Dolphins Stranded along the Tuscan Coastline (Central Italy) of the “Pelagos Sanctuary”: A Parasitological Investigation



Abstract
:1. Introduction
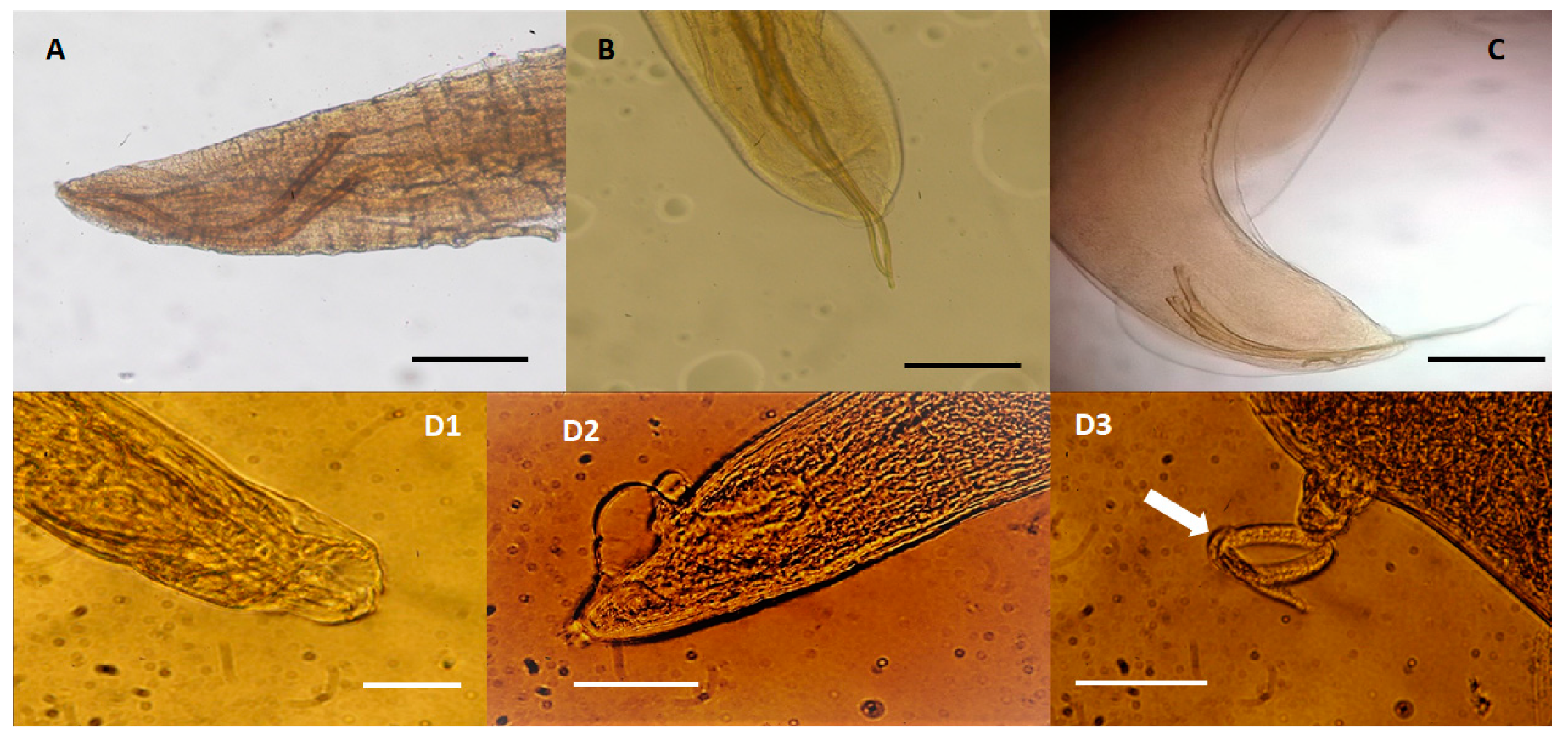
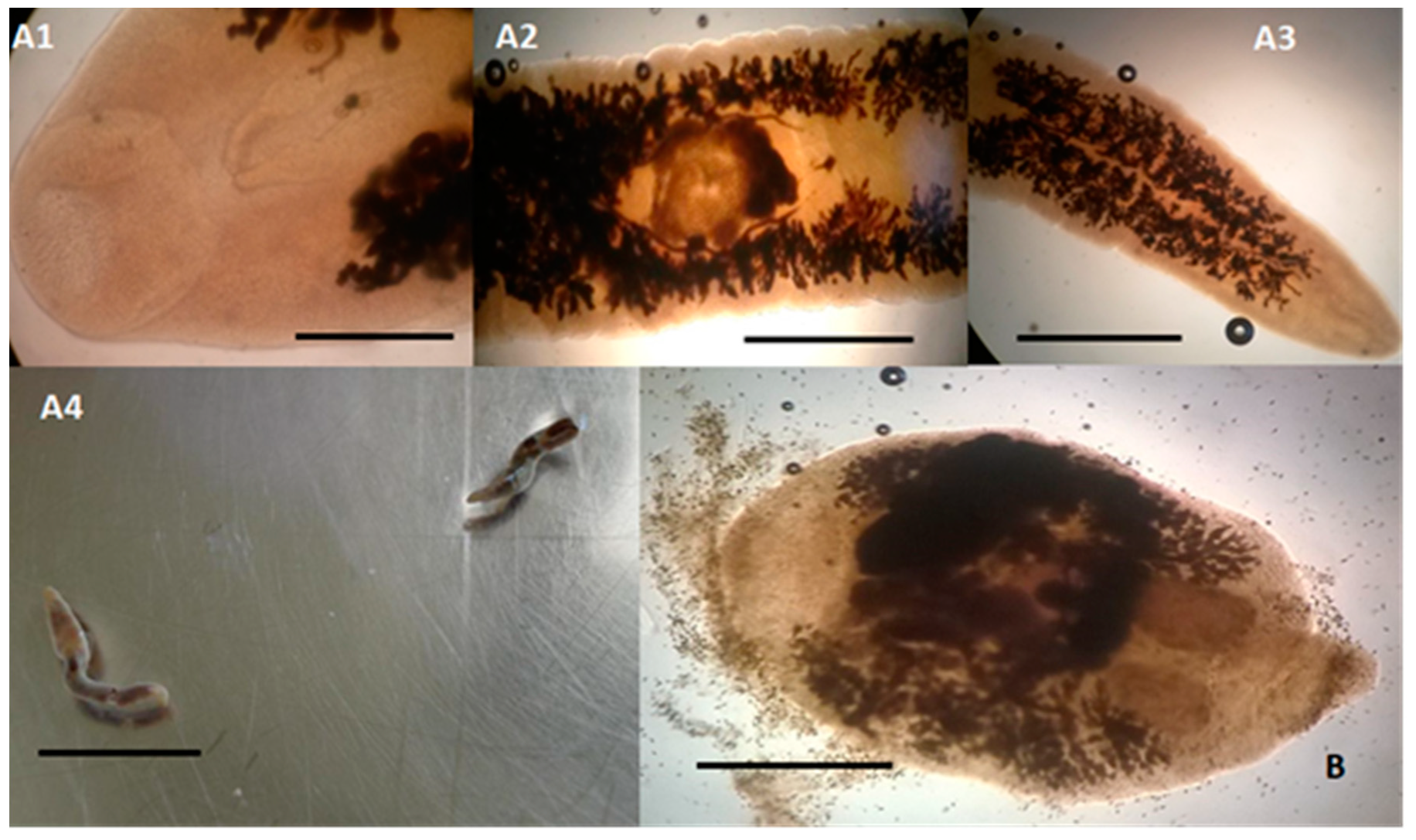
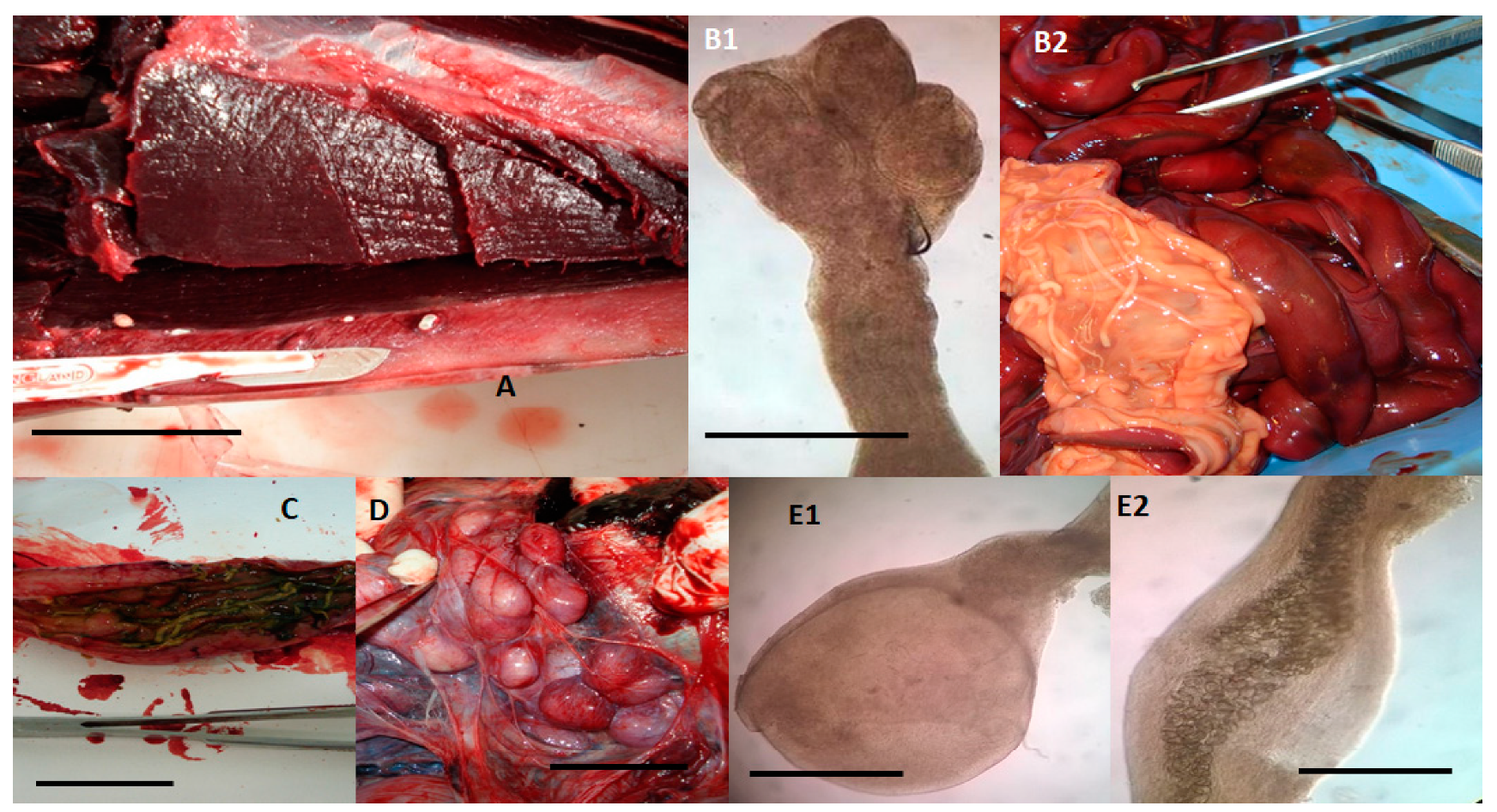
2. Results and Discussion
3. Materials and Methods
3.1. Animals
3.2. Parasitological Examination
3.3. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Panigada, S.; Lauriano, G.; Burt, L.; Pierantonio, N.; Donovan, G. Monitoring winter and summer abundance of cetaceans in the Pelagos Sanctuary (northwestern Mediterranean Sea) through aerial surveys. PLoS ONE 2011, 6, e22878. [Google Scholar] [CrossRef] [PubMed]
- Notarbartolo-di-Sciara, G.; Agardy, T.; Hyrenbach, D.; Scovazzi, T.; Van Klaveren, P. The Pelagos Sanctuary for Mediterranean marine mammals. Aquat. Conserv. 2008, 18, 367–391. [Google Scholar] [CrossRef]
- Pelagos Sanctuary. Available online: https://www.sanctuaire-pelagos.org/en/about-us/area-of-application-and-coastal-municipalities (accessed on 10 June 2020).
- Pennino, M.G.; Arcangeli, A.; Prado Fonseca, V.; Campana, I.; Pierce, G.J.; Rotta, A.; Bellido, J.M. A spatially explicit risk assessment approach: Cetaceans and marine traffic in the Pelagos Sanctuary (Mediterranean Sea). PLoS ONE 2017, 12, e0179686. [Google Scholar] [CrossRef] [PubMed]
- Balbuena, J.A.; Simpkin, A. Role of Crassicauda sp. in natural mortality of pantropical spotted dolphins Stenella attenuata: A reassessment. Dis. Aquat. Organ. 2014, 108, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Lassalle, G.; Gascuel, D.; Le Loc’h, F.; Lobry, J.; Pierce, G.J.; Ridoux, V.; Begona Santos, M.; Spitz, J.; Niquil, N. An ecosystem approach for the assessment of fisheries impacts on marine top predators: The bay of Biscay case study. ICES J. Mar. Sci. 2012, 69, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Lauriano, G.; Pierantonio, N.; Donovan, G.; Panigada, S. Abundance and distribution of Tursiops truncatus in the Western Mediterranean Sea: An assessment towards the Marine Strategy Framework Directive requirements. Mar. Environ. Res. 2014, 100, 86–93. [Google Scholar] [CrossRef]
- Mazzariol, S.; Centelleghe, C.; Cozzi, B.; Povinelli, M.; Marcer, F.; Ferri, N.; Di Francesco, G.; Badagliacca, P.; Profeta, F.; Olivieri, V.; et al. Multidisciplinary studies on a sick-leader syndrome-associated mass stranding of sperm whales (Physeter macrocephalus) along the Adriatic coast of Italy. Sci. Rep. 2018, 8, 11577. [Google Scholar] [CrossRef]
- Hrabar, J.; Bočina, I.; Gudan Kurilj, A.; Đuras, M.; Mladineo, I. Gastric lesions in dolphins stranded along the Eastern Adriatic coast. Dis. Aquat. Organ. 2017, 125, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Casalone, C.; Mazzariol, S.; Pautasso, A.; Di Guardo, G.; Di Nocera, F.; Lucifora, G.; Ligios, C.; Franco, A.; Fichi, G.; Cocumelli, C.; et al. Cetacean strandings in Italy: An unusual mortality event along the Tyrrhenian Sea coast in 2013. Dis. Aquat. Organ. 2014, 109, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Raga, J.A.; Aznar, F.J.; Balbuena, J.A.; Fernandez, M. Parasites. In Encyclopedia of Marine Mammals, 2nd ed.; Perrin, W.F., Wursig, B., Thewissen, H.G.M., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 821–830. [Google Scholar]
- Oliveira, J.B.; Morales, J.A.; González-Barrientos, R.C.; Hernández-Gamboa, J.; Hernández-Mora, G. Parasites of cetaceans stranded on the Pacific coast of Costa Rica. Vet. Parasitol. 2011, 182, 319–328. [Google Scholar] [CrossRef]
- Carvalho, V.L.; Bevilaqua, C.M.; Iñiguez, A.M.; Mathews-Cascon, H.; Bezerra Ribeiro, F.; Bezerra Pessoa, L.M.; Oliveira de Meirelles, A.C.; Gomes Borges, J.C.; Marigo, J.; Soares, L.; et al. Metazoan parasites of cetaceans off the northeastern coast of Brazil. Vet. Parasitol. 2010, 173, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R.; Blasco-Costa, I.; Randhawa, H.S. Integrating parasitology and marine ecology: Seven challenges towards greater synergy. J. Sea Res. 2016, 113, 3–10. [Google Scholar] [CrossRef]
- Lehnert, K.; Randhawa, H.; Poulin, R. Metazoan parasites from odontocetes of New Zealand: New records. Parasitol. Res. 2017, 116, 2861–2868. [Google Scholar] [CrossRef]
- Guimarães, J.P.; Febronio, A.M.; Vergara-Parente, J.E.; Werneck, M.R. Lesions associated with Halocercus brasiliensis Lins de Almeida, 1933 in the lungs of dolphins stranded in the Northeast of Brazil. J. Parasitol. 2015, 101, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Delgado, J.; Fernández, A.; Sierra, E.; Sacchini, S.; Andrada, M.; Vela, A.I.; Quesada-Canales, O.; Paz, Y.; Zucca, D.; Groch, K.; et al. Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006–2012). PLoS ONE 2018, 13, e0204444. [Google Scholar] [CrossRef] [Green Version]
- Obusan, M.C.M.; Villanueva, R.M.D.; Siringan, M.A.T.; Rivera, W.L.; Aragones, L.V. Leptospira spp. and Toxoplasma gondii in stranded representatives of wild cetaceans in the Philippines. BMC Vet. Res. 2019, 15, 372. [Google Scholar] [CrossRef]
- Pretti, C.; Mancianti, F.; Nardoni, S.; Ariti, G.; Monni, G.; Di Bello, D.; Marsili, S.; Papini, R. Detection of Toxoplasma gondii infection in dolphins stranded along the Tuscan coast, Italy. Rev. Med. Vet. 2010, 161, 428–431. [Google Scholar]
- Mateu, P.; Raga, J.A.; Fernandez, M.; Aznar, F.J. Intestinal helminth fauna of striped dolphins (Stenella coeruleoalba) in the western Mediterranean: No effects of host body length, age and sex. Mar. Mam. Sci. 2014, 30, 961–977. [Google Scholar] [CrossRef]
- Quinones, R.; Giovannini, A.; Raga, J.A.; Fernandez, M. Intestinal helminth fauna of bottlenose dolphin Tursiops truncatus and common Dolphin Delphinus delphis from the western mediterranean. J. Parasitol. 2013, 99, 576–579. [Google Scholar] [CrossRef]
- Aznar, F.J.; Fognani, P.; Balbuena, J.A.; Pietrobelli, M.; Raga, J.A. Distribution of Pholeter gastrophilus (Digenea) within the stomach of four odontocete species: The role of the diet and digestive physiology of hosts. Parasitology 2006, 133, 369–380. [Google Scholar] [CrossRef]
- Di Guardo, G.; Agrimi, U.; Morelli, L.; Cardeti, G.; Terracciano, G.; Kennedy, S. Postmortem investigations on cetaceans found stranded on the coasts of Italy between 1990 and 1993. Vet. Rec. 1995, 136, 439. [Google Scholar] [CrossRef]
- Manfredi, M.T.; Gazzonis, A.L.; Merella, P.; Garippa, G.; Musella, V. Cetacei dei Mari Italiani: Diffusione, Spiaggiamenti e Problematiche Sanitarie. 2012. Available online: http://www.parassitologia.unina.it/wp-content/themes/para/mappe/parassiti16.pdf (accessed on 19 May 2020).
- Mazzariol, S.; Marrucchella, G.; Di Guardo, G.; Podestà, M.; Olivieri, V.; Colangelo, P.; Kennedy, S.; Castagnaro, M.; Cozzi, B. Post-mortem findings in cetaceans stranded along Italian Adriatic Sea coastline (2000–2006). In Proceedings of the International Whaling Commission’s 59th Annual Meeting, Anchorage, AK, USA, 28–31 May 2007. [Google Scholar]
- Measures, L.N. Lungworms of marine mammals. In Parasitic Diseases of Wild Mammals, 2nd ed.; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Iowa States University Press: Ames, IA, USA, 2001; Chapter 10; pp. 279–300. [Google Scholar]
- Lambertsen, R.H. Crassicaudosis: A parasitic disease threatening the health and population recovery of large baleen whales. Rev. Sci. Tec. 1992, 11, 1131–1141. [Google Scholar] [CrossRef]
- Blazekovic, K.; Pleic, I.L.; Duras, M.; Gomercic, T.; Mladineo, I. Three Anisakis spp. isolated from toothed whales stranded along the eastern Adriatic Sea coast. Int. J. Parasitol. 2015, 5, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Van Beurden, S.J.; IJsseldijk, L.L.; Cremers, H.J.W.M.; Gröne, A.; Verheije, M.H.; Begeman, L. Anisakis spp. induced granulomatous dermatitis in a harbour porpoise Phocoena phocoena and a bottlenose dolphin Tursiops truncates. Dis. Aquat. Org. 2015, 112, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dailey, M.D. Parasitic disease. In CRC Handbook of Marine Mammals Medicine: Health, Disease and Rehabilitation, 2nd ed.; Dierauf, L.A., Gulland, F.M.D., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 2001; pp. 357–379. [Google Scholar]
- Romero, M.A.; Fernández, M.; Dans, S.L.; García, N.A.; González, R.; Crespo, E.A. Gastrointestinal parasites of bottlenose dolphins Tursiops truncatus from the extreme Southwestern Atlantic, with notes on diet composition. Dis. Aquat. Org. 2014, 108, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaber, J.R.; Perez, J.; Arbelo, M.; Zafra, R.; Fernandez, A. Pathological and immunohistochemical study of gastrointestinal lesions in dolphins stranded in the Canary Islands. Vet. Rec. 2006, 159, 410–414. [Google Scholar] [CrossRef]
- Gibson, D.I.; Harri, E.A.; Bray, R.A.; Jepson, P.D.; Kuiken, T.; Baker, J.R.; Simpson, V.R. A survey of the helminth parasites of cetaceans stranded on the coast of England and Wales during the period 1990–1994. J. Zool. 1998, 244, 563–574. [Google Scholar] [CrossRef]
- Aznar, F.J.; Agusti, C.; Littlewood, D.T.J.; Raga, J.A.; Olson, P.D. Insight into the role of cetaceans in the life cycle of the Tetraphyllideans (Platyhelminthes: Cestoda). Int. J. Parasitol 2007, 37, 243–255. [Google Scholar] [CrossRef]
- Khalil, L.F.; Jones, A.; Bray, R.A. Keys to the Cestode Parasites of Vertebrates; Commonwealth Agricultural Bureaux: Wallingford, UK, 1994; p. 768. [Google Scholar]
- Hoberg, E.P.; Daoust, P.Y.; McBurney, S. Bolbosoma capitatum and Bolbosoma sp. (Acanthocephala) from sperm whales (Physeter macrocephalus) stranded on Prince Edward Island, Canada. Proc. Helminthol. Soc. Wash. 1993, 60, 205–210. [Google Scholar]
- Costa, G.; Chubb, J.C.; Veltkamp, C.J. Cystacanths of Bolbosoma vasculosum in the black scabbard fish Aphanopus carbo, oceanic horse mackerel Trachurus picturatus and common dolphin Delphinus delphis from Madeira, Portugal. J. Helminthol. 2000, 74, 113–120. [Google Scholar] [CrossRef]
- Reboredo-Fernández, A.; Ares-Mazás, E.; Martínez-Cedeira, J.A.; Romero-Suances, R.; Cacciò, S.M.; Gómez-Couso, H. Giardia and Cryptosporidium in cetaceans on the European Atlantic coast. Parasitol. Res. 2015, 114, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Kleinertz, S.; Hermosilla, C.; Ziltener, A.; Kreicker, S.; Hirzmann, J.; Abdel-Ghaffar, F.; Taubert, A. Gastrointestinal parasites of free-living Indo-Pacific bottlenose dolphins (Tursiops aduncus) in the Northern Red Sea, Egypt. Parasitol. Res. 2014, 113, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Fair, P.A.; Bossartf, G.D.; Santin, M. Examination of naturally exposed bottlenose dolphins (Tursiops truncatus) for Microsporidia, Cryptosporidium, and Giardia. J. Parasitol. 2008, 94, 143–147. [Google Scholar] [CrossRef] [PubMed]
- RIDASCREEN Cryptosporidium_Giardia-Combi. Available online: https://clinical.r-biopharm.com/wp-content/uploads/sites/3/2017/05/C1121-RIDASCREEN-Cryptosporidium_Giardia-Combi_lang-2017-04-20_EN.pdf (accessed on 11 June 2020).
- Aguirre, A.A.; Longcore, T.; Barbieri, M.; Dabritz, H.; Hill, D.; Klein, P.N.; Lepczyk, C.; Lilly, E.L.; McLeod, R.; Milcarsky, J.; et al. The one health approach to toxoplasmosis: Epidemiology, control, and prevention strategies. Ecohealth 2019, 16, 378–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, J.P.; Beattie, C.P. Toxoplasmosis of Animals and Man; CRC Press: Boca Raton, FL, USA, 1988; pp. 1–220. [Google Scholar]
- Bigal, E.; Morick, D.; Scheinin, A.P.; Salant, H.; Berkowitz, A.; King, R.; Levy, Y.; Melero, M.; Sánchez-Vizcaíno, J.M.; Goffman, O.; et al. Detection of Toxoplasma gondii in three common bottlenose dolphins (Tursiops truncatus); A first description from the Eastern Mediterranean Sea. Vet. Parasitol. 2018, 258, 74–78. [Google Scholar] [CrossRef]
- Di Guardo, G.; Proietto, U.; Di Francesco, C.E.; Marsilio, F.; Zaccaroni, A.; Scaravelli, D.; Mignone, W.; Garibaldi, F.; Kennedy, S.; Forster, F.; et al. Cerebral toxoplasmosis in striped dolphins (Stenella coeruleoalba) stranded along the Ligurian Sea coast of Italy. Vet. Pathol. 2010, 47, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Van Bressem, M.F.; Raga, J.A.; Di Guardo, G.; Jepson, P.D.; Duignan, P.J.; Siebert, U.; Barrett, T.; Santos, M.C.; Moreno, I.B.; Siciliano, S.; et al. Emerging infectious diseases in cetaceans worldwide and the possible role of environmental stressors. Dis. Aquat. Organ. 2009, 86, 143–157. [Google Scholar] [CrossRef]
- Massie, G.N.; Ware, M.W.; Villegas, E.N.; Black, M.W. Uptake and transmission of Toxoplasma gondii oocysts by migratory, filter-feeding fish. Vet. Parasitol. 2010, 169, 296–303. [Google Scholar] [CrossRef]
- Profeta, F.; Di Francesco, C.E.; Marsilio, F.; Mignone, W.; Di Nocera, F.; De Carlo, E.; Lucifora, G.; Pietroluongo, G.; Baffoni, M.; Cocumelli, C.; et al. Retrospective seroepidemiological investigations against Morbillivirus, Toxoplasma gondii and Brucella spp. in cetaceans stranded along the Italian coastline (1998–2014). Res. Vet. Sci. 2015, 101, 89–92. [Google Scholar] [CrossRef]
- Cabezon, O.; Resendes, A.R.; Domingo, M.; Raga, J.A.; Agusti, C.; Alegre, F.; Mons, J.L.; Dubey, J.P.; Almeria, S. Seroprevalence of Toxoplasma gondii antibodies in wild dolphins from the Spanish Mediterranean coast. J. Parasitol. 2004, 90, 643–644. [Google Scholar] [CrossRef]
- Van de Velde, N.; Devleesschauwer, B.; Leopold, M.; Begeman, L.; IJsseldijk, L.; Hiemstra, S.; IJzer, J.; Brownlow, A.; Davison, N.; Haelters, J.; et al. Toxoplasma gondii in stranded marine mammals from the North Sea and Eastern Atlantic Ocean: Findings and diagnostic difficulties. Vet. Parasitol. 2016, 230, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resendes, A.R.; Almería, S.; Dubey, J.P.; Obon, E.; Juan-Salles, C.; Degollada, E.; Alegre, F.; Cabezon, O.; Pont, S.; Domingo, M. Disseminated toxoplasmosis in a Mediterranean pregnant Risso’s dolphin (Grampus griseus) with transplacental fetal infection. J. Parasitol. 2002, 88, 1029–1032. [Google Scholar] [CrossRef]
- Dubey, J.P.; Zarnke, R.; Thomas, N.J.; Wong, S.K.; Van Bonn, W.; Briggs, M.; Davis, J.W.; Ewing, R.; Mense, M.; Kwok, O.C.H.; et al. Toxoplasma gondii, Neospora caninum, Sarcocystis neurona, and Sarcocystis canis-like infections in marine mammals. Vet. Parasitol. 2003, 116, 275–296. [Google Scholar] [CrossRef]
- Blanchet, M.A.; Godfroid, J.; Breines, E.M.; Heide-Jørgensen, M.P.; Nielsen, N.H.; Hasselmeier, J.; Iversen, M.; Jensen, S.K.; Åsbakk, K. West Greenland harbour porpoises assayed for antibodies against Toxoplasma gondii: False positives with the direct agglutination method. Dis. Aquat. Organ. 2014, 108, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Murata, K.; Mizuta, K.; Imazu, K.; Terasawa, F.; Taki, M.; Endoh, T. The prevalence of Toxoplasma gondii antibodies in wild and captive cetaceans from Japan. J. Parasitol. 2004, 90, 896–898. [Google Scholar] [CrossRef]
- Su, C.; Zhang, X.; Dubey, J.P. Genotyping of Toxoplasma gondii by multilocus PCR-RFLP markers: A high resolution and simple method for identification of parasites. Int. J. Parasitol. 2006, 36, 841–848. [Google Scholar] [CrossRef]
- Su, C.; Shwab, E.K.; Zhou, P.; Zhu, X.Q.; Dubey, J.P. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitology 2010, 137, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dubey, J.P.; Fair, P.A.; Sundar, N.; Velmurugan, G.; Kwok, O.C.; McFee, W.E.; Majumdar, D.; Su, C. Isolation of Toxoplasma gondii from bottlenose dolphins (Tursiops truncatus). J. Parasitol. 2008, 94, 821–823. [Google Scholar] [CrossRef]
- Di Guardo, G.; Di Cesare, A.; Otranto, D.; Casalone, C.; Iulini, B.; Mignone, W.; Tittarelli, C.; Meloni, S.; Castagna, G.; Forster, F.; et al. Genotyping of Toxoplasma gondii isolates in meningo-encephalitis affected striped dolphins (Stenella coeruleoalba) from Italy. Vet. Parasitol. 2011, 183, 31–36. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P. Long-term survival of Toxoplasma gondii sporulated oocysts in seawater. J. Parasitol. 2009, 95, 1019–1020. [Google Scholar] [CrossRef]
- Jones, J.L.; Dubey, J.P. Waterborne toxoplasmosis—Recent developments. Exp. Parasitol. 2009, 124, 10–25. [Google Scholar] [CrossRef]
- Marino, A.M.F.; Giunta, R.P.; Salvaggio, A.; Castello, A.; Alfonzetti, T.; Barbagallo, A.; Aparo, A.; Scalzo, F.; Reale, S.; Buffolano, W.; et al. Toxoplasma gondii in edible fishes captured in the Mediterranean basin. Zoonoses Public Health 2019, 66, 826–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Aguilar, R.; Delgado-Estrella, A.; Moreno-Navarrete, R.G. New host report for nematodes in a stranded short-snouted spinner dolphin Stenella clymene (Cetacea: Delphinidae) from the Mexican Caribbean coast. Helminthologia 2010, 47, 136–138. [Google Scholar] [CrossRef] [Green Version]
- Baylis, H.A.; Daubney, R. A revision of the lungworms of cetaceans. Parasitology 1925, 17, 201–216. [Google Scholar] [CrossRef]
- Dawes, B. The Trematoda, with Special Reference to British and Other European Forms; Cambridge University Press: Cambridge, UK, 1968; p. 364. [Google Scholar]
- Dailey, M.D. Parasites of marine mammals. In Marine Parasitology; Rohde, K., Ed.; CABI Publishing: Wallingford, UK, 2005; pp. 408–414. [Google Scholar]
- Kuwamura, M.; Sawamoto, O.; Yamate, J.; Aokii, M.; Ohnishi, Y.; Kotani, T. Pulmonary vascular proliferation and lungworm (Stenurus ovatus) in a bottlenose dolphin (Tursiops truncates). J. Vet. Med. Sci. 2007, 69, 531–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Cascante, L.; Gendron, D. Application of McMaster’s technique in live blue whales. Vet. Rec. 2012, 171, 220. [Google Scholar] [CrossRef] [PubMed]
- Grilo, M.L.; Gomes, L.; Wohlsein, P.; de Carvalho, L.M.; Siebert, U.; Lehnert, K. Cryptosporidium species and Giardia species prevalence in marine mammal species present in the North and Baltic Seas. J. Zoo. Wildl. Med. 2018, 49, 1002–1006. [Google Scholar] [CrossRef]
- Cacciò, S.M.; De Giacomo, M.; Pozio, E. Sequence analysis of the β-giardin gene and development of a polymerase chain reaction-restriction fragment length polymorphism assay to genotype Giardia duodenalis cysts from human faecal samples. Int. J. Parasitol. 2002, 32, 1023–1030. [Google Scholar] [CrossRef]
- Pedraza-Diaz, S.; Amar, C.; Nichols, G.L.; McLauchin, J. Nested polymerase chain reaction for amplication of the Cryptosporidium oocyst wall protein gene. Emerg. Infect. Dis. 2001, 7, 49–56. [Google Scholar] [CrossRef]
- Measures, L.N.; Dubey, J.P.; Labelle, P.; Martineau, D. Seroprevalence of Toxoplasma gondii in Canadian Pinnipeds. J. Wild. Dis. 2004, 40, 294–300. [Google Scholar] [CrossRef] [Green Version]
- Dubey, J.P.; Morales, N.; Sundar, G.; Velmurugan, V.; Gonzalez-Barrientos, C.R.; Hernandez-Mora, G.; Su, C. Isolation and genetic characterization of Toxoplasma gondii from striped dolphins (Stenella coeruleoalba). J. Parasitol. 2007, 93, 710–711. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Mergi, J.; Gehring, E.; Sundar, N.; Velmurugan, G.V.; Kwok, O.C.H.; Grigg, M.E.; Su, C.; Martineau, D. Toxoplasmosis in captive dolphins (Tursiops truncatus) and Walrus (Odobenus rosmarus). J. Parasitol. 2009, 95, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Alba, P.; Terracciano, G.; Franco, A.; Lorenzetti, S.; Cocumelli, C.; Fichi, G.; Eleni, C.; Zygmunt, M.S.; Cloeckaert, A.; Battisti, A. The presence of Brucella ceti ST26 in a Striped Dolphin (Stenella coeruleoalba) with Meningoencephalitis from the Mediterranean Sea. Vet. Microbiol. 2013, 164, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Glantz, S.A. Statistica per Discipline Biomediche, 5th ed.; McGraw-Hill Companies Srl.: Milan, Italy, 2003. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 26 June 2020).




{kind=link}
{kind=link}
{kind=link}
| Parasites | Positive | Negative | Prevalence | IC95 % |
|---|---|---|---|---|
| Striped dolphin (Stenella coeruleoalba, n = 15) | ||||
| Nematodes | ||||
| Halocercus lagenorhynchi | 1 | 14 | 6.67% | 0.00–19.29 |
| Halocercus delphini | 2 | 13 | 13.33% | 0.00–30.54 |
| Crassicauda spp. | 1 | 14 | 6.67% | 0.00–19.29 |
| Trematodes | ||||
| Pholeter gastrophilus | 4 | 11 | 26.67% | 4.29–49.05 |
| Campula palliata | 1 | 14 | 6.67% | 0.00–19.29 |
| Cestodes | ||||
| Phyllobothrium delphini | 11 | 4 | 73.33% | 50.95–95.71 |
| Monorygma grimaldii | 5 | 10 | 33.33% | 9.48–57.19 |
| Tetrabothrium forsteri | 2 | 13 | 13.33% | 0.00–30.54 |
| Strobilocephalus triangularis | 2 | 13 | 13.33% | 0.00–30.54 |
| Protozoa (serology) | ||||
| Toxoplasma gondii | 5 | 10 | 33.33% | 9.48–57.19 |
| Bottlenose dolphin (Tursiops truncatus, n = 10) | ||||
| Nematodes | ||||
| Skrjabinalius guevarai | 2 | 8 | 20.00% | 0.00–44.79 |
| Stenurus ovatus | 2 | 8 | 20.00% | 0.00–44.79 |
| Crassicauda spp. | 1 | 9 | 10.00% | 0.00–28.59 |
| Trematodes | ||||
| Pholeter gastrophilus | 3 | 7 | 30.00% | 1.60–58.40 |
| Acanthocephalans | ||||
| Bolbosoma vasculosum | 2 | 8 | 20.00% | 0.00–44.79 |
| Protozoa (serology) | ||||
| Toxoplasma gondii | 1 | 9 | 10.00% | 0.00–28.59 |
| Genetic Markers | ||||||
|---|---|---|---|---|---|---|
| SAG1 * | 5′ +3′ SAG2 ** | alt SAG2 *** | SAG3 | BTUB | GRA6 | |
| Genotype | II/III | II | II | II | II | II |
| Species | Gender | Weight (kg) | Year of Stranding | Geographic Localities (Geographic Coordinates) |
|---|---|---|---|---|
| Stenella coeruleoalba | M | 49 | 2013 | Tirrenia (Pisa) 43°37′38″ N–10°17′28″ E |
| Stenella coeruleoalba | M | 100 | 2013 | Monte Argentario (Grosseto) 42°26′07″ N–11°07′00″ E |
| Stenella coeruleoalba | M | - | 2013 | Orbetello (Grosseto) 42°26′22″ N–11°12′45″ E |
| Stenella coeruleoalba | F | 45 | 2013 | Follonica (Grosseto) 42°55′08″ N–10°45′41″ E |
| Stenella coeruleoalba | F | 61 | 2013 | Calambrone (Pisa) 43°35′49″ N–10°17′40″ E |
| Stenella coeruleoalba | M | 65 | 2013 | Piombino (Livorno) 42°56′05.37″ N–10°31′19.63″ E |
| Stenella coeruleoalba | F | 25 | 2013 | Bibbona (Livorno) 16′12.64″ N–10°35′54.73″ E |
| Stenella coeruleoalba | M | 50 | 2013 | Livorno 43°32′36″ N–10°19′1″ E |
| Stenella coeruleoalba | M | 6 | 2013 | Livorno 43°32’36” N–10°19′1″ E |
| Stenella coeruleoalba | M | 75 | 2013 | Marina di Grosseto (Grosseto) 42°42′55″ N–10°59′02″ E |
| Stenella coeruleoalba | F | 60 | 2013 | Orbetello (Grosseto) 42°26′22″ N–11°12′45″ E |
| Stenella coeruleoalba | M | 75 | 2014 | Cecina (Livorno) 43°18′42.53″ N–10°31′08.49″ E |
| Stenella coeruleoalba | M | - | 2014 | Piombino (Livorno) 42°56′05.37″ N–10°31′19.63″ E |
| Stenella coeruleoalba | M | 29.5 | 2015 | Portoferraio (Livorno) 42°48′45″ N–10°18′56″ E |
| Stenella coeruleoalba | M | 45 | 2015 | Marina di Massa (Massa) 44°00′33.8″ N–10°06′06.9″ E |
| Tursiops truncatus | M | - | 2013 | Marina di Castagneto Carducci (Livorno) 43°10′39.93″ N–10°32′20.27″ E |
| Tursiops truncatus | * | - | 2013 | Calambrone (Pisa) 43°35′49″N–10°17′40″E |
| Tursiops truncatus | M | 140 | 2013 | Calambrone (Pisa) 43°35′49″N–10°17′40″E |
| Tursiops truncatus | F | 70 | 2013 | Rosignano Marittimo (Livorno) 43°24′32″ N–10°28′26″ E |
| Tursiops truncatus | F | 27 | 2014 | Livorno 43°32′36″ N–10°19′1″ E |
| Tursiops truncatus | M | 300 | 2014 | Marina di Pietrasanta (Lucca) 43°55′35.1″ N–10°11′48.3″ E |
| Tursiops truncatus | M | 30 | 2014 | Tirrenia (Pisa) 43°37′38″ N–10°17′28″ E |
| Tursiops truncatus | M | 140 | 2014 | Rosignano Marittimo (Livorno) 43°24′32″ N–10°28′26″ E |
| Tursiops truncatus | F | 102 | 2015 | Marina di Pisa (Pisa) 43°40′20″ N–10°16′37″ E |
| Tursiops truncatus | F | 43 | 2015 | Livorno 43°32′36″ N–10°19′1″ E |
| Grampus griseus | F | - | 2015 | San Vincenzo (Livorno) 43°06′02.28″ N–10°32′11.96″ E |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terracciano, G.; Fichi, G.; Comentale, A.; Ricci, E.; Mancusi, C.; Perrucci, S. Dolphins Stranded along the Tuscan Coastline (Central Italy) of the “Pelagos Sanctuary”: A Parasitological Investigation. Pathogens 2020, 9, 612. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080612
Terracciano G, Fichi G, Comentale A, Ricci E, Mancusi C, Perrucci S. Dolphins Stranded along the Tuscan Coastline (Central Italy) of the “Pelagos Sanctuary”: A Parasitological Investigation. Pathogens. 2020; 9(8):612. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080612
Chicago/Turabian StyleTerracciano, Giuliana, Gianluca Fichi, Antonia Comentale, Enrica Ricci, Cecilia Mancusi, and Stefania Perrucci. 2020. "Dolphins Stranded along the Tuscan Coastline (Central Italy) of the “Pelagos Sanctuary”: A Parasitological Investigation" Pathogens 9, no. 8: 612. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9080612