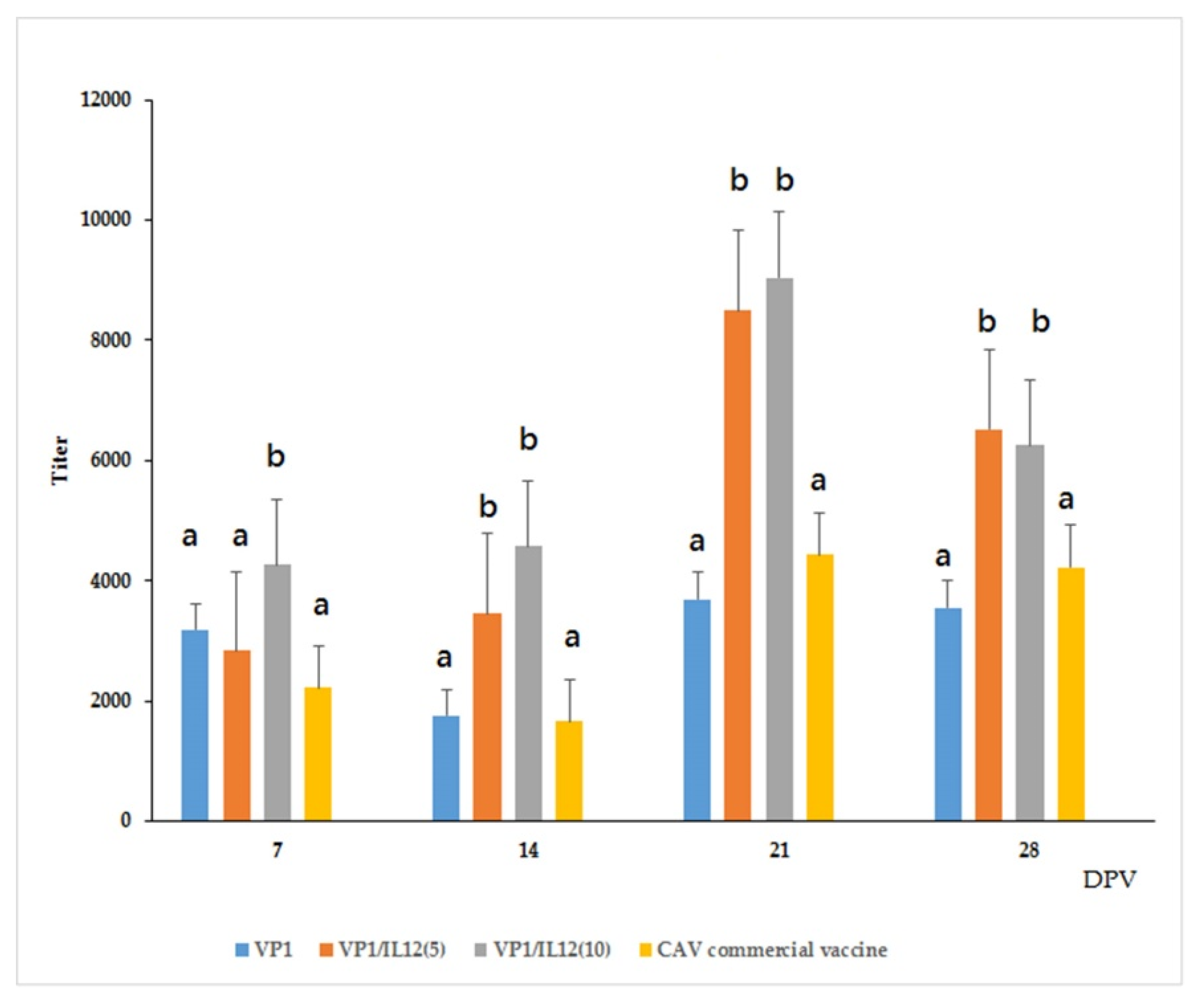
2.4. Immune Response of VLP-Vaccinated Chickens
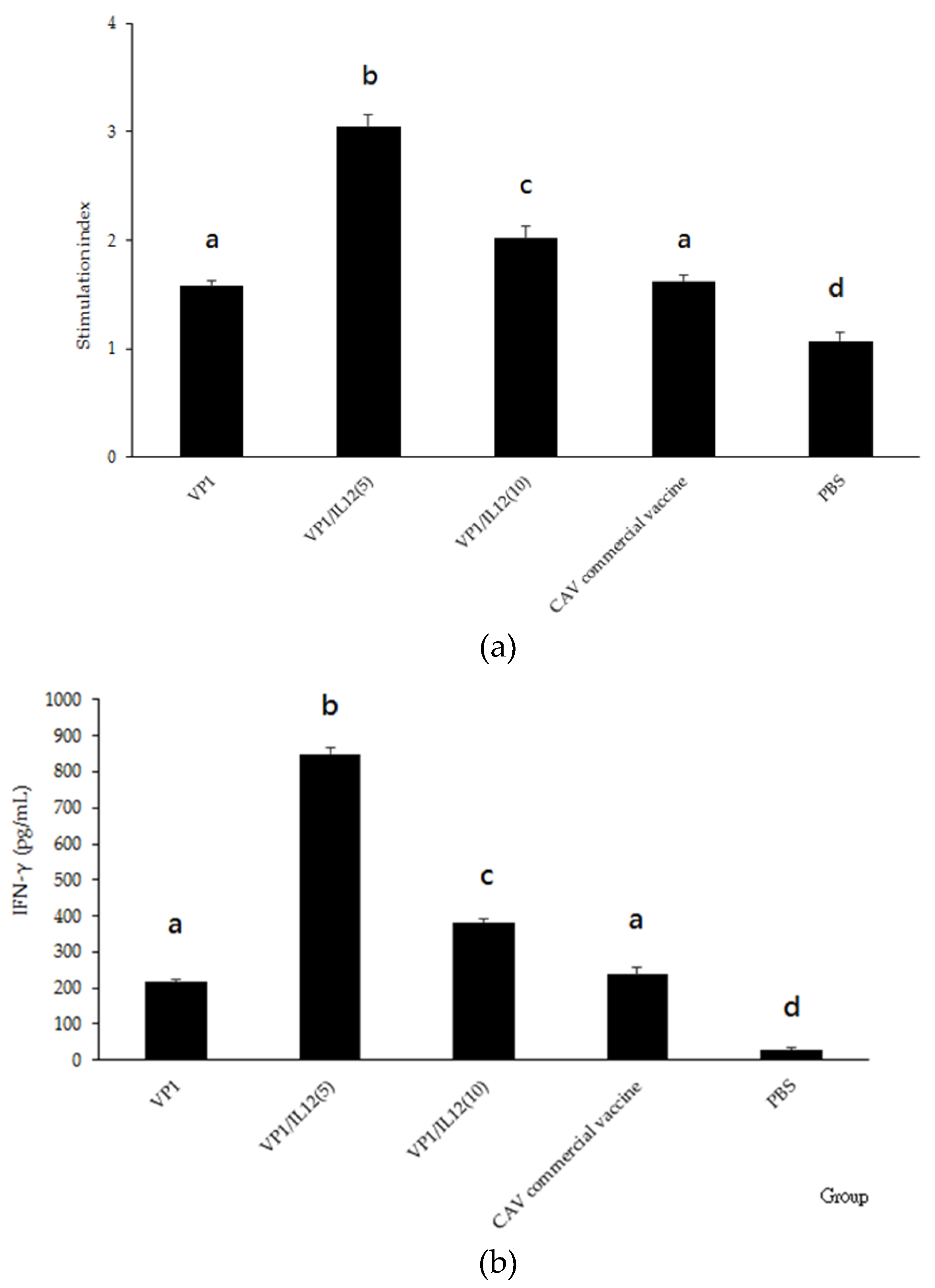
2.5. Antigen-Specific Splenocyte Proliferation and IFN-γ Secretion Assays
3. Discussion
4. Materials and Methods
4.1. Gene Cloning and Virus
4.2. Animals
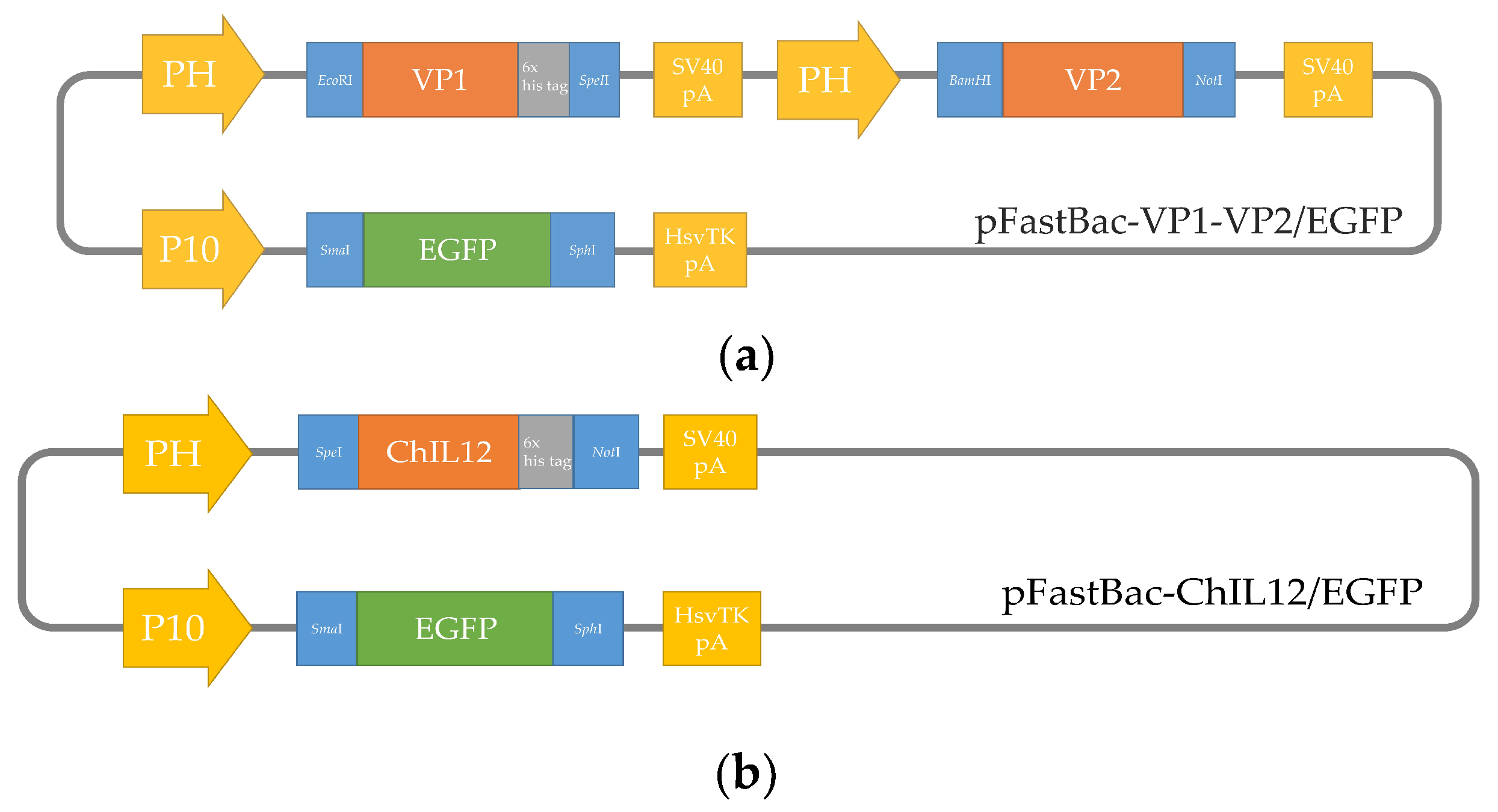
4.3. Construction of Recombinant Baculoviruses
4.4. Western Blot Analysis
4.5. Indirect Immunofluorescence Assay (IFA)
4.6. Purification of Recombinant VP1 and chIL-12
4.7. CAV VLP Observation by Electron Microscope
4.8. Sf9-Expressed chIL-12 Bioactivity Assay
4.9. Immunization of Chickens
4.10. Splenocyte Proliferation Assay
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schat, K.A.; van Santen, V.L. Chicken Infectious Anemia. In Diseases of Poultry, 13th ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 248–264. [Google Scholar]
- Van Santen, V.L.; Li, L.; Hoerr, F.J.; Lauerman, L.H. Genetic characterization of chicken anemia virus from commercial broiler chickens in Alabama. Avian Dis. 2001, 45, 373–388. [Google Scholar] [CrossRef]
- Smyth, J.A.; Moett, D.A.; Connor, T.J.; McNulty, M.S. Chicken anaemia virus inoculated by the oral route causes lymphocyte depletion in the thymus in 3-week-old and 6-week-old chickens. Avian Pathol. 2006, 35, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Breitbart, M.; Harrach, B.; Segales, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the taxonomy of the family Circoviridae: Establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Claessens, J.A.; Schrier, C.C.; Mockett, A.P.; Jagt, E.H.; Sondermeijer, P.J. Molecular cloning and sequence analysis of the genome of chicken anaemia agent. J. Gen. Virol. 1991, 72, 2003–2006. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, H.; Kling, S.; Lurz, R.; Tischer, I.; von Bülow, V. Morphological characterization of chicken anaemia agent (CAA). Arch. Virol. 1989, 109, 115–120. [Google Scholar] [CrossRef]
- McNulty, M.S.; Curran, W.L.; Todd, D.; Mackie, D.P. Chicken anemia agent: An electron microscopic study. Avian Dis. 1990, 34, 736–743. [Google Scholar] [CrossRef]
- Noteborn, M.H.; de Boer, G.F.; van Roozelaar, D.J.; Karreman, C.; Kranenburg, O.; Vos, J.G.; Jeurissen, S.H.; Hoeben, R.C.; Zantema, A.; Koch, G. Characterization of cloned chicken anemia virus DNA that contains all elements for the infectious replication cycle. J. Virol. 1991, 65, 3131–3139. [Google Scholar]
- Noteborn, M.H.; Verschueren, C.A.; Koch, G.; van der Eb, A.J. Simultaneous expression of recombinant baculovirus-encoded chicken anaemia virus (CAV) proteins VP1 and VP2 is required for formation of the CAV-specific neutralizing epitope. J. Gen. Virol. 1998, 79, 3073–3077. [Google Scholar] [CrossRef]
- Peters, M.A.; Jackson, D.C.; Crabb, B.S.; Browning, G.F. Chicken anemia virus VP2 is a novel dual specificity protein phosphatase. J. Biol. Chem. 2002, 277, 39566–39573. [Google Scholar] [CrossRef]
- Koch, G.; van Roozelaar, D.J.; Verschueren, C.A.; van der Eb, A.J.; Noteborn, M.H. Immunogenic and protective properties of chicken anaemia virus proteins expressed by baculovirus. Vaccine 1995, 13, 763–770. [Google Scholar] [CrossRef]
- Noteborn, M.H.; Todd, D.; Verschueren, C.A.; de Gauw, H.W.; Curran, W.L.; Veldkamp, S.; Douglas, A.J.; McNult, M.S.; van der Eb, A.J.; Koch, G.A. Single chicken anemia virus protein induces apoptosis. J. Virol. 1994, 68, 346–351. [Google Scholar] [PubMed]
- McNulty, M.S. Chicken amaemia agent: A review. Avian Pathol. 1991, 20, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Todd, D.; Creelan, J.L.; Connor, T.J.; Ball, N.W.; Scott, A.N.; Meehan, B.M.; McKenna, G.F.; McNulty, M.S. Investigation of the unstable attenuation exhibited by a chicken anaemia virus isolate. Avian Pathol. 2003, 32, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Vaziry, A.; Silim, A.; Bleau, C.; Frenette, D.; Lamontagne, L. Chicken infectious anaemia vaccinal strain persists in the spleen and thymus of young chicks and induces thymic lymphoid cell disorders. Avian Pathol. 2011, 40, 377–385. [Google Scholar] [CrossRef]
- Lee, M.-S.; Hseu, Y.-C.; Lai, G.-H.; Chang, W.-T.; Chen, H.-J.; Huang, C.-H.; Lee, M.-S.; Wang, M.-Y.; Kao, J.-Y.; You, B.-J.; et al. High yield expression in a recombinant E. coli of a codon optimized chicken anemia virus capsid protein VP1 useful for vaccine development. Microb. Cell Fact. 2011, 10, 56. [Google Scholar] [CrossRef]
- Lacorte, C.; Lohuis, H.; Goldbach, R.; Prins, M. Assessing the expression of chicken anemia virus proteins in plants. Virus Res. 2007, 129, 80–86. [Google Scholar] [CrossRef]
- Moeini, H.; Omar, A.R.; Rahim, R.A.; Yusoff, K. Development of a DNA vaccine against chicken anemia virus by using a bicistronic vector expressing VP1 and VP2 proteins of CAV. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 227–236. [Google Scholar] [CrossRef]
- Moeini, H.; Omar, A.R.; Rahim, R.A.; Yusoff, K. Improving the potency of DNA vaccine against Chicken Anemia Virus (CAV) by fusing VP1 protein of CAV to Marek’s Disease Virus (MDV)Type-1 VP22 protein. Virol. J. 2011, 8, 119. [Google Scholar] [CrossRef]
- Pages-Mante, A.; Saubi, N.; Artigas, C.; Espuna, E. Experimental evaluation of an inactivated vaccine against chicken anaemia virus. Avian Pathol. 1997, 26, 721–729. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, B.; Liu, Y.; Chen, W.; Dai, Z.; Bi, Y.; Xie, Q. Assessing the efficacy of an inactivated chicken anemia virus vaccine. Vaccine 2015, 33, 1916–1922. [Google Scholar] [CrossRef]
- Asif, M.; Jenkins, K.A.; Hilton, L.S.; Kimpton, W.G.; Bean, A.G.; Lowenthal, J.W. Cytokines as adjuvants for avian vaccines. Immunol. Cell Biol. 2004, 82, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.-H.; Li, H.-P.; Lien, Y.-Y.; Wu, M.-L.; Chaung, H.-C. Adjuvant effects of chicken interleukin-18 in avian Newcastle disease vaccine. Vaccine 2010, 28, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Rengarajan, J.; Szabo, S.J.; Glimcher, L.H. Transcriptional regulation of Th1/Th2 polarization. Immunol. Today 2000, 21, 479. [Google Scholar] [CrossRef]
- Degen, W.G.; van Daal, N.; van Zuilekom, H.I.; Burnside, J.; Schijns, V.E. Identification and molecular cloning of functional chicken IL-12. J. Immunol. 2004, 172, 4371–4380. [Google Scholar] [CrossRef]
- Medrano, G.; Dolan, M.C.; Stephens, N.T.; McMickle, A.; Erf, G.; Radin, D.; Cramer, C.L. Efficient plant-based production of chicken interleukin-12 yields a strong immunostimulatory cytokine. J. Interferon Cytokine Res. 2010, 30, 143–154. [Google Scholar] [CrossRef]
- Su, B.S.; Yin, H.S.; Shien, J.H.; Chiu, H.H.; Lee, L.H. Production of biologically active chicken interleukin (IL)-12 and IL-18 synthesized by the recombinant fowlpox virus. Process Biochem. 2010, 45, 1057–1064. [Google Scholar] [CrossRef]
- Ha, S.; Chang, J.; Song, M.; Suh, Y.; Jin, H.; Lee, C.; Nam, G.; Choi, G.; Choi, K.; Lee, S.; et al. Engineering N-glycosylation mutations in IL-12 enhances sustained cytotoxic T lymphocyte responses for DNA immunization. Nat. Biotechnol. 2002, 20, 381–386. [Google Scholar] [CrossRef]
- Pallister, J.; Fahey, K.J.; Sheppard, M. Cloning and sequencing of the chicken anaemia virus (CAV) ORF-3 gene, and the development of an ELISA for the detection of serum antibody to CAV. Vet. Microbiol. 1994, 39, 167–178. [Google Scholar] [CrossRef]
- Lee, M.-S.; Lien, Y.-Y.; Feng, S.-H.; Huang, R.-L.; Tsai, M.-C.; Chang, W.-T.; Chen, H.-J. Production of chicken anemia virus (CAV) VP1 and VP2 protein expressed by recombinant Escherichia coli. Process Biochem. 2009, 44, 390–395. [Google Scholar] [CrossRef]
- Sawant, P.M.; Dhama, K.; Rawool, D.B.; Wani, M.Y.; Tiwari, R.; Singh, S.D.; Singh, R.K. Development of a DNA vaccine for chicken infectious anemia and its immunogenicity studies using high mobility group box 1 protein as a novel immunoadjuvant indicated induction of promising protective immune responses. Vaccine 2015, 33, 333–340. [Google Scholar] [CrossRef]
- Canal, C.W.; Ferreira, D.J.; Macagnan, M.; Fallavena, L.C.B.; Moraes, H.L.S.; Wald, V.B. Prevalance of antibodies against chicken anemia virus (CAV) in breeders in Southern Brazil. Pesqui. Vet. Bras. 2004, 24, 89–92. [Google Scholar] [CrossRef]
- Roussan, D.A. Serological survey on the prevalence of chicken infectious anemia virus in commercial broiler chicken flocks in Northern Jordan. Int. J. Poult. Sci. 2006, 5, 544–546. [Google Scholar]
- Metzger, D.W. IL-12 as an adjuvant for the enhancement of protective humoral immunity. Expert Rev. Vaccine 2009, 8, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Sturmhoefel, K.; Lee, K.; Toole, M.O.; Swiniarski, H.M.; Dorner, A.J.; Wolf, S.F. Interleukin 12 as vaccine adjuvant. Res. Immunol. 1998, 149, 37–39. [Google Scholar] [CrossRef]
- Li, X.; Li, P.; Cao, L.; Bai, Y.; Chen, H.; Liu, H.; Ren, X.; Li, G. Porcine IL-12 plasmid as an adjuvant improve the cellular and humoral immune responses of DNA vaccine targeting transmissible gastroenteritis virus spike gene in a miuse model. J. Vet. Med. Sci. 2019, 81, 1438–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, B.S.; Chiu, H.H.; Lin, C.C.; Shien, J.H.; Yin, H.S.; Lee, L.H. Adjuvant active of chicken interleukin-12 co-administrated with infectious bursal disease virus recombinant VP2 antigen in chickens. Vet. Immunol. Immunolpathol. 2011, 139, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Noad, R.; Roy, P. Virus-like particles as immunogens. Trends Microbiol. 2003, 11, 438–444. [Google Scholar] [CrossRef]
- Ou, S.C.; Lin, H.L.; Liu, P.C.; Huang, H.J.; Lee, M.S.; Lien, Y.Y.; Tsai, Y.L. Epidemiology and molecular characterization of chicken anaemia virus from commercial and native chickens in Taiwan. Transbound. Emerg. Dis. 2018, 65, 1493–1501. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Sequence (5’→3’) | Amplicon Size (bp) | Restrict Enzyme |
|---|---|---|---|---|
| CAV-VP1 1 | BVP1-EcoRI-F | GAATTCATGGCTCGTCGCGCTC | 1380 | EcoRI |
| BVP1-SpeI-R | ACTAGTTCAGTGGTGATGATGGTGATGA | SpeI | ||
| CAV-VP2 1 | BVP2-BamHI-F | GGATCCATGCACGGTAACGGTGG | 665 | BamHI |
| BVP2-NotI-R | GCGGCCGCTCACACGATG | NotI | ||
| Chicken IL-12 1 | BChIL-12-XhoI-F | CTCGAGATGTCCCACCTGCTGTT | 1488 | XhoI |
| BChIL-12-KpnI-R | GGTACCTTAGTGATGATGGTGGTGGTG | KpnI | ||
| EGFP | EGFP-SmaI-F | CCCGGGATGGTGAGCAAGGGCGAGGA | 720 | SmaI |
| EGFP-SphI-R | GCATGCTTACTTGTACAGCTCGTCCATGCCGA | SphI |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, T.-Y.; Liu, Y.-C.; Hsu, Y.-C.; Chang, P.-C.; Hsieh, M.-K.; Shien, J.-H.; Ou, S.-C. Preparation of Chicken Anemia Virus (CAV) Virus-Like Particles and Chicken Interleukin-12 for Vaccine Development Using a Baculovirus Expression System. Pathogens 2019, 8, 262. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8040262
Tseng T-Y, Liu Y-C, Hsu Y-C, Chang P-C, Hsieh M-K, Shien J-H, Ou S-C. Preparation of Chicken Anemia Virus (CAV) Virus-Like Particles and Chicken Interleukin-12 for Vaccine Development Using a Baculovirus Expression System. Pathogens. 2019; 8(4):262. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8040262
Chicago/Turabian StyleTseng, Ta-Yuan, Yee-Chen Liu, Yu-Chen Hsu, Poa-Chun Chang, Ming-Kun Hsieh, Jui-Hung Shien, and Shan-Chia Ou. 2019. "Preparation of Chicken Anemia Virus (CAV) Virus-Like Particles and Chicken Interleukin-12 for Vaccine Development Using a Baculovirus Expression System" Pathogens 8, no. 4: 262. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8040262