In Silico Design and Validation of OvMANE1, a Chimeric Antigen for Human Onchocerciasis Diagnosis

 , , , , , ,
, , , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Peptide Selection and Stage-Specific Expression
2.2. Antigenicity, Physicochemical Properties and Solubility of OvMANE1 Chimeric Antigen
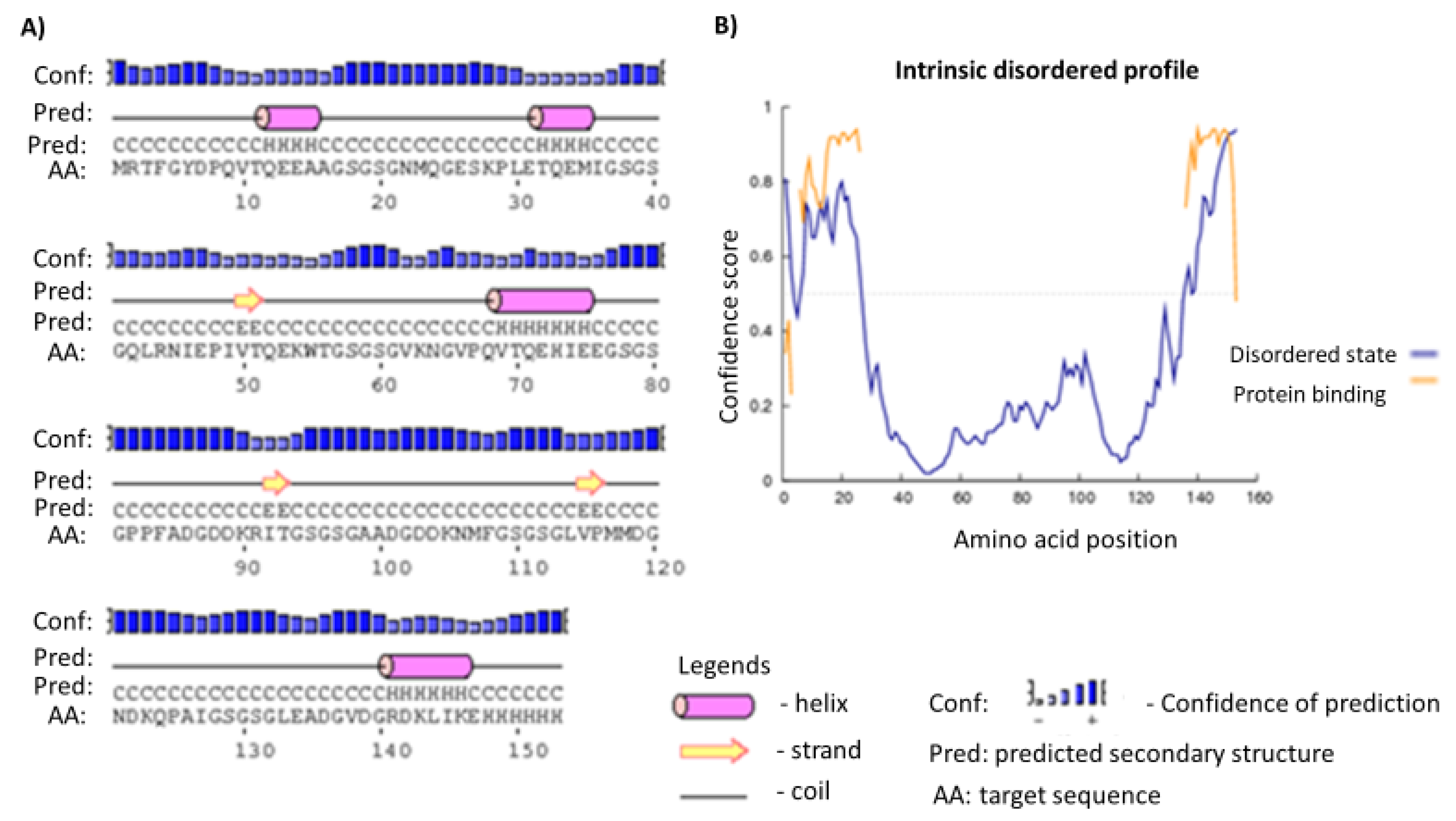
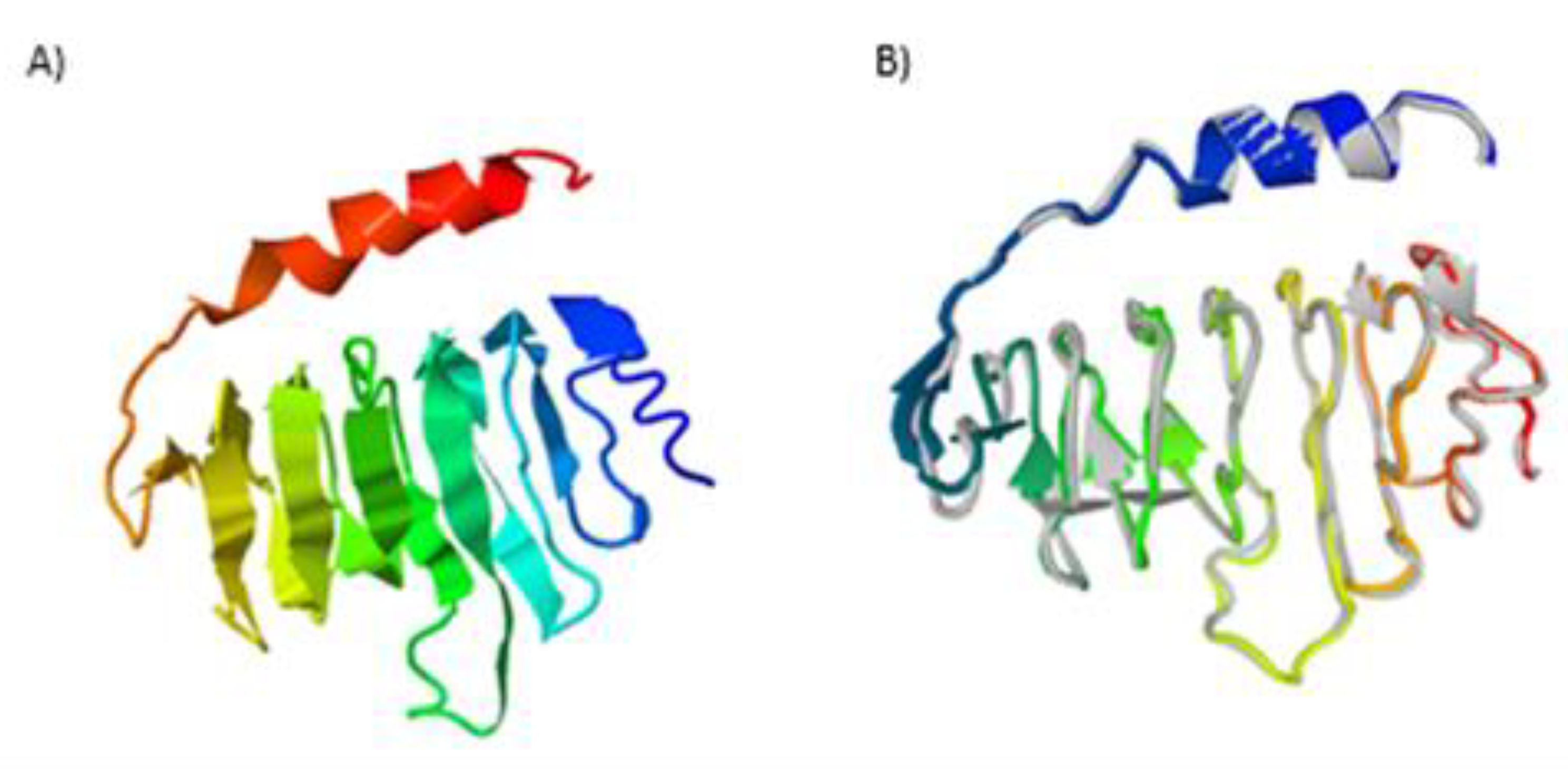
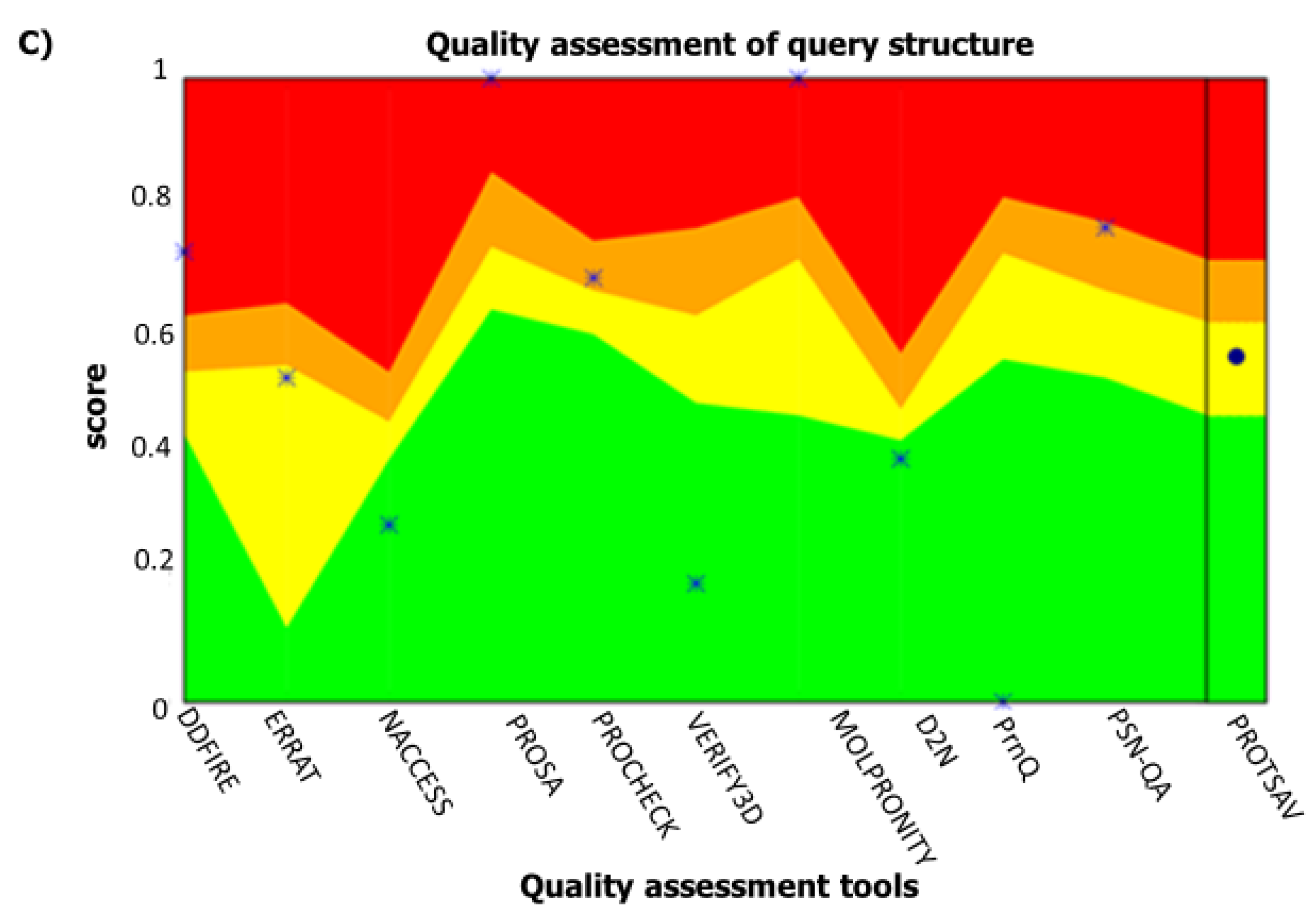
2.3. Secondary and Tertiary Structures
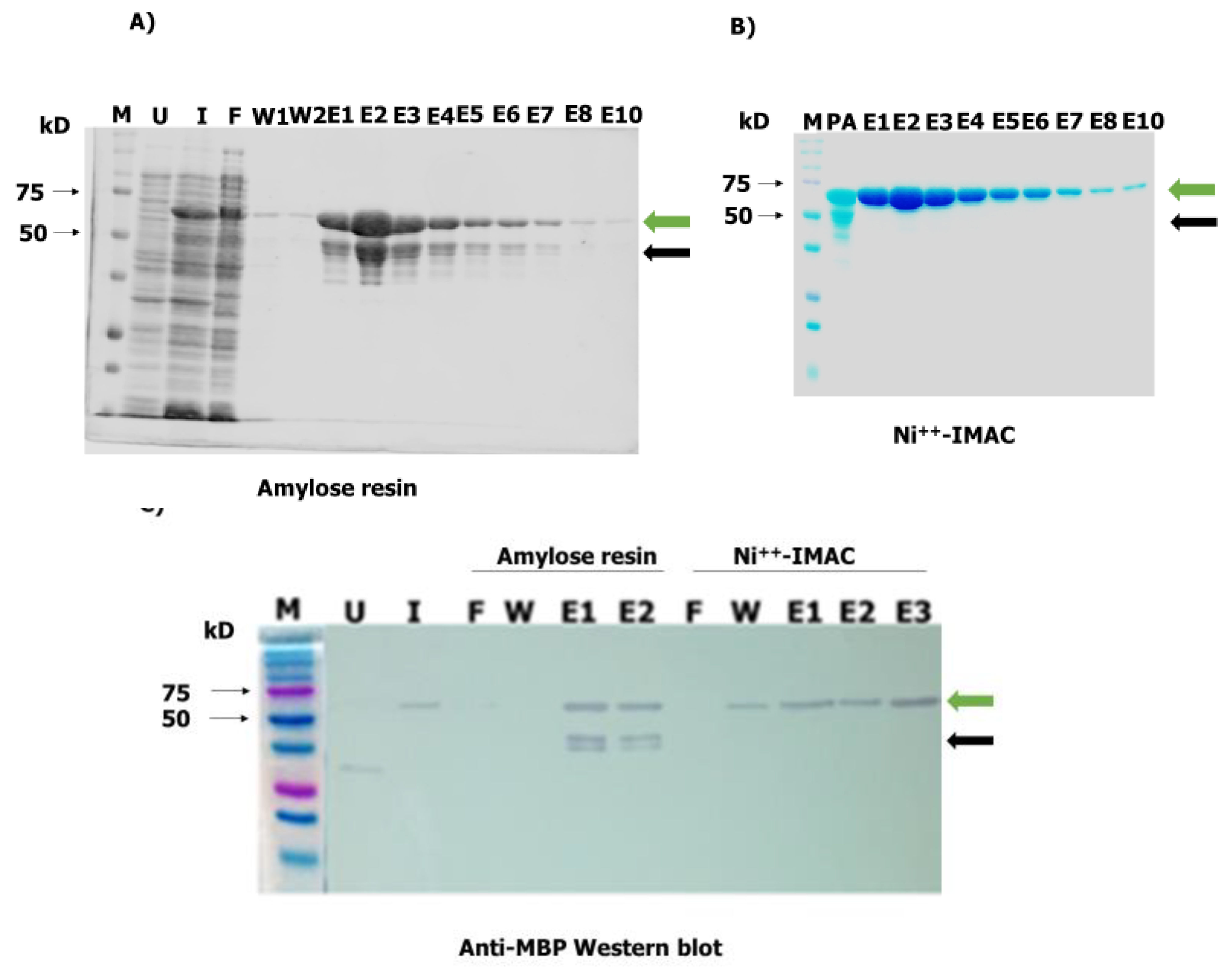
2.4. Mass Expression and Purification of Recombinant OvMANE1 Chimeric Antigen from Bacterial Cells
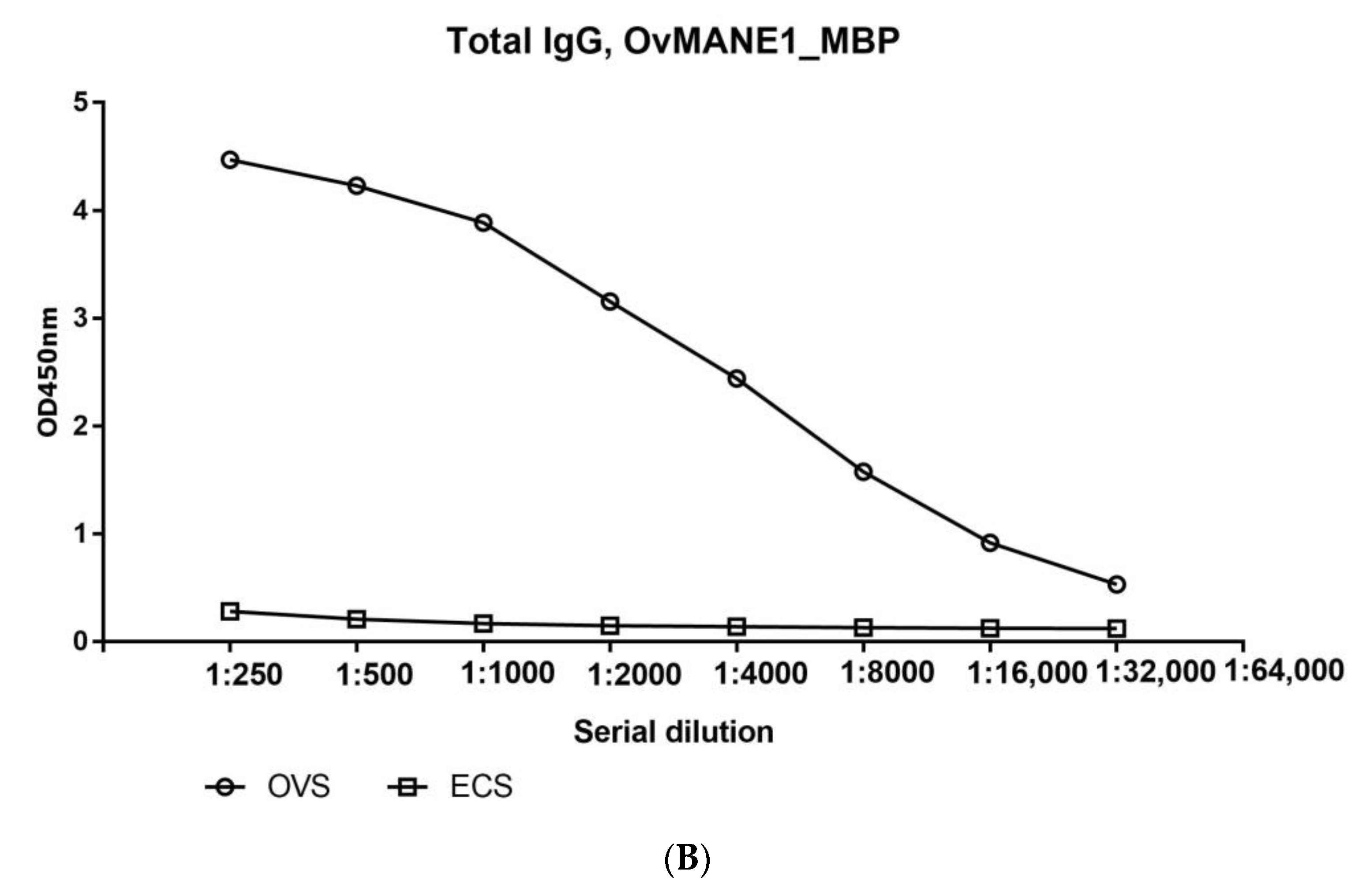
2.5. The Humoral Response to OvMANE1 Chimeric Antigen Divulges Its Diagnostic Potentials
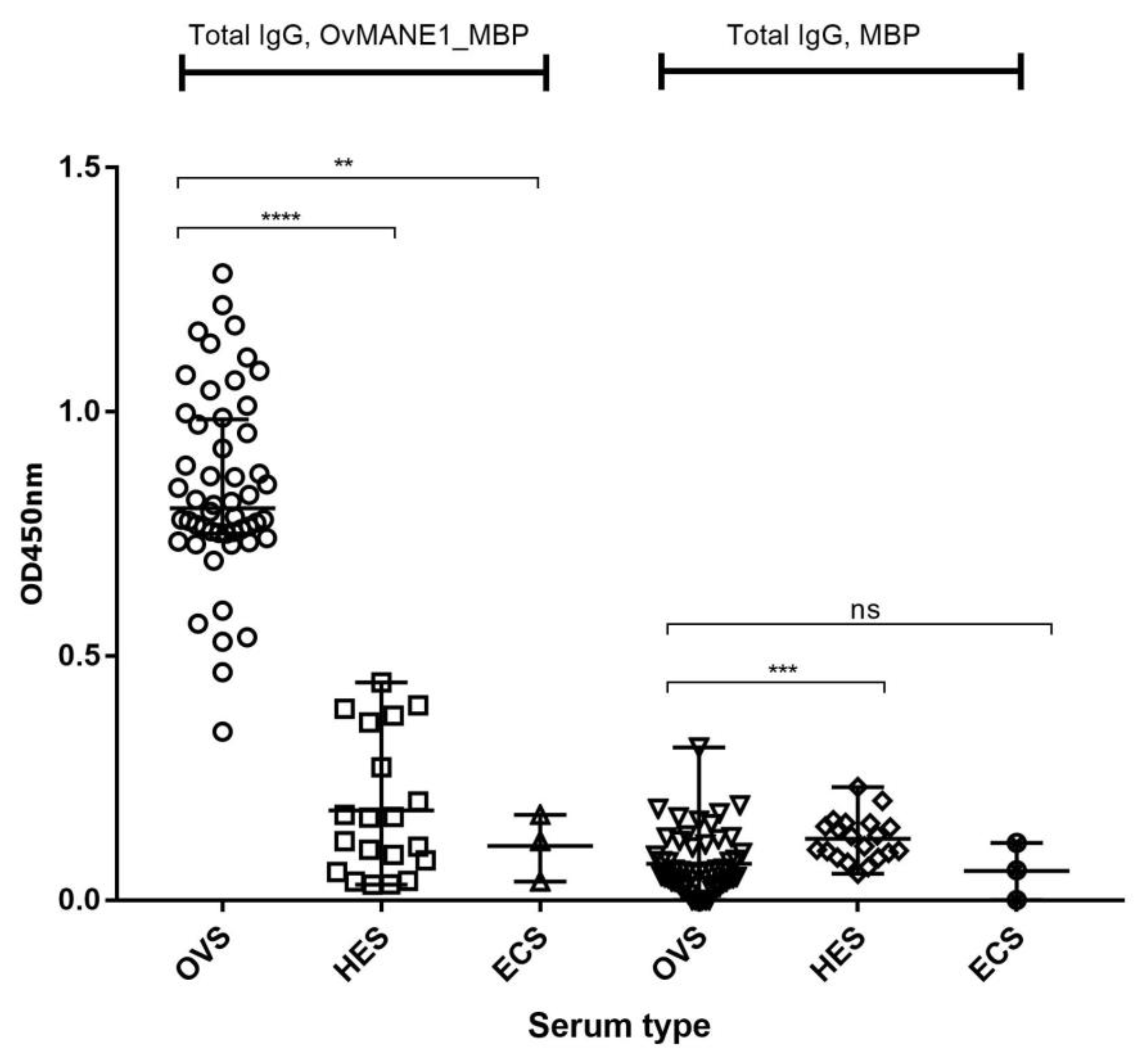
2.6. Analysis of Total IgG Responses to OvMANE1 Chimeric Antigen in Communities of Different Levels of Endemicity
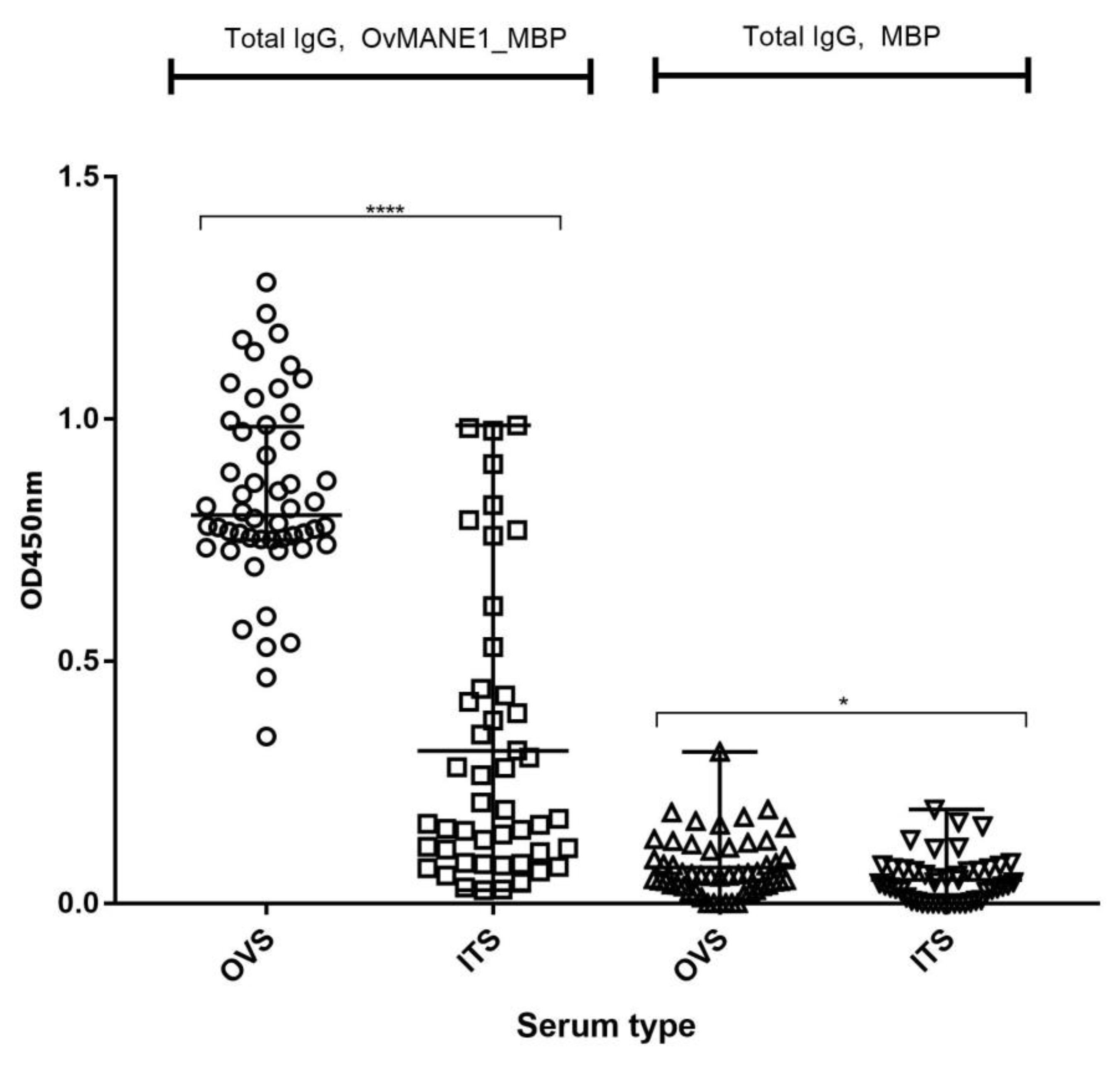
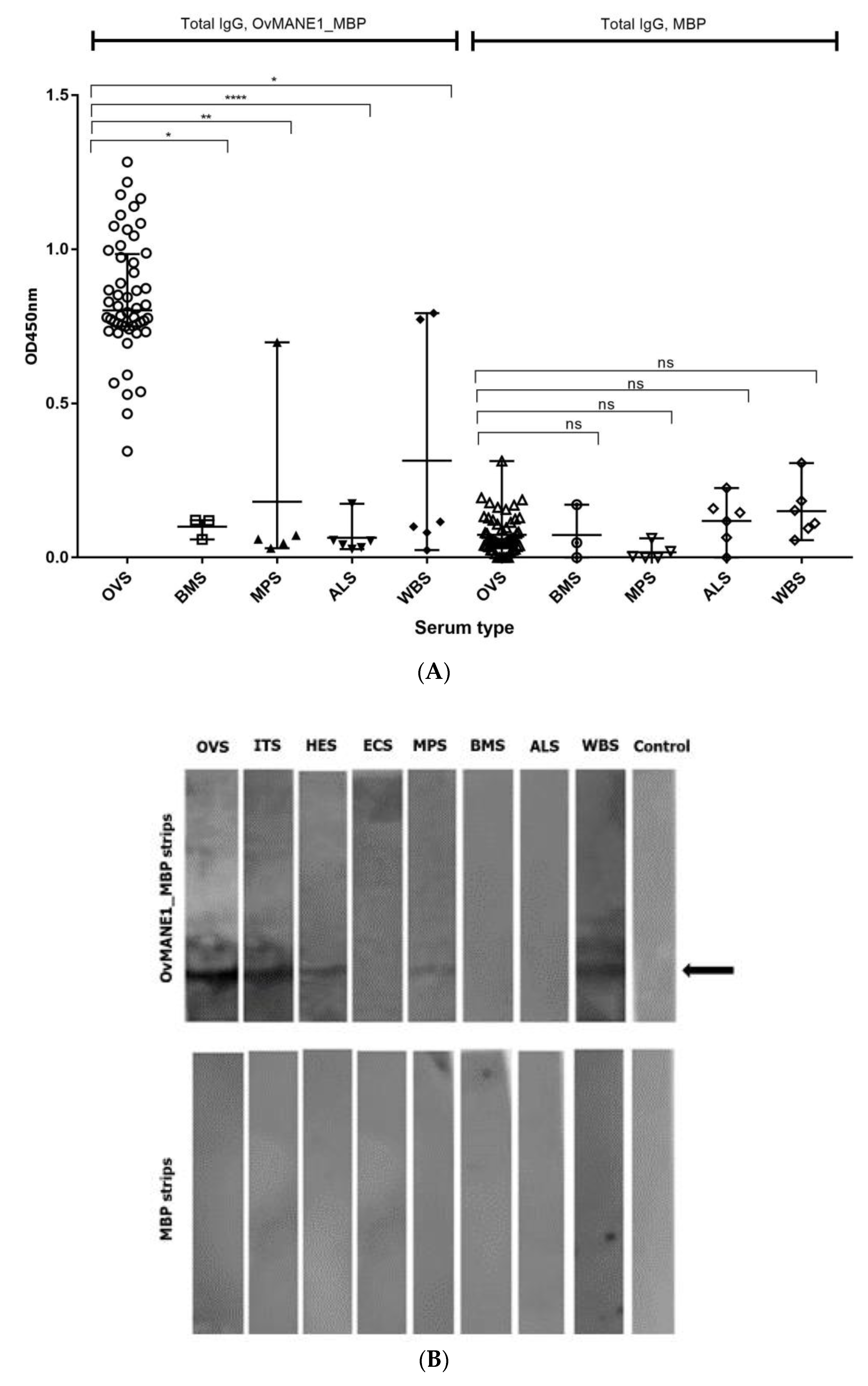
2.7. Cross-Reactivity Test with Related Nematode
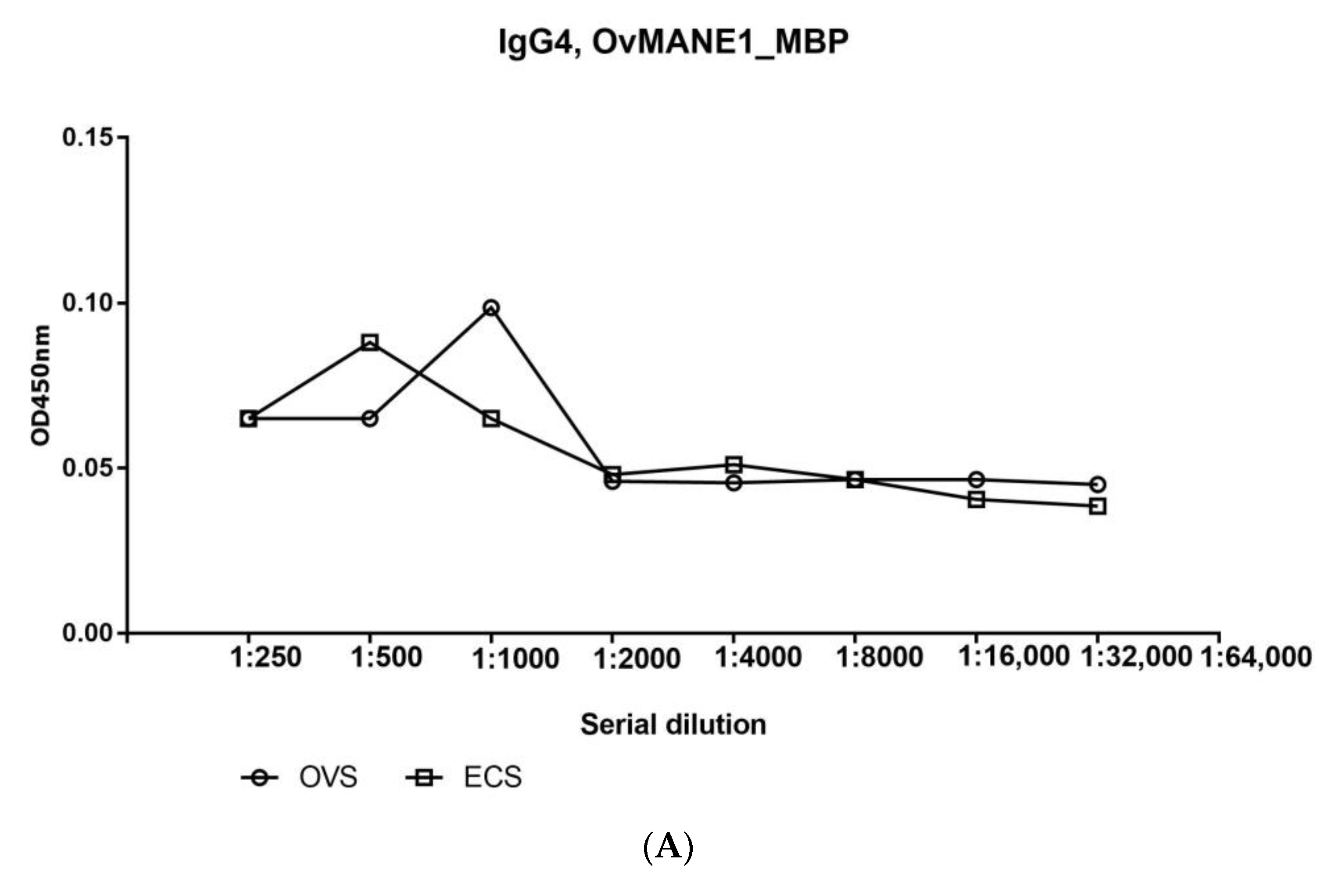
2.8. Total IgG but Not the IgG4 Subclass Responded Positively to OvMANE1 Chimeric Antigen
3. Discussion
4. Materials and Methods
4.1. Ethical Considerations
4.2. Serum Samples
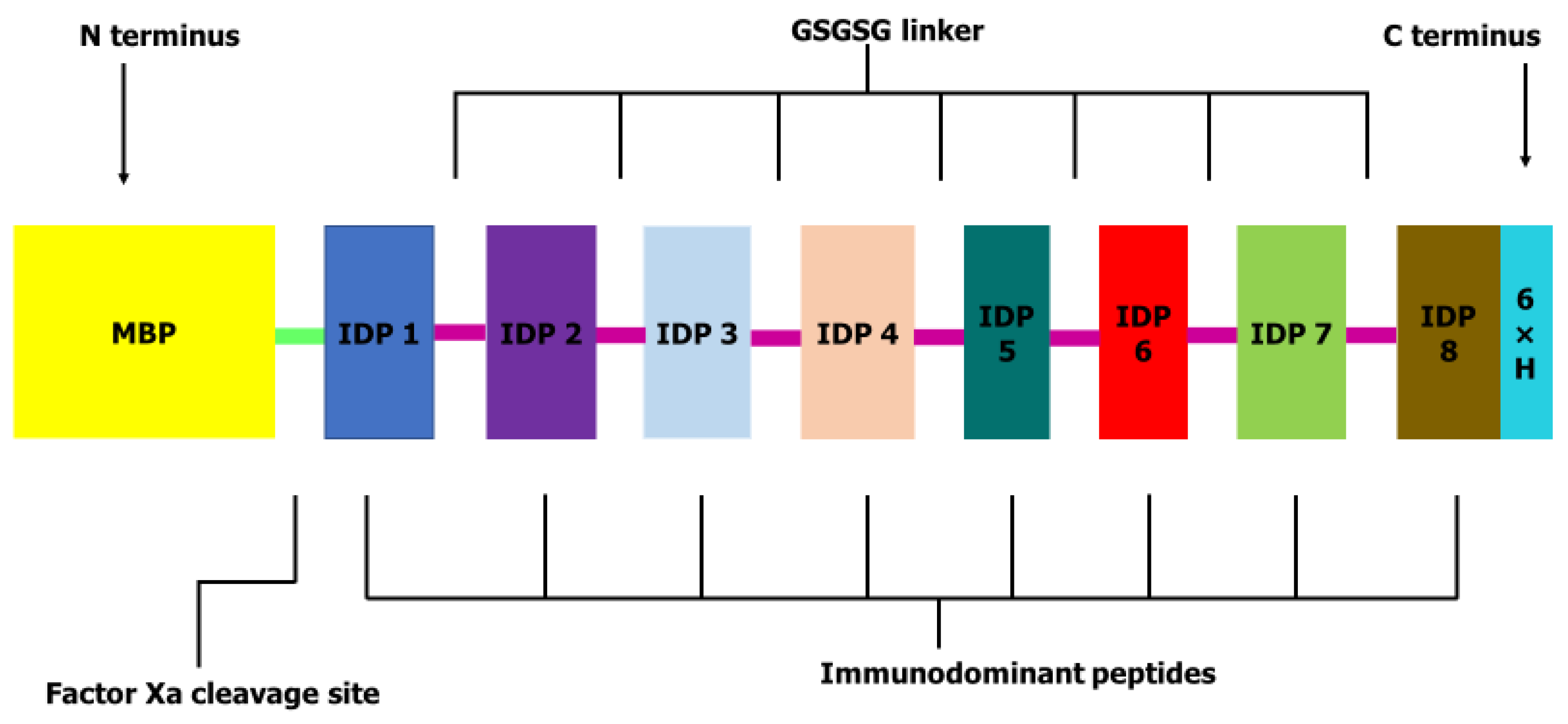
4.3. Peptide Selection, Stage-Specific Expression and Construction of OvMANE1 Chimeric Antigen
4.4. Prediction of Antigenicity, Physicochemical Properties and Solubility of OvMANE1 Chimeric Antigen
4.5. Prediction of Secondary and Tertiary Structures
4.6. Codon Optimization, Cloning, Expression and Purification of OvMANE1 Chimeric Antigen
4.7. Serological Characterization of OvMANE1 Chimeric Antigen
4.8. Data Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shey, R.A.; Ghogomu, S.M.; Esoh, K.K.; Nebangwa, N.D.; Shintouo, C.M.; Nongley, N.F.; Asa, B.F.; Ngale, F.N.; Vanhamme, L.; Souopgui, J. In-silico design of a multi-epitope vaccine candidate against onchocerciasis and related filarial diseases. Sci. Rep. 2019, 9, 4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organisation. Onchocerciasis Key Facts. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/onchocerciasis (accessed on 23 March 2020).
- Etya’alé, D. Eliminating onchocerciasis as a public health problem: The beginning of the end. Br. J. Ophthal. 2002, 86, 844–846. [Google Scholar] [CrossRef] [Green Version]
- Colebunders, R.; Siewe, F.N.; Hotterbeekx, A. Onchocerciasis-Associated Epilepsy, an Additional Reason for Strengthening Onchocerciasis Elimination Programs. Trends Parasitol. 2018, 34, 208–216. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, C.D.; Homeida, M.M.; Hopkins, A.D.; Lawrence, J.C. Elimination of onchocerciasis from Africa: Possible? Trends Parasitol. 2012, 28, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Loum, D.; Katholi, C.R.; Lakwo, T.; Habomugisha, P.; Tukahebwa, E.M.; Unnasch, T.R. Evaluation of Community-Directed Operation of Black Fly Traps for Entomological Surveillance of Onchocerca volvulus Transmission in the Madi-Mid North Focus of Onchocerciasis in Northern Uganda. Am. J. Trop. Med. Hyg. 2017, 97, 1235–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuesel, A.C. Research for new drugs for elimination of onchocerciasis in Africa. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 272–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantey, P.T.; Roy, S.L.; Boakye, D.; Mwingira, U.; Ottesen, E.A.; Hopkins, A.D.; Sodahlon, Y.K. Transitioning from river blindness control to elimination: Steps toward stopping treatment. Int. Health 2018, 10, i7–i13. [Google Scholar] [CrossRef] [Green Version]
- Vlaminck, J.; Fischer, P.U.; Weil, G.J. Diagnostic Tools for Onchocerciasis Elimination Programs. Trends Parasitol. 2015, 31, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Bottomley, C.; Isham, V.; Vivas-Martínez, S.; Kuesel, A.C.; Attah, S.K.; Opoku, N.O.; Lustigman, S.; Walker, M.; Basáñez, M.-G. Modelling Neglected Tropical Diseases diagnostics: The sensitivity of skin snips for Onchocerca volvulus in near elimination and surveillance settings. Parasites Vectors 2016, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.A.; Lustigman, S.; Zhang, S.; Weil, G.J. A comparison of newer tests for the diagnosis of onchocerciasis. Ann. Trop. Med. Parasitol. 2000, 94, 253–258. [Google Scholar] [CrossRef]
- World Health Organisation. Onchocerciasis, Guidelines for Stopping Mass Drug Administration and Verifying Elimination of Human Onchocerciasis; World Health Organisation: Geneva, Switzerland, 2016. [Google Scholar]
- Bennuru, S.; Oduro-Boateng, G.; Osigwe, C.; Del Valle, P.; Golden, A.; Ogawa, G.M.; Cama, V.; Lustigman, S.; Nutman, T.B. Integrating multiple biomarkers to increase sensitivity for the detection of Onchocerca volvulus infection. J. Infect. Dis. 2019, 221, 1805–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipner, E.M.; Dembele, N.; Souleymane, S.; Alley, W.S.; Prevots, D.R.; Toe, L.; Boatin, B.; Weil, G.J.; Nutman, T.B. Field applicability of a rapid-format anti-Ov-16 antibody test for the assessment of onchocerciasis control measures in regions of endemicity. J. Infect. Dis. 2006, 194, 216–221. [Google Scholar] [CrossRef]
- Gass, K.M. Rethinking the serological threshold for onchocerciasis elimination. PLoS Negl. Trop. Dis. 2018, 12, e0006249. [Google Scholar] [CrossRef]
- Noya, O.; Patarroyo, M.-E.; Guzman, F.; De Noya, B. Immunodiagnosis of parasitic diseases with synthetic peptides. Curr. Prot. Pept. Sci. 2003, 4, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.; Pozo, L.; Gómez, I.; Melchor, A. Chimeric synthetic peptide as antigen for immunodiagnosis of HIV-1 infection. Biochem. Biophys. Res. Commun. 2000, 272, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Tourdot, S.; Oukka, M.; Manuguerra, J.C.; Magafa, V.; Vergnon, I.; Riché, N.; Bruley-Rosset, M.; Cordopatis, P.; Kosmatopoulos, K. Chimeric peptides: A new approach to enhancing the immunogenicity of peptides with low MHC class I affinity: Application in antiviral vaccination. J. Immunol. 1997, 159, 2391–2398. [Google Scholar] [PubMed]
- Marin, M.H.; Peña, L.P.; Tanty, C.R.; Clarke, D.H.; Arenas, M.A.; Noguerol, K.R.; León, C.S. Antigenic activity of three chimeric synthetic peptides of the transmembrane (gp41) and the envelope (gp120) glycoproteins of HIV-1 virus. Prep. Biochem. Biotechnol. 2004, 34, 227–237. [Google Scholar] [CrossRef]
- Lu, Y.; Li, Z.; Teng, H.; Xu, H.; Qi, S.; He, J.; Gu, D.; Chen, Q.; Ma, H. Chimeric peptide constructs comprising linear B-cell epitopes: Application to the serodiagnosis of infectious diseases. Sci. Rep. 2015, 5, 13364. [Google Scholar] [CrossRef] [Green Version]
- Hajissa, K.; Zakaria, R.; Suppian, R.; Mohamed, Z. An evaluation of a recombinant multiepitope based antigen for detection of Toxoplasma gondii specific antibodies. BMC Infect. Dis. 2017, 17, 807. [Google Scholar] [CrossRef]
- Santos, F.L.N.; Celedon, P.A.F.; Zanchin, N.I.T.; de Souza, W.V.; da Silva, E.D.; Foti, L.; Krieger, M.A.; de Miranda Gomes, Y. Accuracy of chimeric proteins in the serological diagnosis of chronic chagas disease—A Phase II study. PLoS Negl. Trop. Dis. 2017, 11, e0005433. [Google Scholar] [CrossRef]
- Hajissa, K.; Zakaria, R.; Suppian, R.; Mohamed, Z. Design and evaluation of a recombinant multi-epitope antigen for serodiagnosis of Toxoplasma gondii infection in humans. Parasites Vectors 2015, 8, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, M.O.; Sako, Y.; Nakao, M.; Yamasaki, H.; Nakaya, K.; Ito, A. Evaluation of purified Taenia solium glycoproteins and recombinant antigens in the serologic detection of human and swine cysticercosis. J. Infect. Dis. 2006, 194, 1783–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkouawa, A.; Sako, Y.; Itoh, S.; Kouojip-Mabou, A.; Nganou, C.N.; Saijo, Y.; Knapp, J.; Yamasaki, H.; Nakao, M.; Nakaya, K.; et al. Serological Studies of Neurologic Helminthic Infections in Rural Areas of Southwest Cameroon: Toxocariasis, Cysticercosis and Paragonimiasis. PLoS Negl. Trop. Dis. 2010, 4, e732. [Google Scholar] [CrossRef] [PubMed]
- Lagatie, O.; Van Dorst, B.; Stuyver, L.J. Identification of three immunodominant motifs with atypical isotype profile scattered over the Onchocerca volvulus proteome. PLoS Negl. Trop. Dis. 2017, 11, e0005330. [Google Scholar] [CrossRef] [Green Version]
- Farahmand, B.; Taheri, N.; Shokouhi, H.; Soleimanjahi, H.; Fotouhi, F. Chimeric protein consisting of 3M2e and HSP as a universal influenza vaccine candidate: From in silico analysis to preliminary evaluation. Virus Genes 2019, 55, 22–32. [Google Scholar] [CrossRef]
- Ikai, A. Thermostability and aliphatic index of globular proteins. J. Biochem. 1980, 88, 1895–1898. [Google Scholar]
- Ali, M.; Pandey, R.K.; Khatoon, N.; Narula, A.; Mishra, A.; Prajapati, V.K. Exploring dengue genome to construct a multi-epitope based subunit vaccine by utilizing immunoinformatics approach to battle against dengue infection. Sci. Rep. 2017, 7, 9232. [Google Scholar] [CrossRef]
- Noma, M.; Zouré, H.G.; Tekle, A.H.; Enyong, P.; Nwoke, B.E.B.; Remme, J.H.F. The geographic distribution of onchocerciasis in the 20 participating countries of the African Programme for Onchocerciasis Control: (1) priority areas for ivermectin treatment. Parasites Vectors 2014, 7, 325. [Google Scholar] [CrossRef] [Green Version]
- Njume, F.N.; Ghogomu, S.M.; Shey, R.A.; Gainkam, L.O.T.; Poelvoorde, P.; Humblet, P.; Kamgno, J.; Robert, A.; Mutesa, L.; Lelubre, C.; et al. Identification and characterization of the Onchocerca volvulus Excretory Secretory Product Ov28CRP, a putative GM2 activator protein. PLoS Negl. Trop. Dis. 2019, 13, e0007591. [Google Scholar] [CrossRef] [Green Version]
- Ottesen, E.A.; Skvaril, F.; Tripathy, S.P.; Poindexter, R.W.; Hussain, R. Prominence of IgG4 in the IgG antibody response to human filariasis. J. Immunol. 1985, 134, 2707–2712. [Google Scholar]
- Gebrezgabiher, G.; Mekonnen, Z.; Yewhalaw, D.; Hailu, A. Reaching the last mile: Main challenges relating to and recommendations to accelerate onchocerciasis elimination in Africa. Infect. Dis. Poverty 2019, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vordermeier, H.M.; Whelan, A.; Cockle, P.J.; Farrant, L.; Palmer, N.; Hewinson, R.G. Use of synthetic peptides derived from the antigens ESAT-6 and CFP-10 for differential diagnosis of bovine tuberculosis in cattle. Clin. Diagn. Lab. Immunol. 2001, 8, 571–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corradin, G.; Villard, V.; Kajava, A.V. Protein structure based strategies for antigen discovery and vaccine development against malaria and other pathogens. Endocr. Metab. Immune Disord. Drug Targ. 2007, 7, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Lieutaud, P.; Ferron, F.; Uversky, A.V.; Kurgan, L.; Uversky, V.N.; Longhi, S. How disordered is my protein and what is its disorder for? A guide through the “dark side” of the protein universe. Intrinsically Disord. Prot. 2016, 4, e1259708. [Google Scholar] [CrossRef] [Green Version]
- Lagatie, O.; Verheyen, A.; Nijs, E.; Van Dorst, B.; Debrah, L.B.; Debrah, A.; Supali, T.; Sartono, E.; Stuyver, L. Evaluation of the Diagnostic Performance of Onchocerca volvulus Linear Epitopes in a Peptide Enzyme-Linked Immunosorbent Assay. Am. J. Trop. Med. Hyg. 2018, 98, 779–785. [Google Scholar] [CrossRef]
- Shey, R.A.; Ghogomu, S.M.; Njume, F.N.; Gainkam, L.O.T.; Poelvoorde, P.; Mutesa, L.; Robert, A.; Humblet, P.; Munyampundu, J.-P.; Kamgno, J.; et al. Prediction and validation of the structural features of Ov58GPCR, an immunogenic determinant of Onchocerca volvulus. PLoS ONE 2018, 13, e0202915. [Google Scholar] [CrossRef]
- Chandrashekar, R.; Ogunrinade, A.F.; Weil, G.J. Use of recombinant Onchocerca volvulus antigens for diagnosis and surveillance of human onchocerciasis. Trop. Med. Int. Health 1996, 1, 575–580. [Google Scholar] [CrossRef]
- Mpagi, J.L.; Buttner, D.W.; Tischendorf, F.W.; Erttmann, K.D.; Brattig, N.W. Humoral responses to a secretory Onchocerca volvulus protein: Differences in the pattern of antibody isotypes to recombinant Ov20/OvS1 in generalized and hyperreactive onchocerciasis. Parasite Immunol. 2000, 22, 455–460. [Google Scholar] [CrossRef]
- Katabarwa, M.N.; Eyamba, A.; Nwane, P.; Enyong, P.; Kamgno, J.; Kuété, T.; Yaya, S.; Aboutou, R.; Mukenge, L.; Kafando, C.; et al. Fifteen years of annual mass treatment of onchocerciasis with ivermectin have not interrupted transmission in the west region of cameroon. J. Parasitol. Res. 2013, 2013. [Google Scholar] [CrossRef]
- Kamga, G.-R.; Dissak-Delon, F.N.; Djeunga, H.N.; Biholong, B.D.; Ghogomu, S.M.; Souopgui, J.; Kamgno, J.; Robert, A. Important progress towards elimination of onchocerciasis in the West Region of Cameroon. Parasites Vectors 2017, 10, 373. [Google Scholar] [CrossRef]
- Tuck, M.K.; Chan, D.W.; Chia, D.; Godwin, A.K.; Grizzle, W.E.; Krueger, K.E.; Rom, W.; Sanda, M.; Sorbara, L.; Stass, S.; et al. Standard operating procedures for serum and plasma collection: Early detection research network consensus statement standard operating procedure integration working group. J. Proteome Res. 2009, 8, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, A.R.; Veloso, L.D.C.; Coura-Vital, W.; Reis, A.B.; Damasceno, L.M.; Gazzinelli, R.T.; Andrade, H. Novel recombinant multiepitope proteins for the diagnosis of asymptomatic leishmania infantum-infected dogs. PLoS Negl. Trop. Dis. 2015, 9, e3429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foroutan, M.; Ghaffarifar, F.; Sharifi, Z.; Dalimi, A.; Pirestani, M. Bioinformatics analysis of ROP8 protein to improve vaccine design against Toxoplasma gondii. Infect. Genet. Evol. 2018, 62, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Aathmanathan, S.; Jothi, N.; Prajapati, V.K.; Krishnan, M. Investigation of immunogenic properties of Hemolin from silkworm, Bombyx mori as carrier protein: An immunoinformatic approach. Sci. Rep. 2018, 8, 6957. [Google Scholar] [CrossRef] [PubMed]
- Hebditch, M.; Carballo-Amador, M.A.; Charonis, S.; Curtis, R.; Warwicker, J. Protein-Sol: A web tool for predicting protein solubility from sequence. Bioinformatics 2017, 33, 3098–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Peng, J.; Ma, J.; Xu, J. Protein Secondary Structure Prediction Using Deep Convolutional Neural Fields. Sci. Rep. 2016, 6, 18962. [Google Scholar] [CrossRef] [Green Version]
- Sellers, B.D.; Zhu, K.; Zhao, S.; Friesner, R.A.; Jacobson, M.P. Toward better refinement of comparative models: Predicting loops in inexact environments. Proteins 2008, 72, 959–971. [Google Scholar] [CrossRef] [Green Version]
- Delong, E.R.; Delong, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Antigenicity | |
|---|---|---|
| ANTIGENpro | VaxiJen 2.0 | |
| OvMANE1 | 0.917040 | 0.8524 |
| OvMANE1_MBP | 0.909324 | 0.5816 |
| OVOC5897 | 0.899261 | 0.5319 |
| OVOC4989 | 0.798654 | 0.5678 |
| OVOC5528 | 0.759612 | 0.5324 |
| OVOC9141 | 0.596235 | 0.4489 |
| OVOC7266 | 0.898052 | 0.6243 |
| OVOC1743 | 0.243291 | 0.5040 |
| OVOC1920 | 0.764529 | 0.5875 |
| OVOC3954 | 0.953580 | 0.5964 |
| Total IgG | ||
|---|---|---|
| ROC Curve Analysis | ROC curve area (AUC) | 0.9952 |
| 95% CI of AUC | 0.9845 to 1.006 | |
| p-value (against AUC = 0.5) | <0.0001 | |
| Diagnostic Accuracy Parameter | Cut off value | 0.456 |
| Sensitivity (%) (95% CI) | 98.08 (89.74% to 99.95%) | |
| Specificity (%) (95% CI) | 100 (83.16% to 100%) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shintouo, C.M.; Shey, R.A.; Nebangwa, D.N.; K. Esoh, K.; Nongley, N.F.; Nguve, J.E.; Giron, P.; Mutesa, L.; Vanhamme, L.; Souopgui, J.; et al. In Silico Design and Validation of OvMANE1, a Chimeric Antigen for Human Onchocerciasis Diagnosis. Pathogens 2020, 9, 495. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060495
Shintouo CM, Shey RA, Nebangwa DN, K. Esoh K, Nongley NF, Nguve JE, Giron P, Mutesa L, Vanhamme L, Souopgui J, et al. In Silico Design and Validation of OvMANE1, a Chimeric Antigen for Human Onchocerciasis Diagnosis. Pathogens. 2020; 9(6):495. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060495
Chicago/Turabian StyleShintouo, Cabirou Mounchili, Robert Adamu Shey, Derrick Neba Nebangwa, Kevin K. Esoh, Nkemngo Francis Nongley, Joel Ebai Nguve, Philippe Giron, Léon Mutesa, Luc Vanhamme, Jacob Souopgui, and et al. 2020. "In Silico Design and Validation of OvMANE1, a Chimeric Antigen for Human Onchocerciasis Diagnosis" Pathogens 9, no. 6: 495. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060495