Mycobacterium pseudoshottsii in Mediterranean Fish Farms: New Trouble for European Aquaculture?

 , ,
, ,  ,
,  , ,
, , 
 ,
,
Abstract
:1. Introduction
2. Results
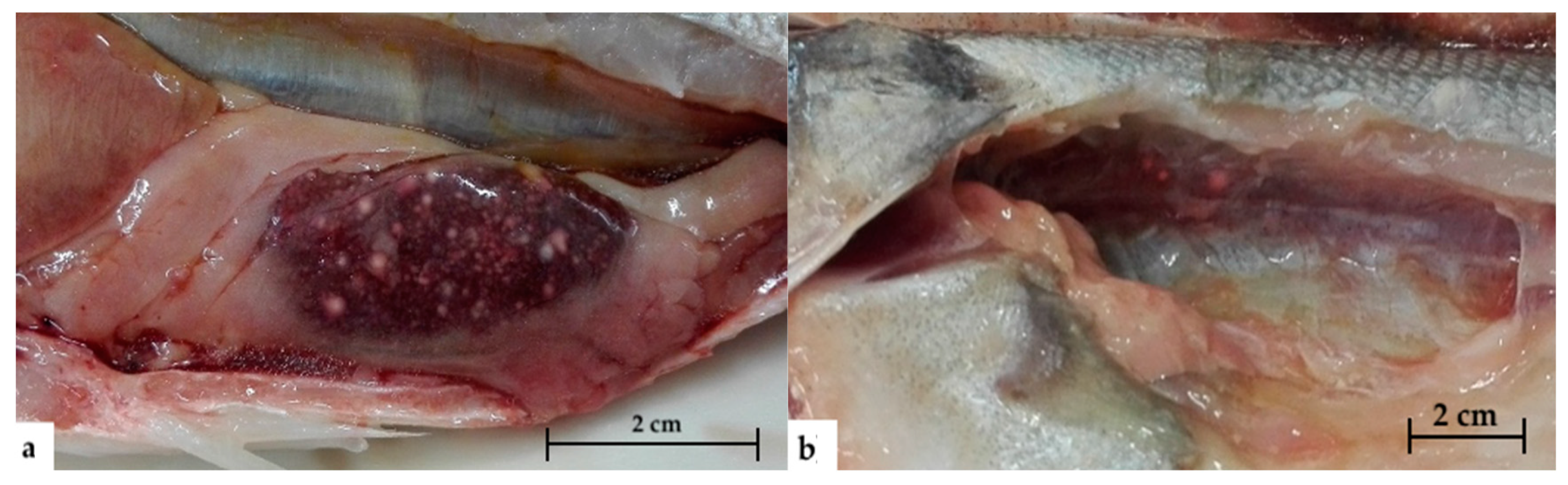
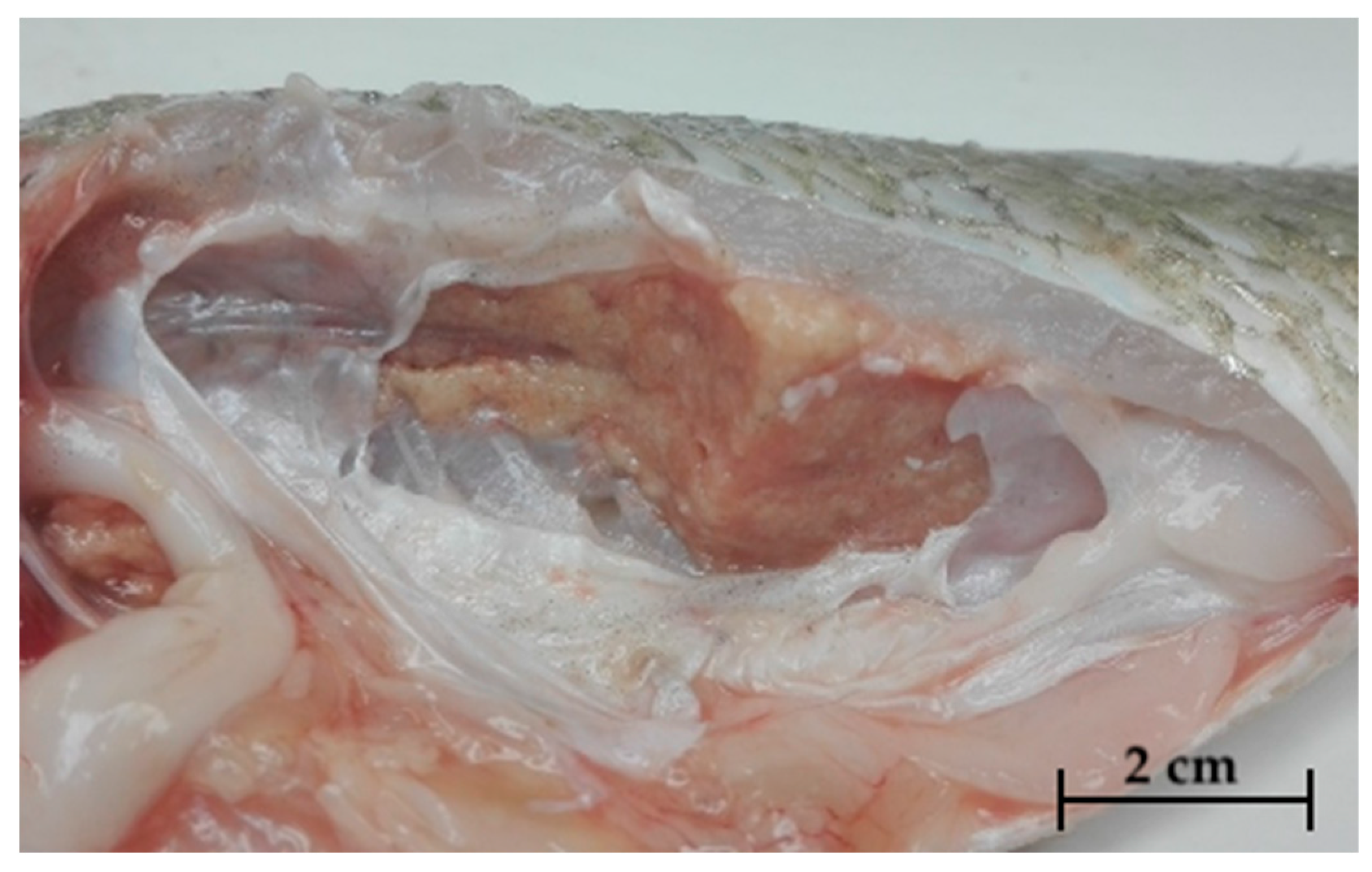
2.1. Gross Pathology
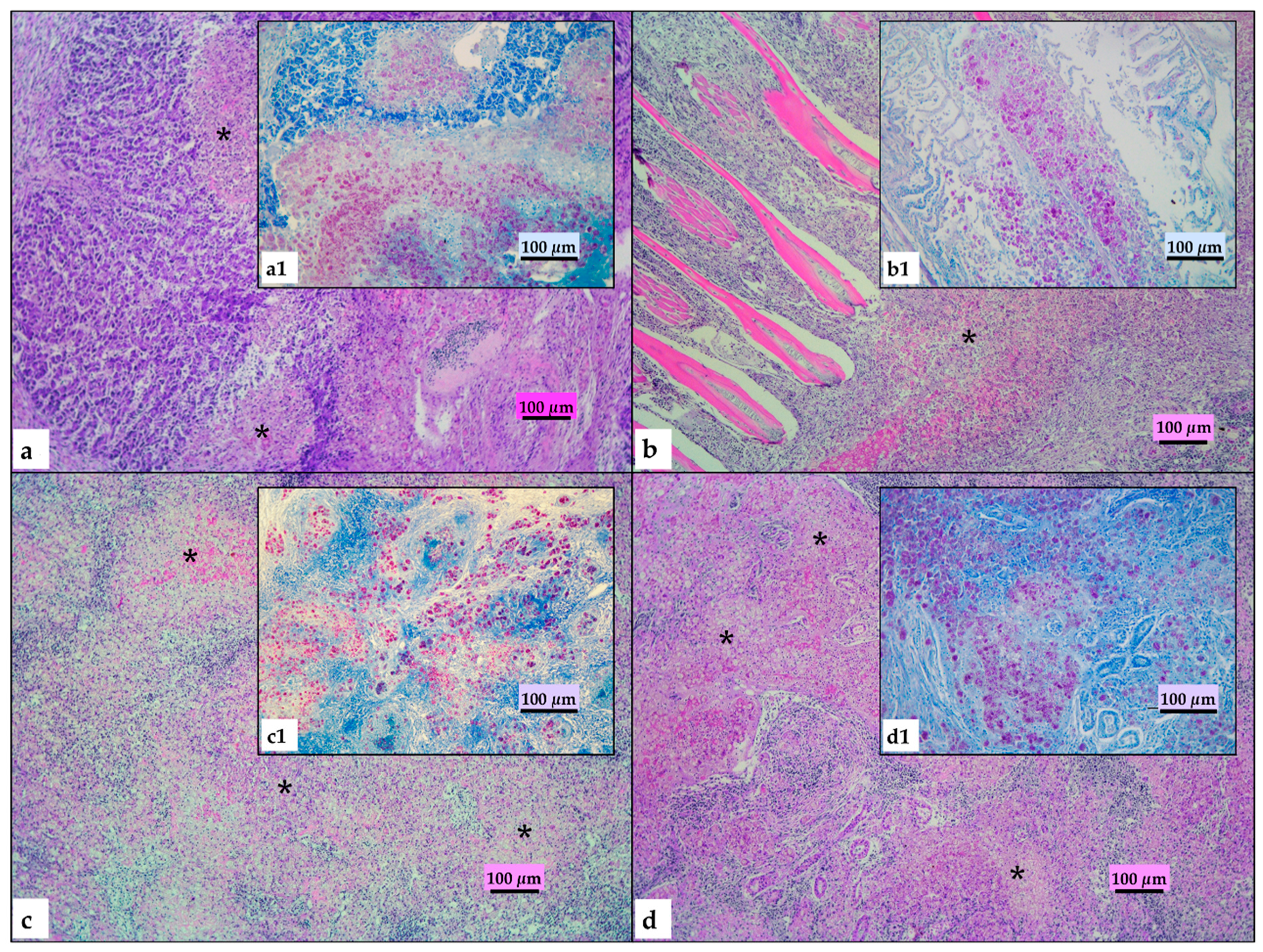
2.2. Histopathological Findings
2.3. Mycobacteria Isolation and Species Identification
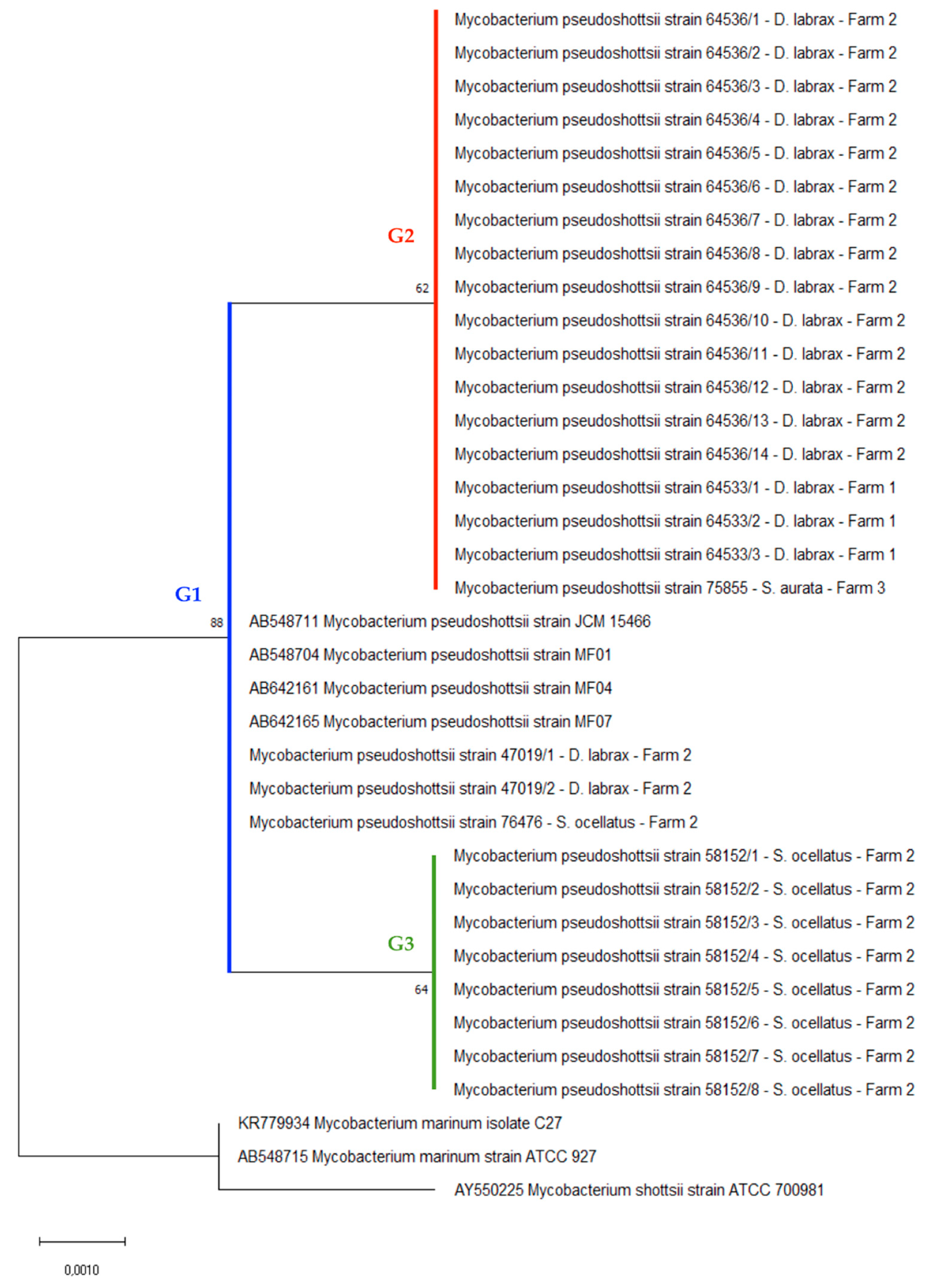
2.4. Phylogenetic Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fish Sampling
5.2. Anatomopathological Examination
5.3. Histopathological Examination
5.4. Culture Exams
5.5. DNA Extraction and PCR
5.6. Phylogenetic Analysis with hsp65 Gene Sequences
Author Contributions
Funding
Conflicts of Interest
References
- Kaattari, I.M.; Rhodes, M.W.; Kaattari, S.L.; Shotts, E.B. The evolving story of Mycobacterium tuberculosis clade members detected in fish. J. Fish. Dis. 2006, 29, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Hashish, E.; Merwad, A.; Elgaml, S.; Amer, A.; Kamal, H.; Elsadek, A.; Marei, A.; Sitohy, M. Mycobacterium marinum infection in fish and man: Epidemiology, pathophysiology and management; a review. Vet. Q. 2018, 38, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decostere, A.; Hermans, K.; Haesebrouck, F. Piscine mycobacteriosis: A literature review covering the agent and the disease it causes in fish and humans. Vet. Microbiol. 2004, 99, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, D.T.; Rhodes, M.W. Mycobacteriosis in fishes: A review. Vet. J. 2009, 180, 33–47. [Google Scholar] [CrossRef]
- Jacobs, J.; Stine, C.; Baya, A.; Kent, M.L. A review of mycobacteriosis in marine fish. J. Fish. Dis. 2009, 32, 119–130. [Google Scholar] [CrossRef]
- Tortoli, E. Impact of genotypic studies on mycobacterial taxonomy: The new mycobacteria of the 1990s. Clin. Microbiol. Rev. 2003, 16, 319–354. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, M.W.; Kator, H.; Kotob, S.; van Berkum, P.; Kaattari, I.; Vogelbein, W.; Floyd, M.M.; Butler, W.R.; Quinn, F.D.; Ottinger, C.; et al. A unique Mycobacterium species isolated from an epizootic of striped bass (Morone saxatilis). Emerg. Infect. Dis. 2001, 7, 896–899. [Google Scholar] [CrossRef]
- Rhodes, M.W.; Kator, H.; Kotob, S.; van Berkum, P.; Kaattari, I.; Vogelbein, W.; Quinn, F.; Floyd, M.M.; Butler, W.R.; Ottinger, C.A. Mycobacterium shottsii sp. nov., a slowly growing species isolated from Chesapeake Bay striped bass (Morone saxatilis). Int. J. Syst. Evol. Microbiol. 2003, 53, 421–424. [Google Scholar] [CrossRef]
- Rhodes, M.W.; Kator, H.; Kaattari, I.; Gauthier, D.; Vogelbein, W.; Ottinger, C.A. Isolation and characterization of mycobacteria from striped bass Morone saxatilis from the Chesapeake Bay. Dis. Aquat. Organ. 2004, 61, 41–51. [Google Scholar] [CrossRef]
- Rhodes, M.W.; Kator, H.; McNabb, A.; Deshayes, C.; Reyrat, J.M.; Brown-Elliott, B.A.; Wallace, R.J.J.; Trott, K.A.; Parker, J.M.; Lifland, B.; et al. Mycobacterium pseudoshottsii sp. nov., a slowly growing chromogenic species isolated from Chesapeake Bay striped bass (Morone saxatilis). Int. J. Syst. Evol. Microbiol. 2005, 55, 1139–1147. [Google Scholar] [CrossRef] [Green Version]
- Mve-Obiang, A.; Lee, R.E.; Umstot, E.S.; Trott, K.A.; Grammer, T.C.; Parker, J.M.; Ranger, B.S.; Grainger, R.; Mahrous, E.A.; Small, P.L.C. A newly discovered mycobacterial pathogen isolated from laboratory colonies of Xenopus species with lethal infections produces a novel form of mycolactone, the Mycobacterium ulcerans macrolide toxin. Infect. Immun. 2005, 73, 3307–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stine, C.B.; Jacobs, J.M.; Rhodes, M.R.; Overton, A.; Fast, M.; Baya, A.M. Expanded range and new host species of Mycobacterium shottsii and M. pseudoshottsii. J. Aquat Anim Health. 2009, 21, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, D.T.; Reece, K.S.; Xiao, J.; Rhodes, M.W.; Kator, H.I.; Latour, R.J.; Bonzek, C.F.; Hoenig, J.M.; Vogelbein, W.K. Quantitative PCR assay for Mycobacterium pseudoshottsii and Mycobacterium shottsii and application to environmental samples and fishes from the Chesapeake Bay. Appl. Environ. Microbiol. 2010, 76, 6171–6179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanaga, K.; Hoshino, Y.; Hattori, Y.; Yamamoto, A.; Wada, S.; Hatai, K.; Makino, M.; Ishii, N. Mycobacterium pseudoshottsii isolated from 24 farmed fishes in western Japan. J. Vet. Med. Sci. 2012, 74, 275–278. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, N.M.; do Vale, A.; Sousa, M.J.; Silva, M.T. Mycobacterial infection in farmed turbot Scophthalmus maximus. Dis Aquat Organ. 2002, 52, 87–91. [Google Scholar] [CrossRef]
- Ucko, M.; Colorni, A.; Kvitt, H.; Diamant, A.; Zlotkin, A.; Knibb, W.R. Strain variation in Mycobacterium marinum fish isolates. Appl. Environ. Microbiol. 2002, 68, 5281–5287. [Google Scholar] [CrossRef] [Green Version]
- Salogni, C.; Zanoni, M.; Covi, M.; Pacciarini, M.L.; Alborali, L. Outbreak of Mycobacterium marinum in farmed rainbow trout (Oncorhynchus mykiss) and brown trout (Salmo trutta). Ittiopatologia. 2007, 4, 227–237. [Google Scholar]
- Bozzetta, E.; Varello, K.; Giorgi, I.; Fioravanti, M.L.; Pezzolato, M.; Zanoni, R.G.; Prearo, M. Mycobacterium marinum infection in a hybrid striped bass farm in Italy. J. Fish. Dis. 2010, 33, 781 –785. [Google Scholar] [CrossRef]
- Salati, F.; Meloni, M.; Fenza, A.; Angelucci, G.; Colorni, A.; Orru, G. A sensitive FRET probe assay for the selective detection of Mycobacterium marinum in fish. J. Fish. Dis. 2010, 33, 47–56. [Google Scholar] [CrossRef]
- Antuofermo, E.; Pais, A.; Polinas, M.; Cubeddu, T.; Righetti, M.; Sanna, M.A.; Prearo, M. Mycobacteriosis caused by Mycobacterium marinum in reared mullets: First evidence from Sardinia (Italy). J. Fish. Dis. 2017, 40, 327–337. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Avsever, M.L.; Çavuşoğlu, C.; Eskiizmirliler, S.; Türe, M.; Korun, J.; Çamkerten, İ. First isolation of Mycobacterium marinum from sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus auratus) cultured in Turkey. Bull. Eur. Ass. Fish. Pathol. 2016, 36, 193. [Google Scholar]
- Regev, Y.; Davidovich, N.; Berzak, R.; Lau, S.C.K.; Scheinin, A.P.; Tchernov, D.; Morick, D. Molecular Identification and Characterization of Vibrio Species and Mycobacterium Species in Wild and Cultured Marine Fish from the Eastern Mediterranean Sea. Microorganisms 2020, 8, 863. [Google Scholar] [CrossRef] [PubMed]
- Davidovich, N.; Pretto, T.; Sharon, G.; Zilberg, D.; Blum, S.E.; Baider, Z.; Edery, N.; Morick, D.; Grossman, R.; Kaidar-Shwartz, H.; et al. Cutaneous appearance of mycobacteriosis caused by Mycobacterium marinum, affecting gilthead seabream (Sparus aurata) cultured in recirculating aquaculture systems. Aquaculture 2020, 528, 735507. [Google Scholar] [CrossRef]
- Diamant, A.; Banet, A.; Ucko, M.; Colorni, A.; Knibb, W.; Kvitt, H. Mycobacteriosis in wild rabbitfish Siganus rivulatus associated with cage farming in the Gulf of Eilat, Red Sea. Dis. Aquat. Organ. 2000, 39, 211–219. [Google Scholar] [CrossRef]
- Chao, L. Sciaenops ocellatus. The IUCN Red List of Threatened Species 2015: E.T193270A49226782. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2015-2.RLTS.T193270A49226782.en (accessed on 22 May 2020).
- Axén, C.; Vendramin, N.; Toffan, A. Outbreak of mortality associated with Acipenser Iridovirus European (AcIV-E) detection in Siberian Sturgeon (Acipenser baerii) farmed in Sweden. Fishes 2018, 3, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Menconi, V.; Pastorino, P.; Cavazza, G.; Santi, M.; Mugetti, D.; Zuccaro, G.; Prearo, M. The role of live fish trade in the translocation of parasites: The case of Cystidicola farionis in farmed rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2019, 27, 1667–1671. [Google Scholar] [CrossRef]
- Varello, K.; Righetti, M.; Meistro, S.; Gastaldi, D.; Burioli, E.A.V.; Pezzolato, M.; Foglini, C.; Bozzetta, E.; Prearo, M. Episodio di micobatteriosi diffusiva e generalizzata in branzini (Dicentrarchus labrax): Aspetti anatomo-istopatologici. In Proceedings of the XXI Convegno Nazionale, S.I.P.I. (Società Italiana Patologia Ittica), Chioggia, Italy, 8–9 October 2015; p. 45. Available online: https://www.sipi-online.it/convegni/2015/atti.pdf (accessed on 27 July 2020).
- Magee, J.G.; Ward, A.C. Family III: Mycobacteriaceae Chester 1897. In Bergey’s Manual of Systematics Bacteriology, 2nd ed.; Kämpfer, P., Busse, H.J., Trujillo, M.E., Suzuki, K., Ludwig, W., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2012; Volume 5, pp. 312–375. [Google Scholar]
- Rónai, Z.; Eszterbauer, E.; Csivincsik, Á.; Guti, C.F.; Dencső, L.; Jánosi, S.; Dán, Á. Detection of wide genetic diversity and several novel strains among non-avium nontuberculosis-like mycobacteria isolated from farmed and wild animals in Hungary. J. Appl. Microbiol. 2016, 121, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Beye, M.; Fahsi, N.; Raoult, D.; Fournier, P.E. Careful use of 16S rRNA gene sequence similarity values for the identification of Mycobacterium species. New Microbes New Infect. 2018, 22, 24–29. [Google Scholar] [CrossRef]
- McNabb, A.; Eisler, D.; Adie, K.; Amos, M.; Rodrigues, M.; Stephens, G.; Black, W.A.; Isaac-Renton, J. Assessment of partial sequencing of the 65-kilodalton heat shock protein gene (hsp65) for routine identification of Mycobacterium species isolated from clinical sources. J. Clin. Microbiol. 2004, 42, 3000–3011. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, S.H.; Shim, T.S.; Kim, M.; Bai, G.H.; Park, Y.G.; Lee, S.H.; Chae, G.T.; Cha, C.Y.; Kook, Y.H.; et al. Differentiation of Mycobacterium species by analysis of the heat-shock protein 65 gene (hsp65). Int. J. Sist. Evol. Microbiol. 2005, 55, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Schinnick, T. The 65-kilodalton antigen of Mycobactenium tuberculosis. J. Bacteriol. 1987, 169, 1080–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, J.M.; Howard, D.W.; Rhodes, M.R.; Newman, M.W.; May, E.B.; Harrell, R.M. Historical presence (1975–1985) of mycobacteriosis in Chesapeake Bay striped bass Morone saxatilis. Dis. Aquat. Organ. 2009, 85, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranger, B.S.; Mahrous, E.A.; Mosi, L.; Adusumilli, S.; Lee, R.E.; Colorni, A.; Rhodes, M.; Small, P.L.C. Globally distributed mycobacterial fish pathogens produce a novel plasmid-encoded toxic macrolide, mycolactone F. Infect. Immun. 2006, 74, 6037–6045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugetti, D.; Pastorino, P.; Menconi, V.; Messina, M.; Masoero, L.; Ceresa, L.; Pedron, C.; Prearo, M. Two new sturgeon species are susceptible to Acipenser Iridovirus European (AcIV-E) infection. Pathogens 2020, 9, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthier, D.T.; Rhodes, M.W.; Vogelbein, W.K.; Kator, H.; Ottinger, C.A. Experimental mycobacteriosis in striped bass Morone saxatilis. Dis. Aquat. Organ. 2003, 54, 105–117. [Google Scholar] [CrossRef]
- Talaat, A.M.; Reimschuessel, R.; Trucksis, M. Identification of mycobacteria infecting fish to the species level using polymerase chain reaction and restriction enzyme analysis. Vet. Microbiol. 1997, 58, 229–237. [Google Scholar] [CrossRef]
- Telenti, A.; Marchesi, F.; Balz, M.; Bally, F.; Böttger, E.C.; Bodmer, T. Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J. Clin. Microbiol. 1993, 31, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Adékambi, T.; Colson, P.; Drancourt, M. rpoB-based identification of nonpigmented and late-pigmenting rapidly growing mycobacteria. J. Clin. Microbiol. 2003, 41, 5699–5708. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega, Accurate Alignment of Very Large Numbers of Sequences. In Multiple Sequence Alignment Methods Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; Volume 1079, pp. 105–116. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018. [Google Scholar] [CrossRef]
- Aronson, J.D. Spontaneous tuberculosis in soft water fish. J. Infect. Dis. 1926, 39, 315–320. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish Species | |||||||
|---|---|---|---|---|---|---|---|
| European Sea Bass (D. labrax) | Red Drum (S. ocellatus) | Gilthead Sea Bream (S. aurata) | |||||
| Pos | Neg | Pos | Neg | Pos | Neg | ||
| Farm | Farm 1 | 3 | 19 | - | - | - | - |
| Farm 2 | 16 (a) | 32 (b) | 9 (c) | 28 (d) | |||
| Farm 3 | - | - | - | - | 1 | 9 | |
| Fish Species | |||||
|---|---|---|---|---|---|
| European Sea bass (D. labrax) | Red Drum (S. ocellatus) | Gilthead Sea Bream (S. aurata) | Total | ||
| Farm | Farm 1 | 22 | - | - | 22 |
| Farm 2 | 48 (a) | 37 (b) | - | 85 | |
| Farm 3 | - | - | 10 | 10 | |
| Total | 70 | 37 | 10 | 117 | |
| Gene | Primer Forward | Primer Reverse | Length (bp) | Reference |
|---|---|---|---|---|
| 16S rRNA | T39 (GCGAACGGGTGAGTAACACG) | TI3 (TGCACACAGGCCACAAGGGA) | 924 | [40] |
| hsp65 | Tb11 (ACCAACGATGGTGTGTCCAT) | Tb12 (CTTGTCGAACCGCATACCCT) | 441 | [41] |
| rpoB | MycoF (GGCAAGGTCACCCCGAAGGG) | MycoR (AGCGGCTGCTGGGTGATCATC) | 723 | [42] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugetti, D.; Varello, K.; Gustinelli, A.; Pastorino, P.; Menconi, V.; Florio, D.; Fioravanti, M.L.; Bozzetta, E.; Zoppi, S.; Dondo, A.; et al. Mycobacterium pseudoshottsii in Mediterranean Fish Farms: New Trouble for European Aquaculture? Pathogens 2020, 9, 610. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080610
Mugetti D, Varello K, Gustinelli A, Pastorino P, Menconi V, Florio D, Fioravanti ML, Bozzetta E, Zoppi S, Dondo A, et al. Mycobacterium pseudoshottsii in Mediterranean Fish Farms: New Trouble for European Aquaculture? Pathogens. 2020; 9(8):610. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080610
Chicago/Turabian StyleMugetti, Davide, Katia Varello, Andrea Gustinelli, Paolo Pastorino, Vasco Menconi, Daniela Florio, Maria Letizia Fioravanti, Elena Bozzetta, Simona Zoppi, Alessandro Dondo, and et al. 2020. "Mycobacterium pseudoshottsii in Mediterranean Fish Farms: New Trouble for European Aquaculture?" Pathogens 9, no. 8: 610. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080610