Frequency of Resistance to Benzimidazoles of Haemonchus contortus Helminths from Dairy Sheep, Goats, Cattle and Buffaloes in Greece

 , , , and
, , , and 
Abstract
:1. Introduction
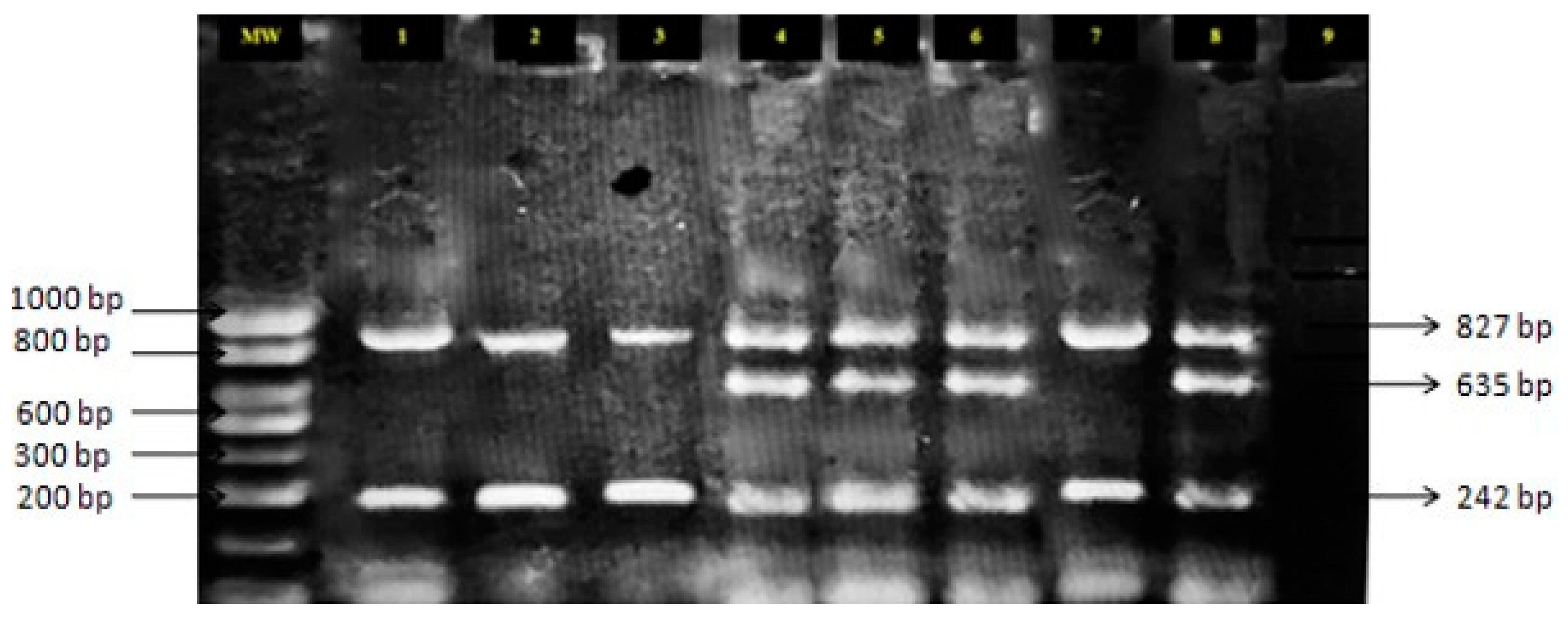
2. Results
3. Discussion
4. Materials and Methods
4.1. Helminths
4.2. DNA Extraction—Allele Specific PCR
4.3. Data Management and Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Familton, A.S.; McAnulty, R.W. Life cycles and development of nematode parasites of ruminants. In Sustainable Control of Internal Parasites in Ruminants; Barrell, G.K., Ed.; Lincoln University: Canterbury, New Zealand, 1997; pp. 67–80. [Google Scholar]
- Perry, B.D.; Randolph, T.F. Improving the assessment of the economic impact of parasitic diseases and of their control in production animals. Vet. Parasitol. 1999, 84, 145–168. [Google Scholar] [CrossRef]
- Stromberg, B.E.; Gasbarre, L.C. Gastrointestinal nematode control programs with an emphasis on cattle. Vet. Clin. N. Am. Food Anim. Pract. 2006, 22, 543–565. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, E.; Arsenos, G.; Sotiraki, S.; Deligiannis, C.; Lainas, T.; Zygogiannis, D. The epizootiology of gastrointestinal nematode parasites in Greek dairy breeds of sheep and goats. Small Rumin. Res. 2003, 47, 193–202. [Google Scholar] [CrossRef]
- Laurenson, Y.; Bishop, S.C.; Kyriazakis, I. In silico exploration of the mechanisms that underlie parasite-induced anorexia in sheep. Br. J. Nutr. 2011, 106, 1023–1039. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.M.; Waghorn, T.S.; Leathwick, D.M.; Candy, P.M.; Oliver, A.M.B.; Watson, T.G. The production cost of anthelmintic resistance in lambs. Vet. Parasitol. 2012, 186, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Lichtenfels, J.R.; Pilitt, P.A.; Hoberg, E.P. New morphological characters for identifying individual specimens of Haemonchus spp. (Nematoda: Trichostrongyloidea) and a key to species in ruminants of North America. J. Parasitol. 1994, 80, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Hoberg, E.R.; Lichtenfels, J.R.; Gibbons, L. Phylogeny for species of Haemonchus (nematoda: Trichostrongyloidea): Considerations of their evolutionary history and global biogeography among camelidae and pecora (artiodactyla). J. Parasitol. 2004, 90, 1085–1102. [Google Scholar] [CrossRef] [Green Version]
- Achi, Y.L.; Zinsstag, J.; Yao, K.; Dorchies, P.; Jacquiet, P. Host specificity of Haemonchus spp. for domestic ruminants in the savanna in northern Ivory Coast. Vet. Parasitol. 2003, 116, 151–158. [Google Scholar] [CrossRef]
- Akkari, H.; Jebali, J.; Gharbi, M.; Mhadhbi, M.; Awadi, S.; Darghouth, M.A. Epidemiological study of sympatric Haemonchus species and genetic characterization of Haemonchus contortus in domestic ruminants in Tunisia. Vet. Parasitol. 2013, 193, 118–125. [Google Scholar] [CrossRef]
- Waller, P.J. Anthelmintic resistance. Vet. Parasitol. 1997, 72, 391–412. [Google Scholar] [CrossRef]
- Cudekova, P.; Varady, M.; Dolinska, M.; Konigova, A. Phenotypic and genotypic characterization of benzimidazole susceptible and resistant isolates of Haemonchus contortus. Vet. Parasitol. 2010, 172, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.M.; Vidyashankar, A.N. An inconvenient truth: Global warming and anthelmintic resistance. Vet. Parasitol. 2012, 186, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.M. Drug resistance in nematodes of veterinary importance: A status report. Trends Parasitol. 2004, 20, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Elard, L.; Cabaret, J.; Humbert, J.F. PCR diagnosis of benzimidazole-susceptibility or -resistance in natural populations of the small ruminant parasite, Teladorsagia circumcincta. Vet. Parasitol. 1999, 80, 231–237. [Google Scholar] [CrossRef]
- Gallidis, E.; Angelopoulou, K.; Papadopoulos, E. First identification of benzimidazole resistant Haemonchus contortus in sheep in Greece. Small Rumin. Res. 2012, 106, 27–29. [Google Scholar] [CrossRef]
- Ali, Q.; Rashid, I.; Shabbir, M.Z.; Rahman, A.U.; Shahzad, K.; Ashraf, K.; Sargison, N.D.; Chaudry, U. Emergence and the spread of the F200Y benzimidazole resistance mutation in Haemonchus contortus and Haemonchus placei from buffalo and cattle. Vet. Parasitol. 2018, 285, 48–54. [Google Scholar] [CrossRef]
- Cazajous, T.; Prevot, F.; Kerbiriou, A.; Milhes, M.; Grisez, C.; Tropee, A.; Godart, C.; Aragon, A.; Jacquiet, P. Multiple resistance to ivermectin and benzimidazole of Haemonchus contortus population in a sheep flock from mainland France, first report. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 103–105. [Google Scholar] [CrossRef]
- Baiak, B.; Lehnen, C.; Rocha, R. Anthelmintic resistance of injectable macrocyclic lactones in cattle: A systematic review and meta-analysis. Braz. J. Vet. Parasitol. 2019, 28, 59–67. [Google Scholar] [CrossRef]
- Borgsteede, F.H.; Dercksen, D.D.; Huijbers, R. Doramectin and albendazole resistance in sheep in the Netherlands. Vet. Parasitol. 2007, 144, 180–183. [Google Scholar] [CrossRef]
- Scheuerle, M.C.; Mahling, M.; Pfister, K. Anthelminthic resistance of Haemonchus contortus in small ruminants in Switzerland and Southern Germany. Wien Klin. Wochenschr. 2009, 121, 46–49. [Google Scholar] [CrossRef]
- Mooney, L.; Good, B.; Hanrahan, J.P.; Mulcahy, G.; de Waal, T. The comparative efficacy of four anthelmintics against a natural acquired Fasciola hepatica infection in hill sheep flock in the west of Ireland. Vet. Parasitol. 2009, 164, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Kwa, M.S.; Veenstra, J.G.; Roos, M.H. Benzimidazole resistance in Haemonchus contortus is correlated with a conserved mutation at amino acid 200 in β tubulin isotype 1. Mol. Biochem. Parasitol. 1994, 63, 299–303. [Google Scholar] [CrossRef]
- Silvestre, A.; Cabaret, J. Mutation in position 167 of isotype I beta-tubulin gene of trichostrongylid nematodes: Role in BZ resistance? Mol. Biochem. Parasitol. 2002, 120, 297–300. [Google Scholar] [CrossRef]
- Ghisi, M.; Kaminsky, R.; Maser, P. Phenotyping and genotyping of Haemonchus contortus isolates reveals a new putative candidate mutation for BZ resistance in nematodes. Vet. Parasitol. 2007, 144, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, A.; Humbert, J.F. Diversity of benzimidazole-resistance alleles in populations of small ruminant parasites. Int. J. Parasitol. 2002, 32, 921–928. [Google Scholar] [CrossRef]
- Silvestre, A.; Sauve, C.; Cortet, J.; Cabaret, J. Contrasting genetic structures of two parasitic nematodes, determined on the basis of neutral microsatellite markers and selected anthelmintic resistance markers. Mol. Ecol. 2009, 18, 5086–5100. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, U.; Redman, E.M.; Raman, M.; Gilleard, J.S. Genetic evidence for the spread of a benzimidazole resistance mutation across southern India from a single origin in the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2015, 45, 721–728. [Google Scholar] [CrossRef]
- Chaudhry, U.; Redman, E.M.; Ashraf, K.; Shabbir, M.Z.; Rashid, M.I.; Ashraf, S.; Gilleard, J.S. Microsatellite marker analysis of Haemonchus contortus populations from Pakistan suggests that frequent benzimidazole drug treatment does not result in a reduction of overall genetic diversity. Parasites Vec. 2016, 9, 349. [Google Scholar] [CrossRef] [Green Version]
- Atanásio-Nhacumbe, A.; Lambert, S.M.; Souza, B.M.; Ayres, M.C.C. Molecular detection of benzimidazole resistance levels associated with F167Y and F200Y polymorphisms in Haemonchus contortus of goats from Mozambique. Parasitol. Res. 2019, 118, 245–253. [Google Scholar] [CrossRef]
- Gasbarre, L.C.; Smith, L.L.; Lichtenfels, J.R.; Pilitt, P.A. The identification of cattle nematode parasites resistant to multiple classes of anthelmintics in a commercial cattle population in the US. Vet. Parasitol. 2009, 166, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, I.A.; Leathwick, D.M. Anthelmintic resistance in nematode parasites of cattle: A global issue? Trends Parasitol. 2011, 27, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Brasil, B.S.; Nunes, R.L.; Bastianetto, E.; Drummond, M.G.; Carvalho, D.C.; Leite, R.C.; Molento, M.B.; Oliveira, D.A. Genetic diversity patterns of Haemonchus placei and Haemonchus contortus populations isolated from domestic ruminants in Brazil. Int. J. Parasitol. 2012, 42, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Ademola, I.O.; Krucken, J.; Ramunke, S.; Demeler, J.; von Samson-Himmelstjerna, G. Absence of detectable benzimidazole-resistance associated alleles in Haemonchus placei in cattle in Nigeria revealed by pyrosequencing of beta-tubulin isotype 1. Parasitol. Res. 2015, 114, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, U.; Redman, E.; Abbas, M.; Muthusamy, R.; Ashraf, K.; Gilleard, J. The presence of benzimidazole resistance mutations in Haemonchus placei of US cattle. Int. J. Parasitol. 2015, 45, 149–159. [Google Scholar] [CrossRef]
- Muñiz-Lagunes, A.; González-Garduño, R.; López-Arellano, M.E.; Ramírez-Valverde, R.; Ruíz-Flores, A.; García-Muñiz, G.; Ramírez-Vargas, G.; Mendoza-de Gives, P.; Torres-Hernández, G. Anthelmintic resistance in gastrointestinal nematodes from grazing beef cattle in Campeche State, Mexico. Trop. Anim. Health Prod. 2015, 47, 1049–1054. [Google Scholar] [CrossRef]
- Lanusse, C.E.; Alvarez, L.I.; Lifschitz, A.L. Gaining insights into the pharmacology of anthelmintics using Haemonchuscontortus as a model nematode. Adv. Parasitol. 2016, 93, 465–518. [Google Scholar]
- McKellar, Q.A.; Jackson, F. Veterinary anthelmintics: Old and new. Trends Parasitol. 2004, 20, 456–461. [Google Scholar] [CrossRef]
- Papadopoulos, E.; Himonas, C.; Gerald, C.C. Drought and flock isolation may enhance the development of anthelmintic resistance in nematodes. Vet. Parasitol. 2001, 97, 253–259. [Google Scholar] [CrossRef]
- Karteri, O.; Papadopoulos, E.; Vasileiou, N.G.C.; Mavrogianni, V.S.; Fthenakis, G.C. Practices in controlling helminth infections of small ruminant by veterinarians in clinical practice in Greece. In Proceedings of the 25th International Conference of the World Association for the Advancement of Veterinary Parasitology, Liverpool, UK, 16–20 August 2015; p. 303. [Google Scholar]
- Hoste, H.; Torres-Acosta, J.F.; Paolini, V.; Aguilar-Caballero, A.; Etter, E.; Lefrileux, Y.; Chartier, C.; Broqua, C. Interactions between nutrition and gastrointestinal infections with parasitic nematodes in goats. Small Rumin. Res. 2005, 60, 141–151. [Google Scholar] [CrossRef]
- Jackson, F. Anthelmintic resistance-the state of play. Br. Vet. J. 1993, 149, 123–138. [Google Scholar] [CrossRef]
- Papadopoulou, A. Sustainable Distribution of the Use of Pasture Lands to Animal Farmers in Mountainous and Plain Areas in Association with their Productivity and the Nutritional Requirements of Farm Animals. The Case of Lagkadas Local Authority. Master’s Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2015. [Google Scholar]
- Papadopoulos, E.; Hamhougias, K.; Himonas, C.; Dorchies, P. Strongyle anthelmintic resistance in horses and cattle from Greece. Rev. Med. Vet. 2000, 151, 1139–1142. [Google Scholar]
- Hellenic Statistics Authority. Statistics of Horticulture, Agriculture and Fisheries. Statistics of Animals. Statistics of Sheep—Farms with Sheep 2017. Available online: www.statistics.grel/statistics/agr (accessed on 20 April 2020).
- ELGO Demeter. Deliveries of Sheep and Goat Milk by Region and Local Authority for Calendar Year 2019; ELGO Demeter: Athens, Greece, 2020. [Google Scholar]
- Lianou, L.T.; Michael, C.K.; Vasileiou, N.G.C.; Chatzopoulos, D.C.; Mavrogianni, V.S.; Katsafadou, A.I.; Fthenakis, G.C. Association of vaccination with a biofilm-embedded bacteria-based vaccine against staphylococcal mastitis in sheep with lower somatic cell counts in bulk milk and lower frequency of biofilm-forming staphylococci in milking-machine clusters. In Proceedings of the Abstracts of Presentations of Conference “Next Generation Dairying for Scotland”, Edinburgh, UK, 25–26 November 2019. [Google Scholar]
- Umair, S.; Dagnicourt, E.; Knight, J.S.; Simpson, H.V.; Pernthaner, A. Molecular and biochemical characterisation of abomasal nematode parasites Teladorsagia circumcincta and Haemonchus contortus phosphofructokinases and their recognition by the immune host. Exp. Parasitol. 2015, 151, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Hoglund, J.; Gustafsson, K.; Ljungstrom, B.L.; Engstrom, A.; Donnan, A.; Skuce, P. Anthelmintic resistance in Swedish sheep flocks based on a comparison of the results from the faecal egg count reduction test and resistant allele frequencies of the betatubulin gene. Vet. Parasitol. 2009, 161, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Kotze, A.C.; Cowling, K.; Bagnall, N.H.; Hines, B.M.; Ruffell, A.P.; Hunt, P.W.; Coleman, G.T. Relative level of thiabendazole resistance associated with the E198A and F200Y SNPs in larvae of a multi-drug resistant isolate of Haemonchus contortus. Int. J. Parasitol. Drugs Drug. Resist. 2012, 2, 92–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redman, E.; Whitelaw, F.; Tait, A.; Burgess, C.; Bartley, Y.; Skuce, P.J.; Jackson, F.; Gilleard, J.S. The emergence of resistance to the benzimidazole anthlemintics in parasitic nematodes of livestock is characterised by multiple independent hard and soft selective sweeps. PLoS Negl. Trop. Dis. 2015, 6, e0003494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrère, V.; Álvarez, L.; Suárez, G.; Ceballos, L.; Moreno, L.; Lanusse, C.; Prichard, R.K. Relationship between increased albendazole systemic exposure and changes in single nucleotide polymorphisms on the β-tubulin isotype 1 encoding gene in Haemonchus contortus. Vet. Parasitol. 2012, 186, 344–349. [Google Scholar] [CrossRef]
- Barrère, V.; Falzon, L.C.; Shakya, K.P.; Menzies, P.I.; Peregrine, A.S.; Prichard, R.K. Assessment of benzimidazole resistance in Haemonchus contortus in sheep flocks in Ontario, Canada: Comparison of detection methods for drug resistance. Vet. Parasitol. 2013, 198, 159–165. [Google Scholar] [CrossRef]
- Barrère, V.; Keller, K.; von Samson-Himmelstjerna, G.; Prichard, R.K. Efficiency of a genetic test to detect benzimidazole resistant Haemonchus contortus nematodes in sheep farms in Quebec. Can. Parasitol. Int. 2013, 62, 464–470. [Google Scholar] [CrossRef]
- Rufener, L.; Kaminsky, R.; Maser, P. In vitro selection of Haemonchus contortus for benzimidazole resistance reveals a mutation at amino acid 198 of beta-tubulin. Mol. Biochem. Parasitol. 2009, 168, 120–122. [Google Scholar] [CrossRef]
- Mottier, M.L.; Prichard, R.K. Genetic analysis of a relationship between macrocyclic lactone and benzimidazole anthelmintic selection on Haemonchus contortus. Pharmacogenet. Genom. 2008, 18, 129–140. [Google Scholar] [CrossRef]
- Hillis, D.; Mable, B.K.; Larson, A.; Davis, S.K.; Zimmer, E.A. Nucleic acids IV: Sequencing and cloning. In Molecular Systematics, 2nd ed.; Hills, D., Moritz, C., Mable, B.K., Eds.; Sinauer Associates: Sunderland, MA, USA, 1996; pp. 321–382. [Google Scholar]
- Humbert, J.H.; Elard, L. A simple PCR method for rapidly detecting defined point mutations. Tech. Tips Online 1997, 2, 48–49. [Google Scholar] [CrossRef] [Green Version]
- Gallidis, E. Evaluation and Efficacy of the Targeted Selective Treatments against Gastrointestinal Nematode Parasites of Small Ruminants. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2012. [Google Scholar]



{kind=link}
{kind=link}
| Species | Susceptibility/Resistance Status (%) | |||
|---|---|---|---|---|
| S/S | R/S | R/R | Total | |
| Sheep | 0 (0.0%) | 3 (3.1%) | 93 (96.9%) | 96 (100.0%) |
| Goats | 0 (0.0%) | 0 (0.0%) | 96 (100.0%) | 96 (100.0% |
| Cattle | 0 (0.0%) | 36 (75.0%) | 12 (25.0%) | 48 (100.0%) |
| Buffaloes | 0 (0.0%) | 44 (91.7%) | 4 (8.3%) | 48 (100.0%) |
| Total | 0 (0.0%) | 83 (28.8%) | 205 (71.2%) | 288 (100.0%) |
| Primer | Sequence | Quantity in Reaction Mixture | |
| P1 | Fw: 5′-GTCCCACGTGCTGTTCTTGT -3′ | 0.4 μL | |
| P2S | Rv: 5′-TACAGAGCTTCATTATCGATGCAGA -3′ | 1.0 μL | |
| P3R | Fw: 5′-TTGGTAGAAAACACCGATGAAACATA-3′ | 1.0 μL | |
| P4 | Rv: 5′-GATCAGCATTCAGCTGTCCA-3′ | 0.4 μL | |
| Working Conditions | |||
| Target | Primer Pair | Product Size (bp) | Cycling Protocol |
| Presence of parasite | P1/P4 | 827 | Denaturation: 94 °C for 30 s; Annealing: 57 °C for 30 s; Extension: at 72 °C for 45 s |
| Susceptible alleles | P1/P2S | 635 | |
| Resistant alleles | P3R/P4 | 242 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arsenopoulos, K.; Minoudi, S.; Symeonidou, I.; Triantafyllidis, A.; Katsafadou, A.I.; Lianou, D.T.; Fthenakis, G.C.; Papadopoulos, E. Frequency of Resistance to Benzimidazoles of Haemonchus contortus Helminths from Dairy Sheep, Goats, Cattle and Buffaloes in Greece. Pathogens 2020, 9, 347. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050347
Arsenopoulos K, Minoudi S, Symeonidou I, Triantafyllidis A, Katsafadou AI, Lianou DT, Fthenakis GC, Papadopoulos E. Frequency of Resistance to Benzimidazoles of Haemonchus contortus Helminths from Dairy Sheep, Goats, Cattle and Buffaloes in Greece. Pathogens. 2020; 9(5):347. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050347
Chicago/Turabian StyleArsenopoulos, Konstantinos, Styliani Minoudi, Isaia Symeonidou, Alexandros Triantafyllidis, Angeliki I. Katsafadou, Daphne T. Lianou, George C. Fthenakis, and Elias Papadopoulos. 2020. "Frequency of Resistance to Benzimidazoles of Haemonchus contortus Helminths from Dairy Sheep, Goats, Cattle and Buffaloes in Greece" Pathogens 9, no. 5: 347. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9050347