The Epstein-Barr Virus-Encoded EBNA1 Protein Activates the Bone Morphogenic Protein (BMP) Signalling Pathway to Promote Carcinoma Cell Migration



Abstract
:1. Introduction
2. Results
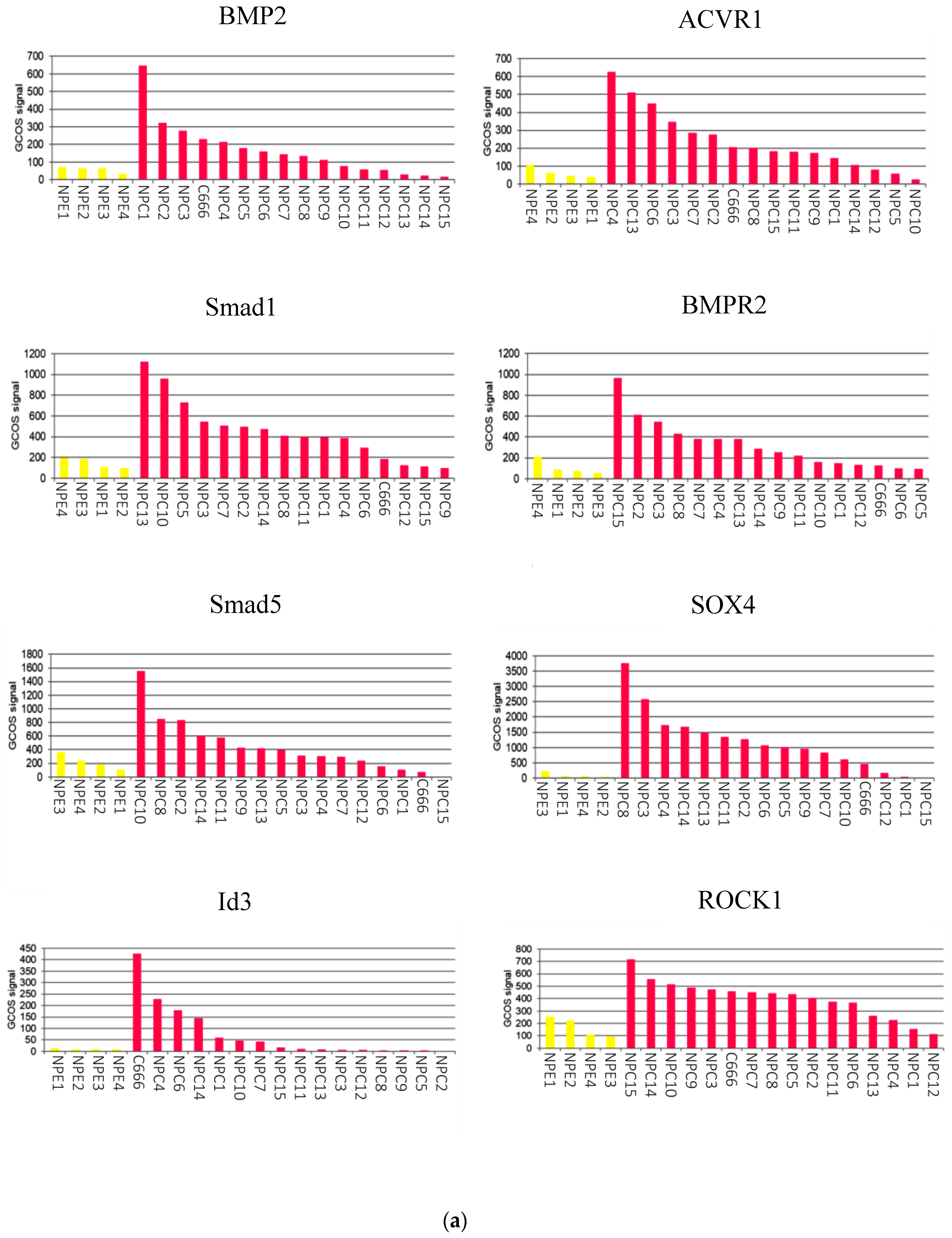
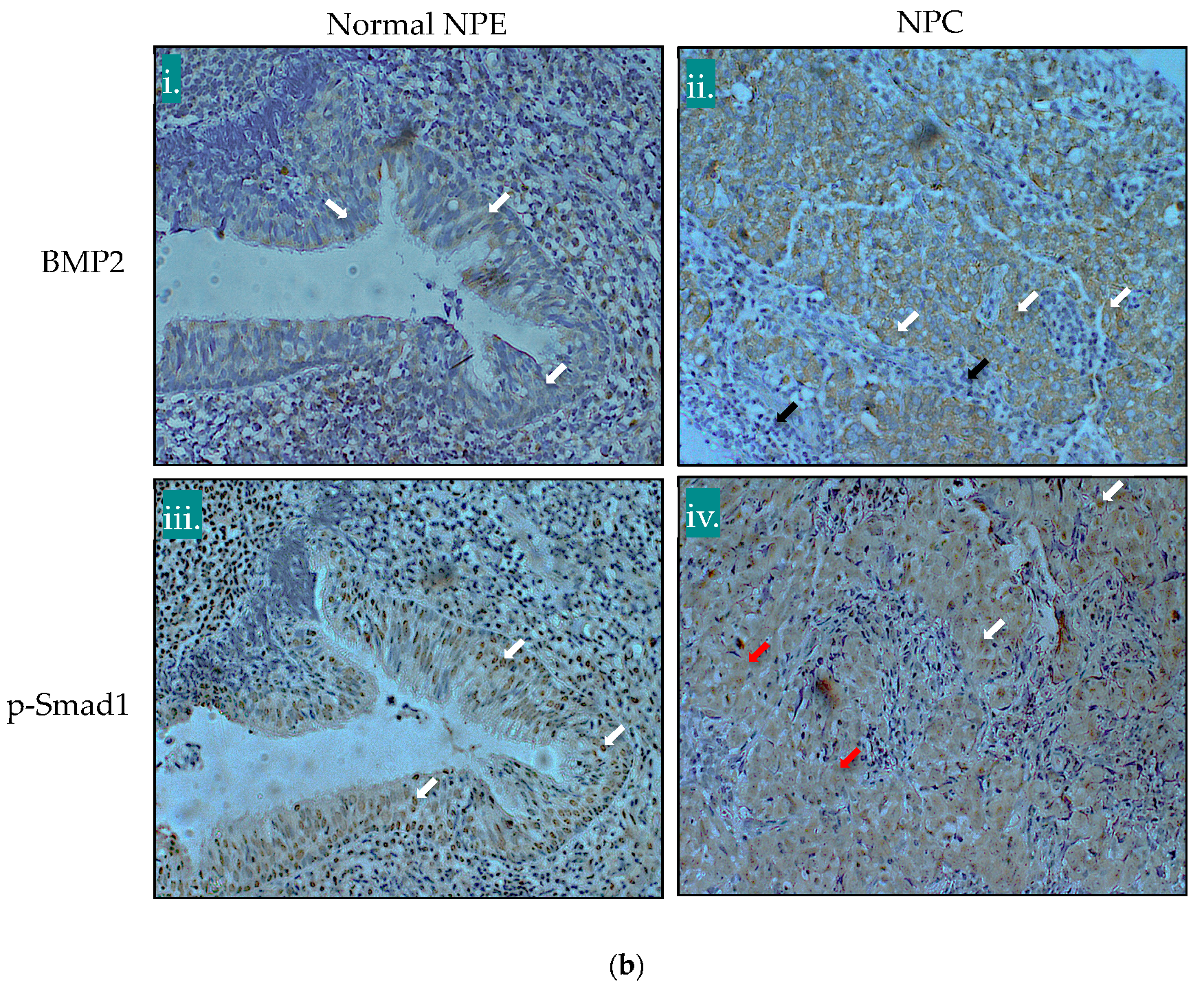
2.1. The BMP Signalling Pathway is Aberrantly Activated in NPC
2.2. The EBV-Positive NPC Cell Line, C666-1, Shows Constitutive BMP Pathway Activity
BRE-Luciferase Reporter Activity is Elevated in the EBV-Positive C666-1 Cell Line
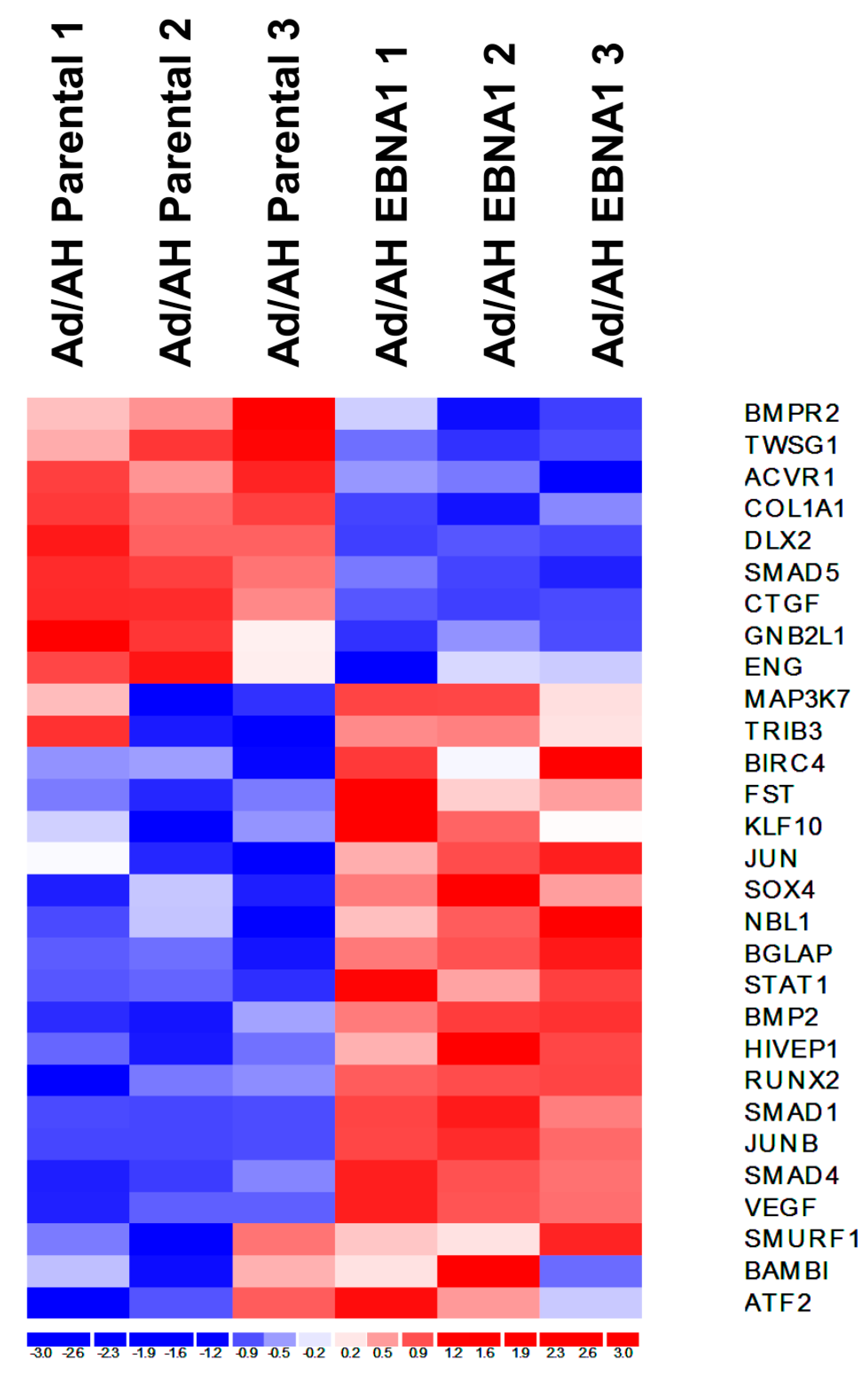
2.3. Gene Expression Profiling of BMP Pathway-Associated Genes in EBNA1-Expressing Ad/AH Cells
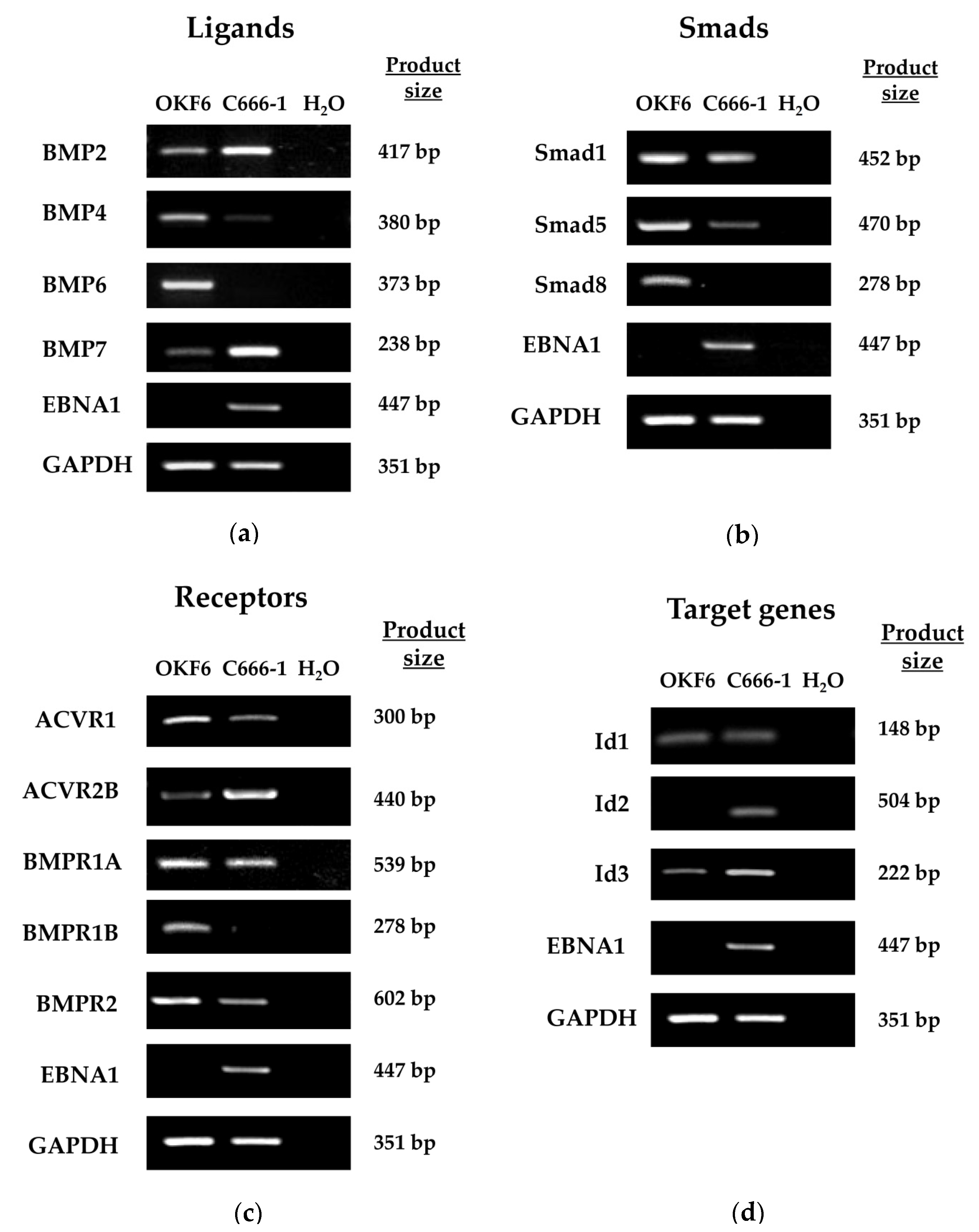
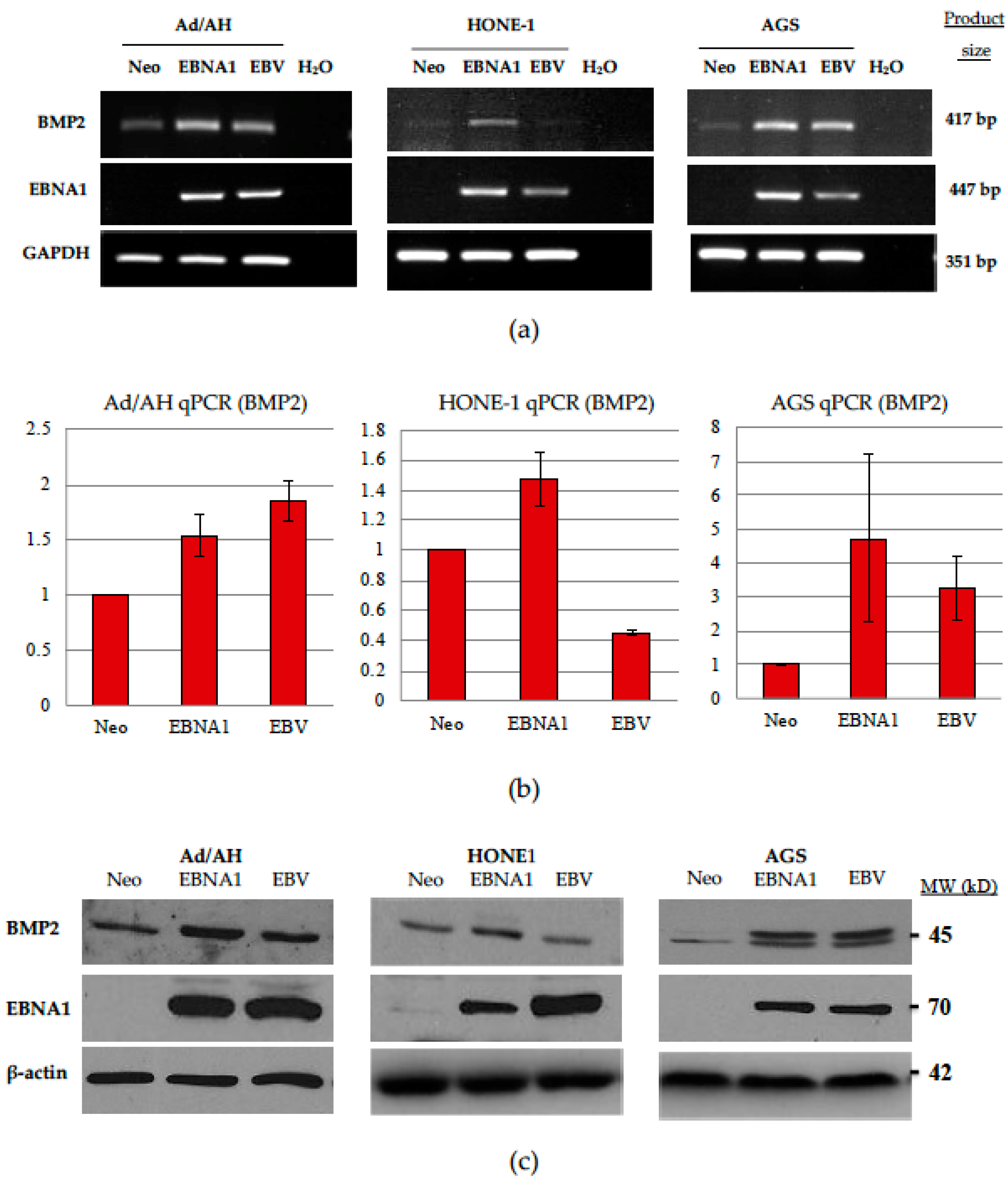
2.4. Expression of BMP2 Ligand Is Increased in Ad/AH, HONE-1 and AGS Cells Expressing EBNA1 or Latently Infected with EBV
2.5. Expression of BMP Pathway Components in Ad/AH, HONE-1 and AGS Cell Lines
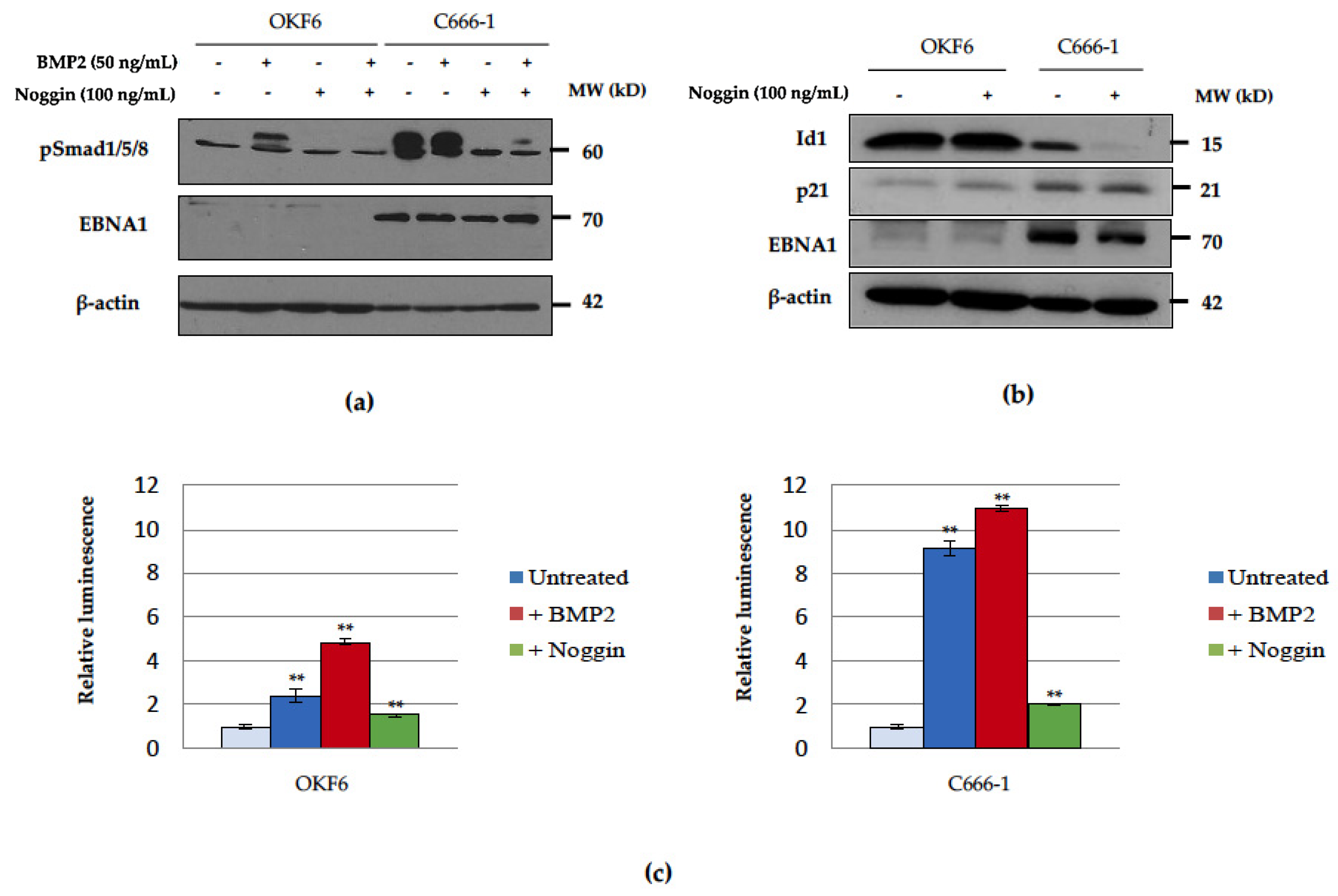
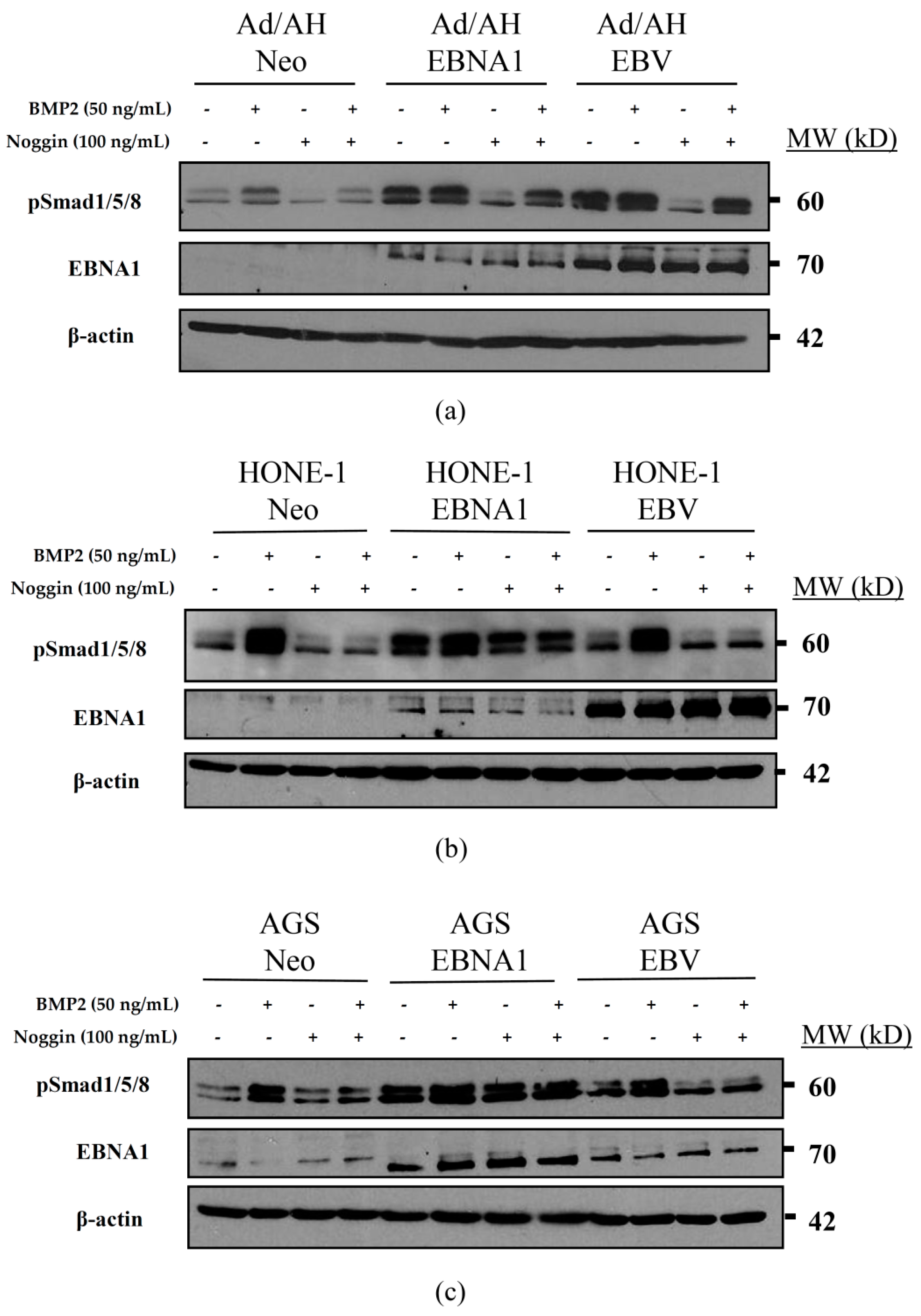
2.6. Basal Unstimulated Levels of Phosphorylated Smad1/5/8 Are Increased in the Presence of EBNA1 and EBV
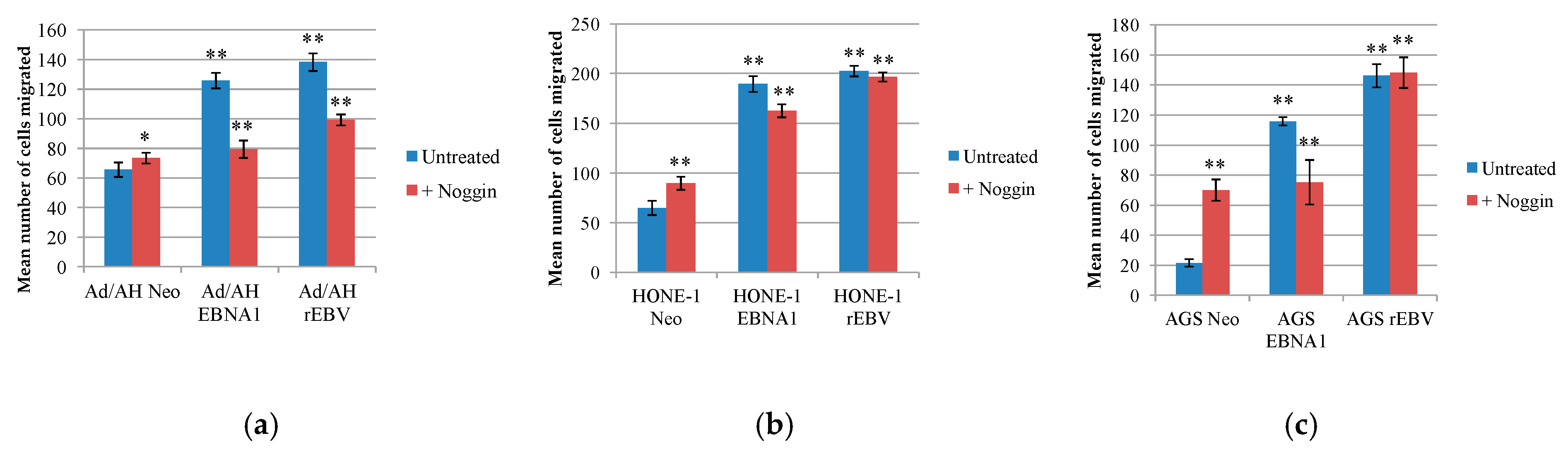
2.7. EBNA1 Induction of BMP2 Promotes Cell Migration, an Effect That Can Be Reduced by the Addition of the Natural BMP Antagonist, Noggin
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Tissue Culture
4.2. RT-PCR and Real-Time Quantitative PCR (qPCR)
4.3. GCOS Analysis of Gene Expression Data
4.4. Western Immunoblotting
4.5. Luciferase Reporter Assays
4.6. Transwell Migration Assays
4.7. Immunohistochemistry (IHC) and IHC Scoring
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kieff, E.D.; Rickinson, A.B. Epstein-Barr virus and its replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2603–2654. [Google Scholar]
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein–Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Thorley-Lawson, D.A.; Miyashita, E.M.; Khan, G. Epstein-Barr virus, and the B cell: That is all it takes. Trends Microbiol. 1996, 4, 204–208. [Google Scholar] [CrossRef]
- Miyashita, E.M.; Yang, B.; Babcock, G.J.; A Thorley-Lawson, D. Identification of the site of Epstein-Barr virus persistence in vivo as a resting B cell. J. Virol. 1997, 71, 4882–4891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, L.S.; Rickinson, A.B. Epstein–Barr virus: 40 years on. Nat. Rev. Cancer 2004, 4, 757–768. [Google Scholar] [CrossRef]
- Reisman, D.; Sugden, B. Trans activation of an Epstein-Barr viral transcriptional enhancer by the Epstein-Barr viral nuclear antigen 1. Mol. Cell. Boil. 1986, 6, 3838–3846. [Google Scholar] [CrossRef] [Green Version]
- Sugden, B.; Warren, N. A promoter of Epstein-Barr virus that can function during latent infection can be transactivated by EBNA-1, a viral protein required for viral DNA replication during latent infection. J. Virol. 1989, 63, 2644–2649. [Google Scholar] [CrossRef] [Green Version]
- Sample, J.; Henson, E.B.; Sample, C. The Epstein-Barr virus nuclear protein 1 promoter active in type I latency is autoregulated. J. Virol. 1992, 66, 4654–4661. [Google Scholar] [CrossRef] [Green Version]
- Gahn, T.A.; Sugden, B. An EBNA-1-dependent enhancer acts from 10 kilobase pairs to increase expression of the Epstein-Barr virus LMP gene. J. Virol. 1995, 69, 2633–2636. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.B.; Manet, E.; Gruffat, H.; Busson, P.; Blondel, M.; Fåhraeus, R. EBNA1: Oncogenic Activity, Immune Evasion and Biochemical Functions Provide Targets for Novel Therapeutic Strategies against Epstein-Barr Virus- Associated Cancers. Cancers 2018, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- Humme, S.; Reisbach, G.; Feederle, R.; Delecluse, H.-J.; Bousset, K.; Hammerschmidt, W.; Schepers, A. The EBV nuclear antigen 1 (EBNA1) enhances B cell immortalisation several thousandfold. Proc. Natl. Acad. Sci. USA 2003, 100, 10989–10994. [Google Scholar] [CrossRef] [Green Version]
- Komano, J.; Sugiura, M.; Takada, K. Epstein-Barr Virus Contributes to the Malignant Phenotype and to Apoptosis Resistance in Burkitt’s Lymphoma Cell Line Akata. J. Virol. 1998, 72, 9150–9156. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.B.; Bell, J.L.; Levine, A.J. Expression of Epstein-Barr virus nuclear antigen-1 induces B cell neoplasia in transgenic mice. EMBO J. 1996, 15, 3117–3126. [Google Scholar] [CrossRef]
- Kang, M.-S.; Lu, H.; Yasui, T.; Sharpe, A.; Warren, H.; Cahir-McFarland, E.; Bronson, R.; Hung, S.C.; Kieff, E. Epstein-Barr virus nuclear antigen 1 does not induce lymphoma in transgenic FVB mice. Proc. Natl. Acad. Sci. USA 2005, 102, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.-S.; Soni, V.; Bronson, R.; Kieff, E. Epstein-Barr Virus Nuclear Antigen 1 Does Not Cause Lymphoma in C57BL/6J Mice. J. Virol. 2008, 82, 4180–4183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, G.; Komano, J.; Sugden, B. Epstein-Barr virus provides a survival factor to Burkitt’s lymphomas. Proc. Natl. Acad. Sci. USA 2003, 100, 14269–14274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Murakami, M.; Verma, S.C.; Cai, Q.; Haldar, S.; Kaul, R.; Wasik, M.A.; Middeldorp, J.M.; Robertson, E.S. Epstein–Barr Virus nuclear antigen 1 (EBNA1) confers resistance to apoptosis in EBV-positive B-lymphoma cells through up-regulation of survivin. Virology 2011, 410, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holowaty, M.N.; Zeghouf, M.; Wu, H.; Tellam, J.; Athanasopoulos, V.; Greenblatt, J.; Frappier, L. Protein Profiling with Epstein-Barr Nuclear Antigen-1 Reveals an Interaction with the Herpesvirus-associated Ubiquitin-specific Protease HAUSP/USP7. J. Biol. Chem. 2003, 278, 29987–29994. [Google Scholar] [CrossRef] [Green Version]
- Holowaty, M.; Frappier, L. HAUSP/USP7 as an Epstein–Barr virus target. Biochem. Soc. Trans. 2004, 32, 731–732. [Google Scholar] [CrossRef] [PubMed]
- Sivachandran, N.; Sarkari, F.; Frappier, L. Epstein-Barr Nuclear Antigen 1 Contributes to Nasopharyngeal Carcinoma through Disruption of PML Nuclear Bodies. PLoS Pathog. 2008, 4, e1000170. [Google Scholar] [CrossRef] [Green Version]
- Sivachandran, N.; Cao, J.Y.; Frappier, L. Epstein-Barr Virus Nuclear Antigen 1 Hijacks the Host Kinase CK2 To Disrupt PML Nuclear Bodies. J. Virol. 2010, 84, 11113–11123. [Google Scholar] [CrossRef] [Green Version]
- Kube, D.; Vockerodt, M.; Weber, O.; Hell, K.; Wolf, J.; Haier, B.; Grässer, A.; Müller-Lantzsch, N.; Kieff, E.; Diehl, V.; et al. Expression of Epstein-Barr Virus Nuclear Antigen 1 Is Associated with Enhanced Expression of CD25 in the Hodgkin Cell Line L428. J. Virol. 1999, 73, 1630–1636. [Google Scholar] [CrossRef] [Green Version]
- Baumforth, K.R.; Birgersdotter, A.; Reynolds, G.M.; Wei, W.; Kapatai, G.; Flavell, J.R.; Kalk, E.; Piper, K.; Lee, S.P.; Machado, L.R.; et al. Expression of the Epstein-Barr Virus-Encoded Epstein-Barr Virus Nuclear Antigen 1 in Hodgkin’s Lymphoma Cells Mediates Up-Regulation of CCL20 and the Migration of Regulatory T Cells. Am. J. Pathol. 2008, 173, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Flavell, J.R.; Baumforth, K.R.N.; Wood, V.H.J.; Davies, G.L.; Wei, W.; Reynolds, G.M.; Morgan, S.; Boyce, A.; Kelly, G.L.; Young, L.S.; et al. Down-regulation of the TGF-beta target gene, PTPRK, by the Epstein-Barr virus–encoded EBNA1 contributes to the growth and survival of Hodgkin lymphoma cells. Blood 2008, 111, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, S.K.; Sixbey, J.W. Epstein-Barr virus induction of recombinase-activating genes RAG1 and RAG2. J. Virol. 1995, 69, 8155–8158. [Google Scholar] [CrossRef] [Green Version]
- Wood, V.H.J.; O’Neil, J.D.; Wei, W.; Stewart, S.E.; Dawson, C.W.; Young, L.S. Epstein–Barr virus-encoded EBNA1 regulates cellular gene transcription and modulates the STAT1 and TGFβ signaling pathways. Oncogene 2007, 26, 4135–4147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neil, J.D.; Owen, T.J.; Wood, V.H.J.; Date, K.L.; Valentine, R.; Chukwuma, M.B.; Arrand, J.R.; Dawson, C.W.; Young, L.S. Epstein–Barr virus-encoded EBNA1 modulates the AP-1 transcription factor pathway in nasopharyngeal carcinoma cells and enhances angiogenesis in vitro. J. Gen. Virol. 2008, 89, 2833–2842. [Google Scholar] [CrossRef]
- Valentine, R.; Dawson, C.W.; Hu, C.; Shah, K.M.; Owen, T.J.; Date, K.L.; Maia, S.P.; Shao, J.-Y.; Arrand, J.R.; Young, L.S.; et al. Epstein-Barr virus-encoded EBNA1 inhibits the canonical NF-κB pathway in carcinoma cells by inhibiting IKK phosphorylation. Mol. Cancer 2010, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, T.J.; O’Neil, J.D.; Dawson, C.W.; Hu, C.; Chen, X.; Yao, Y.; Wood, V.H.; Mitchell, L.; White, R.J.; Young, L.S.; et al. Epstein-Barr virus-encoded EBNA1 enhances RNA polymerase III-dependent EBER expression through induction of EBER-associated cellular transcription factors. Mol. Cancer 2010, 9, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katagiri, T.; Yamaguchi, A.; Komaki, M.; Abe, E.; Takahashi, N.; Ikeda, T.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic protein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Boil. 1994, 127, 1755–1766. [Google Scholar] [CrossRef] [Green Version]
- Ducy, P.; Karsenty, G. The family of bone morphogenetic proteins. Kidney Int. 2000, 57, 2207–2214. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Chen, D. The BMP signaling and in vivo bone formation. Gene 2005, 357, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, C.; Kopf, J.; Hiepen, C.; Knaus, P. Recent advances in BMP receptor signaling. Cytokine Growth Factor Rev. 2009, 20, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.L. Bone morphogenetic proteins: Multifunctional regulators of vertebrate development. Genes Dev. 1996, 10, 1580–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.; Mulder, K.M. Transforming growth factor-β signal transduction in epithelial cells. Pharmacol. Ther. 2001, 91, 1–34. [Google Scholar] [CrossRef]
- Miyazawa, K.; Shinozaki, M.; Hara, T.; Furuya, T.; Miyazono, K. Two major Smad pathways in TGF-β superfamily signalling. Genes Cells 2002, 7, 1191–1204. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L. BMP signaling and stem cell regulation. Dev. Boil. 2005, 284, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Alarmo, E.; Kallioniemi, A. Bone morphogenetic proteins in breast cancer: Dual role in tumourigenesis? Endocrine-Relat. Cancer 2010, 17, R123–R139. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Morris, R.J. The Yin and Yang of bone morphogenetic proteins in cancer. Cytokine Growth Factor Rev. 2010, 21, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Lorente-Trigos, A.; Varnat, F.; Melotti, A.; Altaba, A.R.I. BMP Signaling Promotes the Growth of Primary Human Colon Carcinomas in vivo. J. Mol. Cell Boil. 2010, 2, 318–332. [Google Scholar] [CrossRef] [Green Version]
- Langenfeld, E.M.; Kong, Y.; Langenfeld, J. Bone morphogenetic protein 2 stimulation of tumour growth involves the activation of Smad-1/5. Oncogene 2006, 25, 685–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markić, D.; Ćelić, T.; Španjol, J.; Gršković, A.; Bobinac, D.; Fučkar, Ž. Expression of bone morphogenetic protein 7, its receptors and Smad1/5/8 in normal human kidney and renal cell cancer. Coll. Antropol. 2010, 34, 149–153. [Google Scholar] [PubMed]
- Helms, M.W.; Packeisen, J.; August, C.; Schittek, B.; Boecker, W.; Brandt, B.H.; Buerger, H. First evidence supporting a potential role for the BMP/SMAD pathway in the progression of oestrogen receptor-positive breast cancer. J. Pathol. 2005, 206, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Katsuno, Y.; Hanyu, A.; Kanda, H.; Ishikawa, Y.; Akiyama, F.; Iwase, T.; Ogata, E.; Ehata, S.; Miyazono, K.; Imamura, T. Bone morphogenetic protein signaling enhances invasion and bone metastasis of breast cancer cells through Smad pathway. Oncogene 2008, 27, 6322–6333. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, S.; Gao, Y.-H.; Ohara-Nemoto, Y.; Kataoka, H.; Satoh, M. Expression of bone morphogenetic proteins of human neoplastic epithelial cells. IUBMB Life 1997, 42, 497–505. [Google Scholar] [CrossRef]
- Kleeff, J.; Maruyama, H.; Ishiwata, T.; Sawhney, H.; Friess, H.; Büchler, M.W.; Korc, M. Bone morphogenetic protein 2 exerts diverse effects on cell growth in vitro and is expressed in human pancreatic cancer in vivo. Gastroenterology 1999, 116, 1202–1216. [Google Scholar] [CrossRef]
- Jin, Y.; Tipoe, G.L.; Liong, E.C.; Lau, T.; Fung, P.; Leung, K.M. Overexpression of BMP-2/4, -5 and BMPR-IA associated with malignancy of oral epithelium. Oral Oncol. 2001, 37, 225–233. [Google Scholar] [CrossRef]
- Soares, A.F.; Xavier, R.L.D.F.; Miguel, M.C.D.C.; de Souza, L.B.; Pinto, L.P. Bone morphogenetic protein 2/4 and bone morphogenetic protein receptor type IA expression in metastatic and nonmetastatic oral squamous cell carcinoma. Am. J. Otolaryngol. 2010, 31, 266–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenfeld, E.M.; Bojnowski, J.; Perone, J.; Langenfeld, J. Expression of Bone Morphogenetic Proteins in Human Lung Carcinomas. Ann. Thorac. Surg. 2005, 80, 1028–1032. [Google Scholar] [CrossRef]
- Hu, C.; Wei, W.; Chen, X.; Woodman, C.B.; Yao, Y.; Nicholls, J.M.; Joab, I.; Sihota, S.K.; Shao, J.-Y.; Derkaoui, K.D.; et al. A Global View of the Oncogenic Landscape in Nasopharyngeal Carcinoma: An Integrated Analysis at the Genetic and Expression Levels. PLoS ONE 2012, 7, e41055. [Google Scholar] [CrossRef]
- Dickson, M.A.; Hahn, W.C.; Ino, Y.; Ronfard, V.; Wu, J.Y.; Weinberg, R.A.; Louis, D.N.; Li, F.P.; Rheinwald, J.G. Human Keratinocytes That Express hTERT and Also Bypass a p16INK4a-Enforced Mechanism That Limits Life Span Become Immortal yet Retain Normal Growth and Differentiation Characteristics. Mol. Cell. Biol. 2000, 20, 1436–1447. [Google Scholar] [CrossRef] [Green Version]
- Korchynskyi, O.; Dijke, P.T. Identification and Functional Characterization of Distinct Critically Important Bone Morphogenetic Protein-specific Response Elements in the Id1 Promoter. J. Biol. Chem. 2001, 277, 4883–4891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollnagel, A.; Oehlmann, V.; Heymer, J.; Rüther, U.; Nordheim, A. IdGenes Are Direct Targets of Bone Morphogenetic Protein Induction in Embryonic Stem Cells. J. Boil. Chem. 1999, 274, 19838–19845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Chen, C.-R.; Massagué, J. A Self-Enabling TGFβ Response Coupled to Stress Signaling. Mol. Cell 2003, 11, 915–926. [Google Scholar] [CrossRef]
- Kowanetz, M.; Valcourt, U.; Bergström, R.; Heldin, C.-H.; Moustakas, A. Id2 and Id3 Define the Potency of Cell Proliferation and Differentiation Responses to Transforming Growth Factor β and Bone Morphogenetic Protein. Mol. Cell. Boil. 2004, 24, 4241–4254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.-Z.; Miyake, S.; Akiyama, Y.; Yuasa, Y. BMP-2 modulates the proliferation and differentiation of normal and cancerous gastric cells. Biochem. Biophys. Res. Commun. 2004, 316, 100–106. [Google Scholar] [CrossRef]
- Feeley, B.T.; Krenek, L.; Liu, N.; Hsu, W.K.; Gamradt, S.C.; Schwarz, E.M.; Huard, J.; Lieberman, J.R. Overexpression of noggin inhibits BMP-mediated growth of osteolytic prostate cancer lesions. Bone 2006, 38, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Feeley, B.T.; Gamradt, S.C.; Hsu, W.K.; Liu, N.; Krenek, L.; Robbins, P.; Huard, J.; Lieberman, J.R. Influence of BMPs on the Formation of Osteoblastic Lesions in Metastatic Prostate Cancer. J. Bone Miner. Res. 2005, 20, 2189–2199. [Google Scholar] [CrossRef]
- Wu, J.-B.; Fu, H.-Q.; Huang, L.-Z.; Liu, A.-W.; Zhang, J.-X. Effects of siRNA-targeting BMP-2 on the abilities of migration and invasion of human liver cancer SMMC7721 cells and its mechanism. Cancer Gene Ther. 2010, 18, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Feeley, B.T.; Liu, N.Q.; Conduah, A.H.; Krenek, L.; Roth, K.; Dougall, W.C.; Huard, J.; Dubinett, S.; Lieberman, J.R. Mixed Metastatic Lung Cancer Lesions in Bone Are Inhibited by Noggin Overexpression and Rank:Fc Administration. J. Bone Miner. Res. 2006, 21, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Secondini, C.; Wetterwald, A.; Schwaninger, R.; Thalmann, G.N.; Cecchini, M.G. The Role of the BMP Signaling Antagonist Noggin in the Development of Prostate Cancer Osteolytic Bone Metastasis. PLoS ONE 2011, 6, e16078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrams, K.L.; Xu, J.; Nativelle-Serpentini, C.; Dabirshahsahebi, S.; Rogers, M.B. An Evolutionary and Molecular Analysis of Bmp2 Expression. J. Boil. Chem. 2004, 279, 15916–15928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Rogers, M.B. Modulation of Bone Morphogenetic Protein (BMP) 2 gene expression by Sp1 transcription factors. Gene 2007, 392, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Helvering, L.M.; Sharp, R.L.; Ou, X.; Geiser, A. Regulation of the promoters for the human bone morphogenetic protein 2 and 4 genes. Gene 2000, 256, 123–138. [Google Scholar] [CrossRef]
- Choi, K.-Y.; Lee, M.-H.; Kwon, T.-G.; Nah, H.-D.; Furuichi, T.; Komori, T.; Nam, S.-H.; Kim, Y.-J.; Kim, H.-J.; Ryoo, H.-M. Runx2 regulates FGF2-inducedBmp2 expression during cranial bone development. Dev. Dyn. 2005, 233, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Sand, J.P.; Kokorina, N.A.; Zakharkin, S.O.; Lewis, J.S.; Nussenbaum, B. BMP-2 Expression Correlates with Local Failure in Head and Neck Squamous Cell Carcinoma. Otolaryngol. Neck Surg. 2013, 150, 245–250. [Google Scholar] [CrossRef]
- Mundy, G.R. Metastasis to bone: Causes, consequences and therapeutic opportunities. Nat. Rev. Cancer 2002, 2, 584–593. [Google Scholar] [CrossRef]
- Dai, J.; Hall, C.L.; Escara-Wilke, J.; Mizokami, A.; Keller, J.M.; Keller, E.T. Prostate cancer induces bone metastasis through Wnt-induced bone morphogenetic protein-dependent and independent mechanisms. Cancer Res. 2008, 68, 5785–5794. [Google Scholar] [CrossRef] [Green Version]
- Alarmo, E.-L.; Korhonen, T.; Kuukasjärvi, T.; Huhtala, H.; Holli, K.; Kallioniemi, A. Bone morphogenetic protein 7 expression associates with bone metastasis in breast carcinomas. Ann. Oncol. 2008, 19, 308–314. [Google Scholar] [CrossRef]
- Micheau, C.; Boussen, H.; Klijanienko, J.; Cvitkovic, E.; Stosić, S.; Schwaab, G.; Eschwege, F.; Armand, J.-P. Bone marrow biopsies in patients with undifferentiated carcinoma of the nasopharyngeal type. Cancer 1987, 60, 2459–2464. [Google Scholar] [CrossRef]
- Ide, H.; Yoshida, T.; Matsumoto, N.; Aoki, K.; Osada, Y.; Sugimura, T.; Terada, M. Growth regulation of human prostate cancer cells by bone morphogenetic protein 2. Cancer Res. 1997, 57, 5022–5027. [Google Scholar] [PubMed]
- Kim, M.-J.; Park, S.Y.; Chang, H.R.; Jung, E.Y.; Munkhjargal, A.; Lim, J.-S.; Lee, M.-S.; Kim, Y. Clinical significance linked to functional defects in bone morphogenetic protein type 2 receptor, BMPR2. BMB Rep. 2017, 50, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.Y.; Lee, D.-H.; Lee, D.K.; Ahn, H.-J.; Kim, M.M.; Kim, S.J.; A Morton, R. Loss of expression of bone morphogenetic protein receptor type II in human prostate cancer cells. Oncogene 2004, 23, 7651–7659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.Y.; Lee, N.-H.; Lee, D.K.; Kim, B.-C.; Kim, H.T.; Leach, F.S.; Linehan, W.M.; A Morton, R.; Kim, S.J. Decreased expression of bone morphogenetic protein (BMP) receptor type II correlates with insensitivity to BMP-6 in human renal cell carcinoma cells. Clin. Cancer Res. 2003, 9, 6046–6051. [Google Scholar] [PubMed]
- Sneddon, J.B.; Zhen, H.H.; Montgomery, K.; van de Rijn, M.; Tward, A.D.; West, R.; Gladstone, H.; Chang, H.Y.; Morganroth, G.S.; Oro, A.E.; et al. Bone morphogenetic protein antagonist gremlin 1 is widely expressed by cancer-associated stromal cells and can promote tumor cell proliferation. Proc. Natl. Acad. Sci. USA 2006, 103, 14842–14847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.B.; Beppu, H.; Kawai, N.; Li, E.; Bloch, K.D. Bone Morphogenetic Protein (BMP) Type II Receptor Deletion Reveals BMP Ligand-specific Gain of Signaling in Pulmonary Artery Smooth Muscle Cells. J. Boil. Chem. 2005, 280, 24443–24450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ide, H.; Katoh, M.; Sasaki, H.; Yoshida, T.; Aoki, K.; Nawa, Y.; Osada, Y.; Sugimura, T.; Terada, M. Cloning of human bone morphogenetic protein type IB receptor (BMPR-IB) and its expression in prostate cancer in comparison with other BMPRs. Oncogene 1997, 14, 1377–1382. [Google Scholar] [CrossRef]
- Owens, P.; Pickup, M.W.; Novitskiy, S.V.; Chytil, A.; Gorska, A.E.; Aakre, M.E.; West, J.; Moses, H.L. Disruption of bone morphogenetic protein receptor 2 (BMPR2) in mammary tumors promotes metastases through cell autonomous and paracrine mediators. Proc. Natl. Acad. Sci. USA 2011, 109, 2814–2819. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.Y.; Lee, D.H.; Ahn, H.J.; Tokunaga, H.; Song, W.; Devereaux, L.M.; Jin, D.; Sampath, T.K.; A Morton, R. Expression of bone morphogenetic protein receptors type-IA, -IB and -II correlates with tumor grade in human prostate cancer tissues. Cancer Res. 2000, 60, 2840–2844. [Google Scholar]
- Raval, P.; Hsu, H.; Schneider, D.; Masuhara, K.; Anderson, H.; Sarras, M.; Bonewald, L. Expression of Bone Morphogenetic Proteins by Osteoinductive and Non-osteoinductive Human Osteosarcoma Cells. J. Dent. Res. 1996, 75, 1518–1523. [Google Scholar] [CrossRef]
- Yin, Q.; McBride, J.; Fewell, C.; Lacey, M.; Wang, X.; Lin, Z.; Cameron, J.E.; Flemington, E.K. MicroRNA-155 Is an Epstein-Barr Virus-Induced Gene That Modulates Epstein-Barr Virus-Regulated Gene Expression Pathways. J. Virol. 2008, 82, 5295–5306. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.; Wang, X.; Fewell, C.; Cameron, J.E.; Zhu, H.; Baddoo, M.; Lin, Z.; Flemington, E.K. MicroRNA miR-155 Inhibits Bone Morphogenetic Protein (BMP) Signaling and BMP-Mediated Epstein-Barr Virus Reactivation. J. Virol. 2010, 84, 6318–6327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.-X.; Ma, X.-Q.; Yang, L.; Wang, Z.-L.; Zeng, L.; Li, Z.-J.; Li, X.; Tang, M.; Yi, W.; Gong, J.-P.; et al. DNAzymes targeted to EBV-encoded latent membrane protein-1 induce apoptosis and enhance radiosensitivity in nasopharyngeal carcinoma. Cancer Lett. 2008, 265, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-C.; Chen, G.-H.; Chen, Y.-H.; Liao, W.-L.; Liu, C.-Y.; Chang, K.-P.; Chang, Y.-S.; Chen, S.-J. MicroRNA deregulation and pathway alterations in nasopharyngeal carcinoma. Br. J. Cancer 2009, 100, 1002–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, P.; Wu, J.; Zhang, J.; Zhao, S.; Zheng, H.; Gao, G.; Wei, L.; Li, Y. Viral Infection Induces Expression of Novel Phased MicroRNAs from Conserved Cellular MicroRNA Precursors. PLoS Pathog. 2011, 7, e1002176. [Google Scholar] [CrossRef] [PubMed]
- Gatto, G.; Rossi, A.; Rossi, D.; Kroening, S.; Bonatti, S.; Mallardo, M. Epstein–Barr virus latent membrane protein 1 trans-activates miR-155 transcription through the NF-κB pathway. Nucleic Acids Res. 2008, 36, 6608–6619. [Google Scholar] [CrossRef]
- Rahadiani, N.; Takakuwa, T.; Tresnasari, K.; Morii, E.; Aozasa, K. Latent membrane protein-1 of Epstein-Barr virus induces the expression of B-cell integration cluster, a precursor form of microRNA-155, in B lymphoma cell lines. Biochem. Biophys. Res. Commun. 2008, 377, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Park, S.H.; Yi, Y.; Choi, S.-G.; Lee, C.; Parks, W.T.; Cho, H.; de Caestecker, M.P.; Shaul, Y.; Roberts, A.B.; et al. The hepatitis B virus encoded oncoprotein pX amplifies TGF-beta family signaling through direct interaction with Smad4: Potential mechanism of hepatitis B virus-induced liver fibrosis. Genes Dev. 2001, 15, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Hu, H.; Li, S.; Dong, J.; Wang, X.; Wang, Y.; He, L.; He, Z.; Gao, Y.; Gao, S.-J.; et al. Oncogenic Herpesvirus KSHV Hijacks BMP-Smad1-Id Signaling to Promote Tumorigenesis. PLoS Pathog. 2014, 10, e1004253. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| ACVR1 | Forward: GCTGCCCACTAAAGGAAAAT Reverse: GCGAGCCACTGTTCTTTGTA |
| ACVR2B | Forward: TCCCTCACGGATTACCTCA Reverse: CCTCCTCAAAAGGCAGCA |
| BMP2 | Forward: CCTGAAACAGAGACCCACC Reverse: GCATTCTGATTCACCAACCT |
| BMPR1A | Forward: GTGGGTCTGGACTACCTT Reverse: GGGCACATCAACTTCATT |
| BMPR1B | Forward: CCACCCTAGACGCTAAAT Reverse: GCTCTCGTCCAACACTTCT |
| BMPR2 | Forward: CCTGATGTTCTGCCTACT Reverse: GCTCTTCTGGGCTTTGAT |
| EBNA1 | Forward: GGGTGGTTTGGAAAGCAT Reverse: TGGAAACCAGGGAGGCAAAT |
| GAPDH | Forward: GCCTCCTGCACCACCAACTG Reverse: CGACGCCTGCTTCACCACCTTCT |
| Smad1 | Forward: CTCTCCCACCAGCTCAGA Reverse: CACTAAGGCATTCGGCAT |
| Smad5 | Forward: CGCCTCCTCCTGCCTATA Reverse: GCTGCTGGGAATCTTACA |
| Smad9 | Forward: CGCCTACTATGAACTGAA Reverse: GGAAGCCGTGTTGATAGT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bridgewater, H.E.; Date, K.L.; O’Neil, J.D.; Hu, C.; Arrand, J.R.; Dawson, C.W.; Young, L.S. The Epstein-Barr Virus-Encoded EBNA1 Protein Activates the Bone Morphogenic Protein (BMP) Signalling Pathway to Promote Carcinoma Cell Migration. Pathogens 2020, 9, 594. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9070594
Bridgewater HE, Date KL, O’Neil JD, Hu C, Arrand JR, Dawson CW, Young LS. The Epstein-Barr Virus-Encoded EBNA1 Protein Activates the Bone Morphogenic Protein (BMP) Signalling Pathway to Promote Carcinoma Cell Migration. Pathogens. 2020; 9(7):594. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9070594
Chicago/Turabian StyleBridgewater, Hannah E., Kathryn L. Date, John D. O’Neil, Chunfang Hu, John R. Arrand, Christopher W. Dawson, and Lawrence S. Young. 2020. "The Epstein-Barr Virus-Encoded EBNA1 Protein Activates the Bone Morphogenic Protein (BMP) Signalling Pathway to Promote Carcinoma Cell Migration" Pathogens 9, no. 7: 594. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9070594