
Enhanced Immunogenicity of Inactivated Dengue Vaccines by Novel Polysaccharide-Based Adjuvants in Mice

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
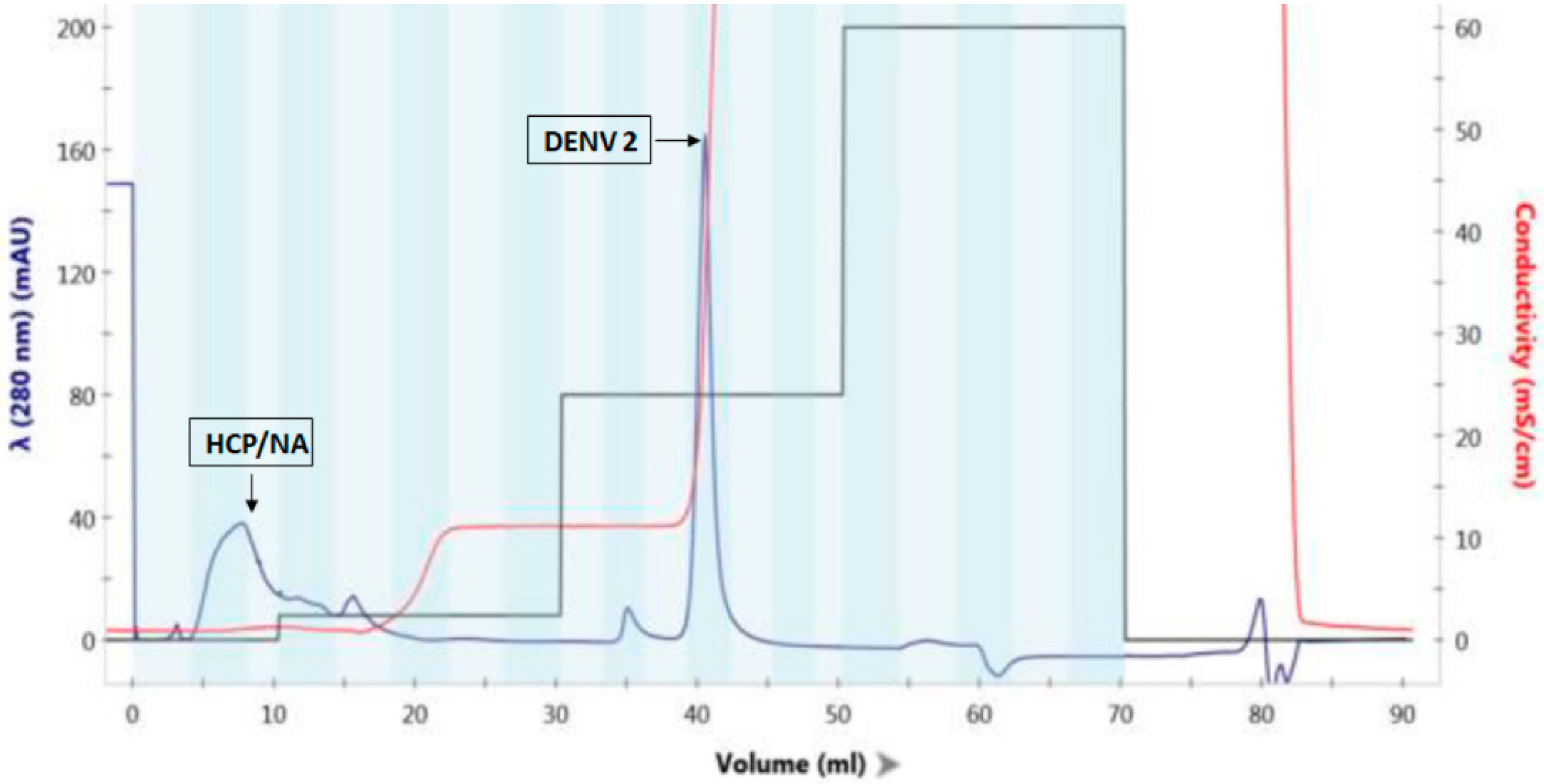
2.1. Chemical Inactivation of DENV2 Particles
2.1.1. DENV2 Inactivation Using Formalin
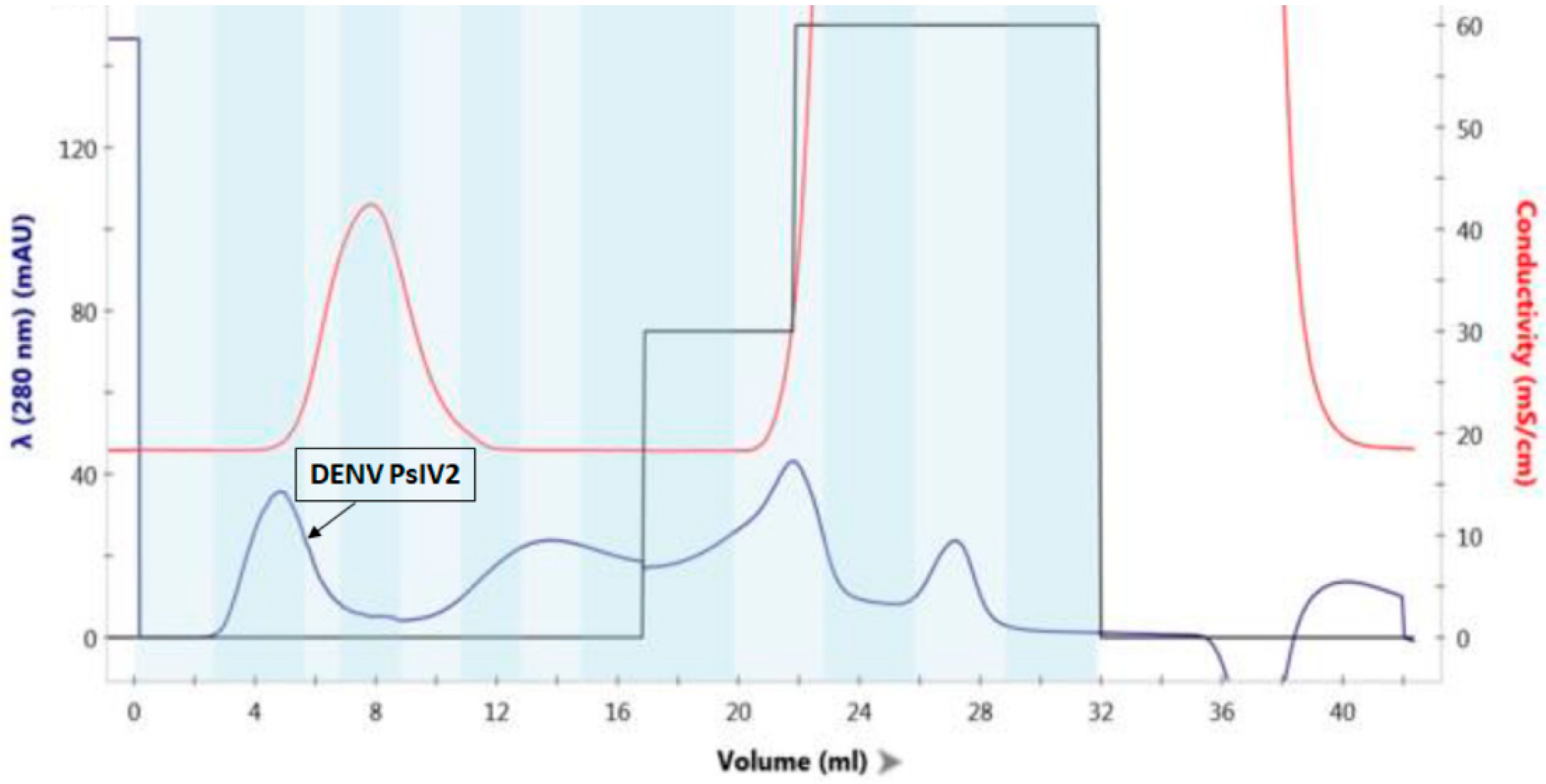
2.1.2. DENV2 Inactivation Using Psoralen
2.2. Quantitation of DENV2 PsIV and PIV
2.3. Preparation of Vaccine-Adjuvant Formulations
2.4. Animals and Vaccine Administration
2.5. Microneutralization Tests to Measure Neutralizing Antibody Responses
2.6. ELISPOT Assay to Measure T-Cell IFNγ and IL4 Responses
2.7. ELISA to Measure the Mouse Immunoglobulin Isotypes
3. Results
3.1. Characterization of DENV2 PsIV Vaccine Antigen
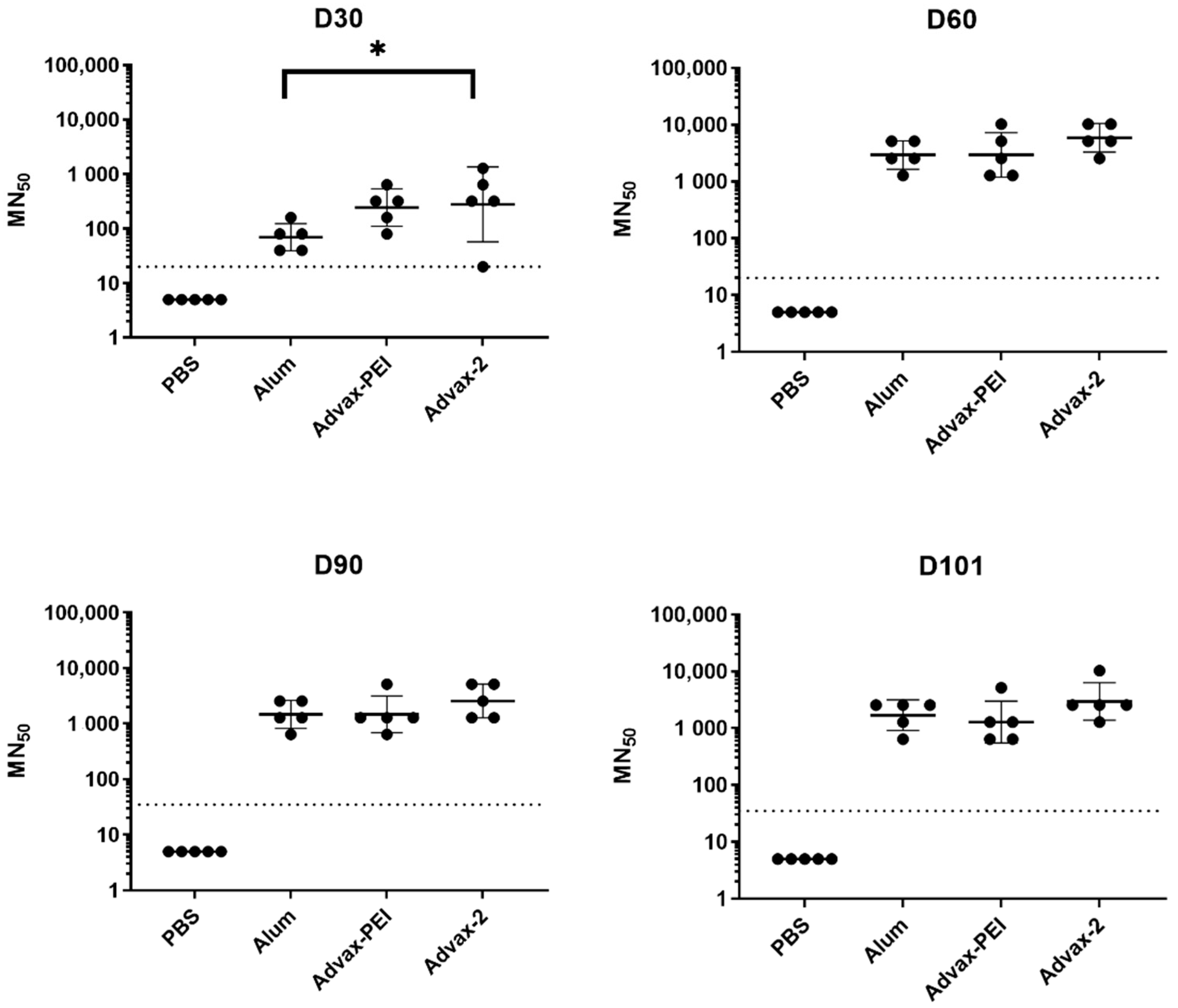
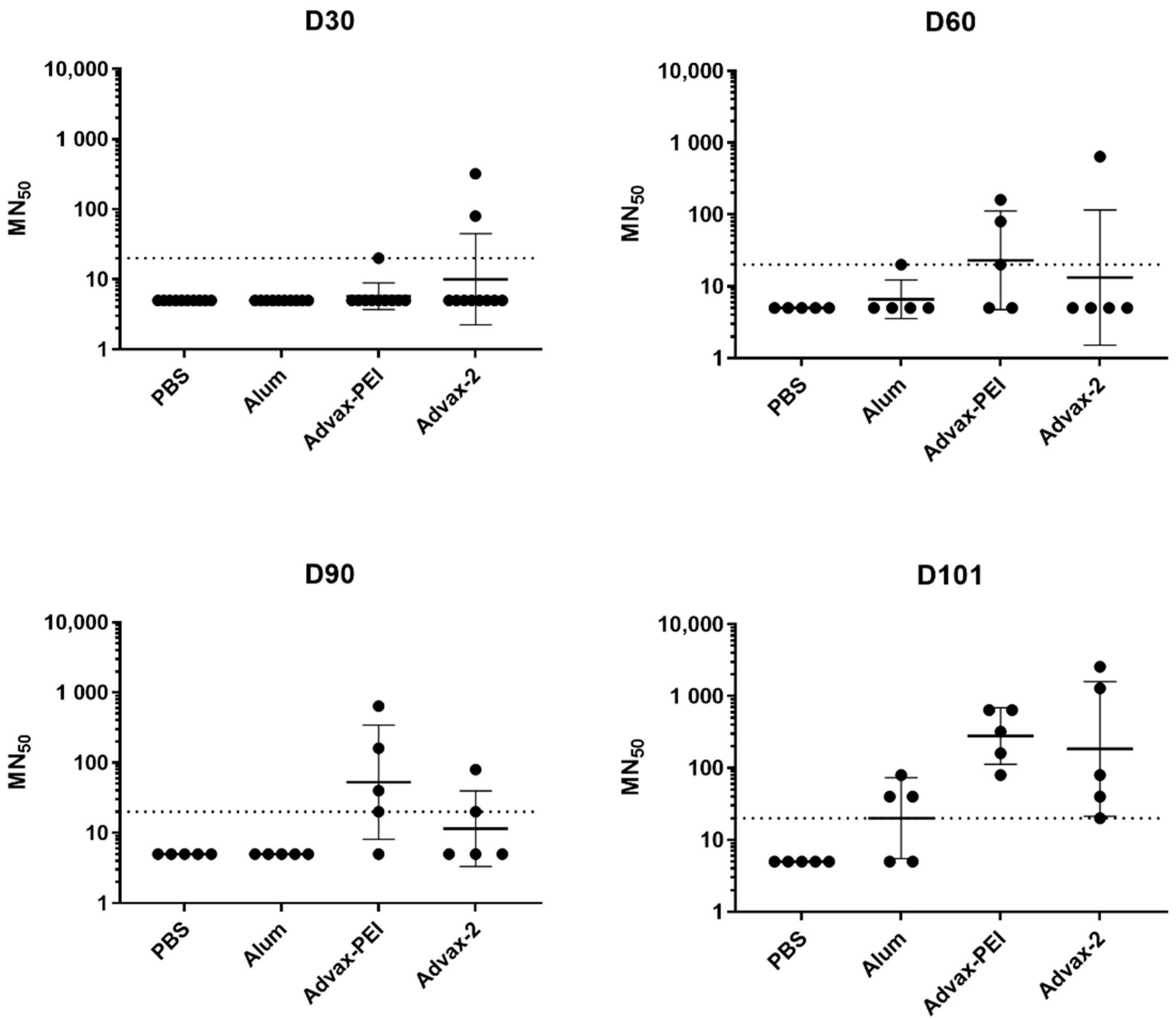
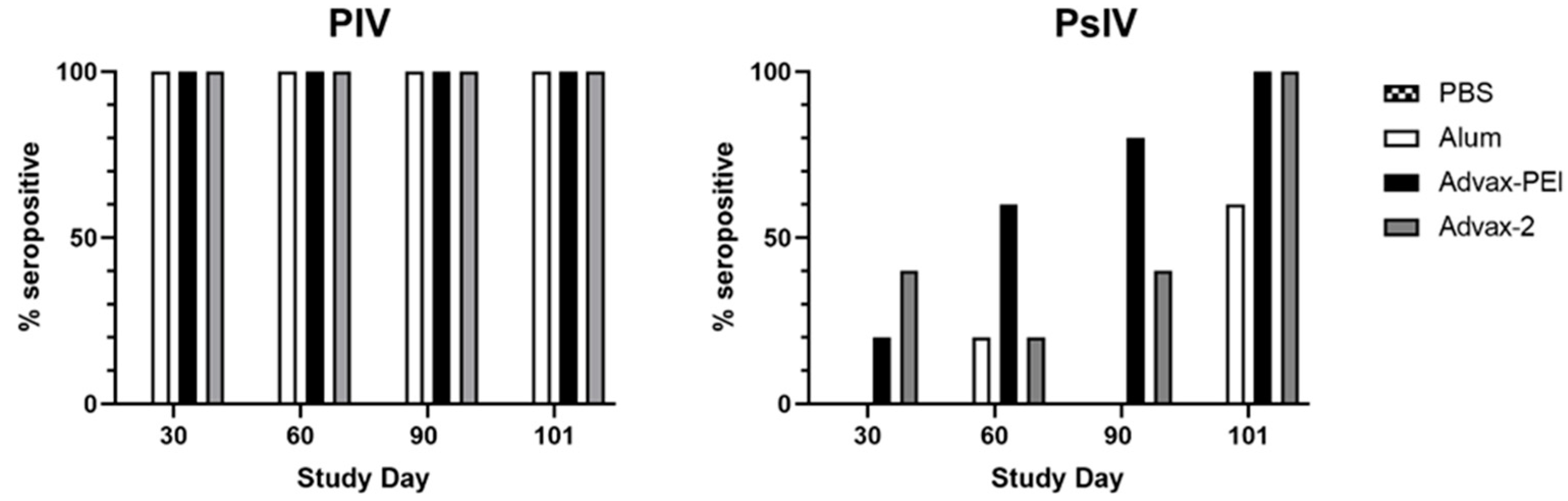
3.2. Neutralizing Antibody Responses
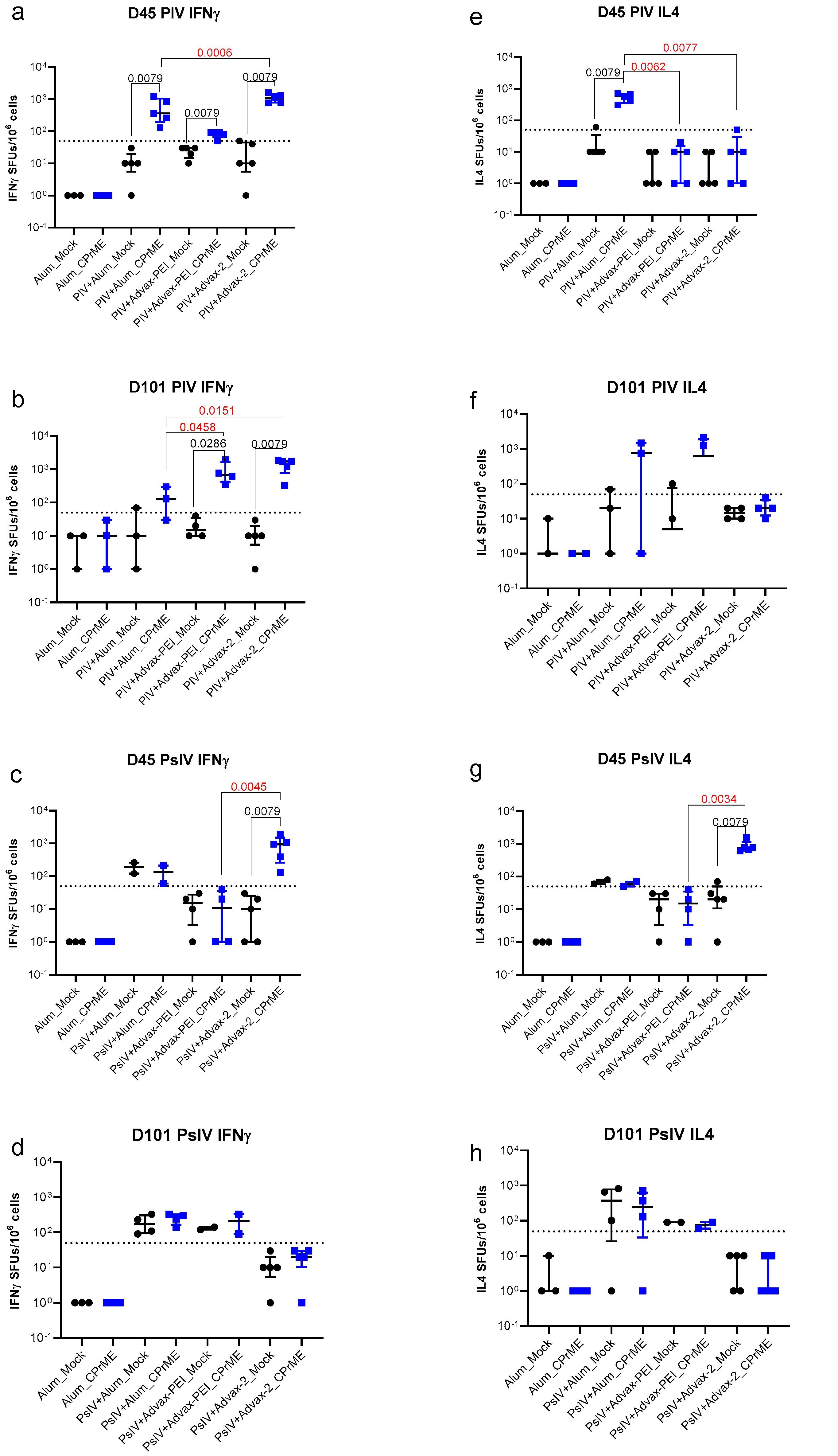
3.3. ELISPOT Assays for Measurement of Cell-Mediated Immunogenicity
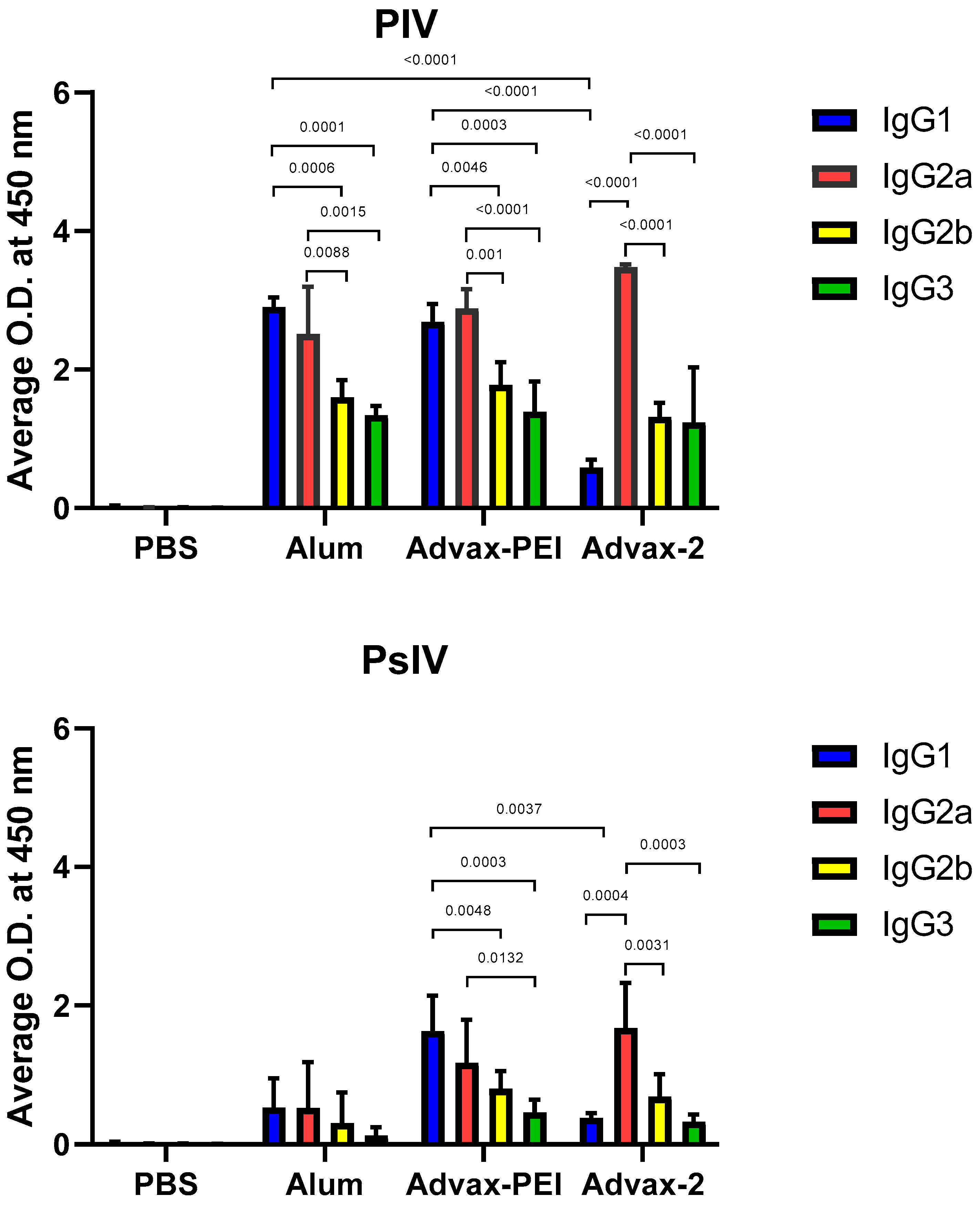
3.4. Immunoglobulin G Isotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Burnette, W.N.; Hoke, C.H.; Scovill, J.; Clark, K.; Abrams, J.; Kitchen, L.W.; Hanson, K.; Palys, T.J.; Vaughn, D.W. Infectious Diseases Investment Decision Evaluation Algorithm: A Quantitative Algorithm for Prioritization of Naturally Occurring Infectious Disease Threats to the U.S. Military. Mil. Med. 2008, 173, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Bozza, F.A.; Cruz, O.G.; Zagne, S.M.; Azeredo, E.L.; Nogueira, R.M.; Assis, E.F.; Bozza, P.T.; Kubelka, C.F. Multiplex cytokine profile from dengue patients: MIP-1beta and IFN-gamma as predictive factors for severity. BMC Infect. Dis. 2008, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Raviprakash, K.; Defang, G.; Burgess, T.; Porter, K. Advances in dengue vaccine development. Hum. Vaccines 2009, 5, 520–528. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.J.; Endy, T.P. Critical issues in dengue vaccine development. Curr. Opin. Infect. Dis. 2011, 24, 442–450. [Google Scholar] [CrossRef]
- WHO Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 14 April 2022).
- Domingo, E.; Baranowski, E.; Ruiz-Jarabo, C.M.; Martin-Hernandez, A.M.; Saiz, J.C.; Escarmis, C. Quasispecies structure and persistence of RNA viruses. Emerg. Infect. Dis. 1998, 4, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Putnak, R.; Cassidy, K.; Conforti, N.; Lee, R.; Sollazzo, D.; Truong, T.; Ing, E.; Dubois, D.; Sparkuhl, J.; Gastle, W.; et al. Immunogenic and protective response in mice immunized with a purified, inactivated, Dengue-2 virus vaccine prototype made in fetal rhesus lung cells. Am. J. Trop. Med. Hyg. 1996, 55, 504–510. [Google Scholar] [CrossRef]
- Fernandez, S.; Thomas, S.J.; De La Barrera, R.; Im-Erbsin, R.; Jarman, R.G.; Baras, B.; Toussaint, J.F.; Mossman, S.; Innis, B.L.; Schmidt, A.; et al. An adjuvanted, tetravalent dengue virus purified inactivated vaccine candidate induces long-lasting and protective antibody responses against dengue challenge in rhesus macaques. Am. J. Trop. Med. Hyg. 2015, 92, 698–708. [Google Scholar] [CrossRef]
- Martinez, L.J.; Lin, L.; Blaylock, J.M.; Lyons, A.G.; Bauer, K.M.; De La Barrera, R.; Simmons, M.; Jarman, R.G.; Currier, J.R.; Friberg, H.; et al. Safety and Immunogenicity of a Dengue Virus Serotype-1 Purified-Inactivated Vaccine: Results of a Phase 1 Clinical Trial. Am. J. Trop. Med. Hyg. 2015, 93, 454–460. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.C.; Lin, L.; Martinez, L.J.; Ruck, R.C.; Eckels, K.H.; Collard, A.; De La Barrera, R.; Paolino, K.M.; Toussaint, J.-F.; Lepine, E.; et al. Phase 1 Randomized Study of a Tetravalent Dengue Purified Inactivated Vaccine in Healthy Adults in the United States. Am. J. Trop. Med. Hyg. 2017, 96, 1325–1337. [Google Scholar] [CrossRef] [Green Version]
- Diaz, C.; Lin, L.; Martinez, L.J.; Eckels, K.H.; Campos, M.; Jarman, R.G.; De La Barrera, R.; Lepine, E.; Toussaint, J.-F.; Febo, I.; et al. Phase I Randomized Study of a Tetravalent Dengue Purified Inactivated Vaccine in Healthy Adults from Puerto Rico. Am. J. Trop. Med. Hyg. 2018, 98, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Raviprakash, K.; Sun, P.; Raviv, Y.; Luke, T.; Martin, N.; Kochel, T. Dengue virus photo-inactivated in presence of 1,5-iodonaphthylazide (INA) or AMT, a psoralen compound (4’-aminomethyl-trioxsalen) is highly immunogenic in mice. Hum. Vaccines Immunother. 2013, 9, 2336–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaram, A.K.; Ewing, D.; Blevins, M.; Liang, Z.; Sink, S.; Lassan, J.; Raviprakash, K.; Defang, G.; Williams, M.; Porter, K.R.; et al. Comparison of purified psoralen-inactivated and formalin-inactivated dengue vaccines in mice and nonhuman primates. Vaccine 2020, 38, 3313–3320. [Google Scholar] [CrossRef] [PubMed]
- Alving, C.R.; Rao, M.; Steers, N.J.; Matyas, G.R.; Mayorov, A.V. Liposomes containing lipid A: An effective, safe, generic adjuvant system for synthetic vaccines. Expert Rev. Vaccines 2012, 11, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Kelley, P.; Heinzel, S.; Cooper, P.; Petrovsky, N. Immunogenicity and safety of Advax™, a novel polysaccharide adjuvant based on delta inulin, when formulated with hepatitis B surface antigen: A randomized controlled Phase 1 study. Vaccine 2014, 32, 6469–6477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, D.L.; Sajkov, D.; Honda-Okubo, Y.; Wilks, S.H.; Aban, M.; Barr, I.G.; Petrovsky, N. Human Phase 1 trial of low-dose inactivated seasonal influenza vaccine formulated with Advax delta inulin adjuvant. Vaccine 2016, 34, 3780–3786. [Google Scholar] [CrossRef]
- Heddle, R.; Smith, A.; Woodman, R.; Hissaria, P.; Petrovsky, N. Randomized controlled trial demonstrating the benefits of delta inulin adjuvanted immunotherapy in patients with bee venom allergy. J. Allergy Clin. Immunol. 2019, 144, 504–513.e16. [Google Scholar] [CrossRef]
- Honda-Okubo, Y.; Barnard, D.; Ong, C.H.; Peng, B.-H.; Tseng, C.-T.K.; Petrovsky, N. Severe Acute Respiratory Syndrome-Associated Coronavirus Vaccines Formulated with Delta Inulin Adjuvants Provide Enhanced Protection while Ameliorating Lung Eosinophilic Immunopathology. J. Virol. 2015, 89, 2995–3007. [Google Scholar] [CrossRef] [Green Version]
- Komiya, T.; Honda-Okubo, Y.; Baldwin, J.; Petrovsky, N. An Advax-Adjuvanted Inactivated Cell-Culture Derived Japanese Encephalitis Vaccine Induces Broadly Neutralising Anti-Flavivirus Antibodies, Robust Cellular Immunity and Provides Single Dose Protection. Vaccines 2021, 9, 1235. [Google Scholar] [CrossRef]
- Fernandez, S.; Cisney, E.D.; Tikhonov, A.P.; Schweitzer, B.; Putnak, R.J.; Simmons, M.; Ulrich, R.G. Antibody Recognition of the Dengue Virus Proteome and Implications for Development of Vaccines. Clin. Vaccine Immunol. 2011, 18, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Simmons, M.; Sun, P.; Putnak, R. Recombinant Dengue 2 Virus NS3 Helicase Protein Enhances Antibody and T-Cell Response of Purified Inactivated Vaccine. PLoS ONE 2016, 11, e0152811. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.; Ewing, D.; Blevins, M.; Sun, P.; Sundaram, A.K.; Raviprakash, K.S.; Porter, K.R.; Sanders, J.W. Enhanced immunogenicity and protective efficacy of a tetravalent dengue DNA vaccine using electroporation and intradermal delivery. Vaccine 2019, 37, 4444–4453. [Google Scholar] [CrossRef] [PubMed]
- Danko, J.R.; Kochel, T.; Teneza-Mora, N.; Luke, T.C.; Raviprakash, K.; Sun, P.; Simmons, M.; Moon, J.E.; De La Barrera, R.; Martinez, L.J.; et al. Safety and Immunogenicity of a Tetravalent Dengue DNA Vaccine Administered with a Cationic Lipid-Based Adjuvant in a Phase 1 Clinical Trial. Am. J. Trop. Med. Hyg. 2018, 98, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.; Triccas, A.J.; Petrovsky, N. Adjuvant Strategies for More Effective Tuberculosis Vaccine Immunity. Microorganisms 2019, 7, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountford, A.P.; Fisher, A.; Wilson, R.A. The profile of IgG1 and IgG2a antibody responses in mice exposed to Schistosoma mansoni. Parasite Immunol. 1994, 16, 521–527. [Google Scholar] [CrossRef]
- Petrovsky, N.; Cooper, P.D. Advax, a novel microcrystalline polysaccharide particle engineered from delta inulin, provides robust adjuvant potency together with tolerability and safety. Vaccine 2015, 33, 5920–5926. [Google Scholar] [CrossRef]
- Honda-Okubo, Y.; Saade, F.; Petrovsky, N. Advax™, a polysaccharide adjuvant derived from delta inulin, provides improved influenza vaccine protection through broad-based enhancement of adaptive immune responses. Vaccine 2012, 30, 5373–5381. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; Aoshi, T.; Haseda, Y.; Kobiyama, K.; Wijaya, E.; Nakatsu, N.; Igarashi, Y.; Standley, D.M.; Yamada, H.; Honda-Okubo, Y.; et al. Advax, a Delta Inulin Microparticle, Potentiates In-built Adjuvant Property of Co-administered Vaccines. eBioMedicine 2016, 15, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Monslow, M.A.; Freed, D.C.; Chang, D.; Li, F.; Gindy, M.; Wang, D.; Vora, K.; Espeseth, A.S.; Petrovsky, N.; et al. Novel adjuvants enhance immune responses elicited by a replication-defective human cytomegalovirus vaccine in nonhuman primates. Vaccine 2021, 39, 7446–7456. [Google Scholar] [CrossRef]
- Li, L.; Honda-Okubo, Y.; Li, C.; Sajkov, D.; Petrovsky, N. Delta Inulin Adjuvant Enhances Plasmablast Generation, Expression of Activation-Induced Cytidine Deaminase and B-Cell Affinity Maturation in Human Subjects Receiving Seasonal Influenza Vaccine. PLoS ONE 2015, 10, e0132003. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Vaccine-Adjuvant Formation | Number of Mice |
|---|---|---|
| 1 | PIVD2 + Alum | 10 |
| 2 | PsIVD2 + Alum | 10 |
| 3 | PIVD2 + Advax-PEI | 10 |
| 4 | PIVD2 + Adxax-2 | 10 |
| 5 | PsIVD2 + Advax-PEI | 10 |
| 6 | PsIVD2 + Adxax-2 | 10 |
| 7 | Alum | 10 |
| Procedure | Day of Study | |||||
|---|---|---|---|---|---|---|
| 0 | 30 | 45 | 60 | 90 | 101 | |
| Vaccination (n = 10) | X | X | X | |||
| Bleeding (n = 10) | X | X | X | X | X | |
| Spleen harvest (n = 5) | X | X | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.-J.; Ewing, D.; Sundaram, A.K.; Chen, H.-W.; Liang, Z.; Cheng, Y.; Jani, V.; Sun, P.; Gromowski, G.D.; De La Barrera, R.A.; et al. Enhanced Immunogenicity of Inactivated Dengue Vaccines by Novel Polysaccharide-Based Adjuvants in Mice. Microorganisms 2022, 10, 1034. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051034
Wu S-J, Ewing D, Sundaram AK, Chen H-W, Liang Z, Cheng Y, Jani V, Sun P, Gromowski GD, De La Barrera RA, et al. Enhanced Immunogenicity of Inactivated Dengue Vaccines by Novel Polysaccharide-Based Adjuvants in Mice. Microorganisms. 2022; 10(5):1034. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051034
Chicago/Turabian StyleWu, Shuenn-Jue, Dan Ewing, Appavu K. Sundaram, Hua-Wei Chen, Zhaodong Liang, Ying Cheng, Vihasi Jani, Peifang Sun, Gregory D. Gromowski, Rafael A. De La Barrera, and et al. 2022. "Enhanced Immunogenicity of Inactivated Dengue Vaccines by Novel Polysaccharide-Based Adjuvants in Mice" Microorganisms 10, no. 5: 1034. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051034