
First Case Report of Detection of Multidrug-Resistant Enterobacter hormaechei in Clinical Sample from an Aborted Ruminant


Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens from a Cow
2.2. Determination of AMR Phenotype
2.3. DNA Extraction and Sequencing
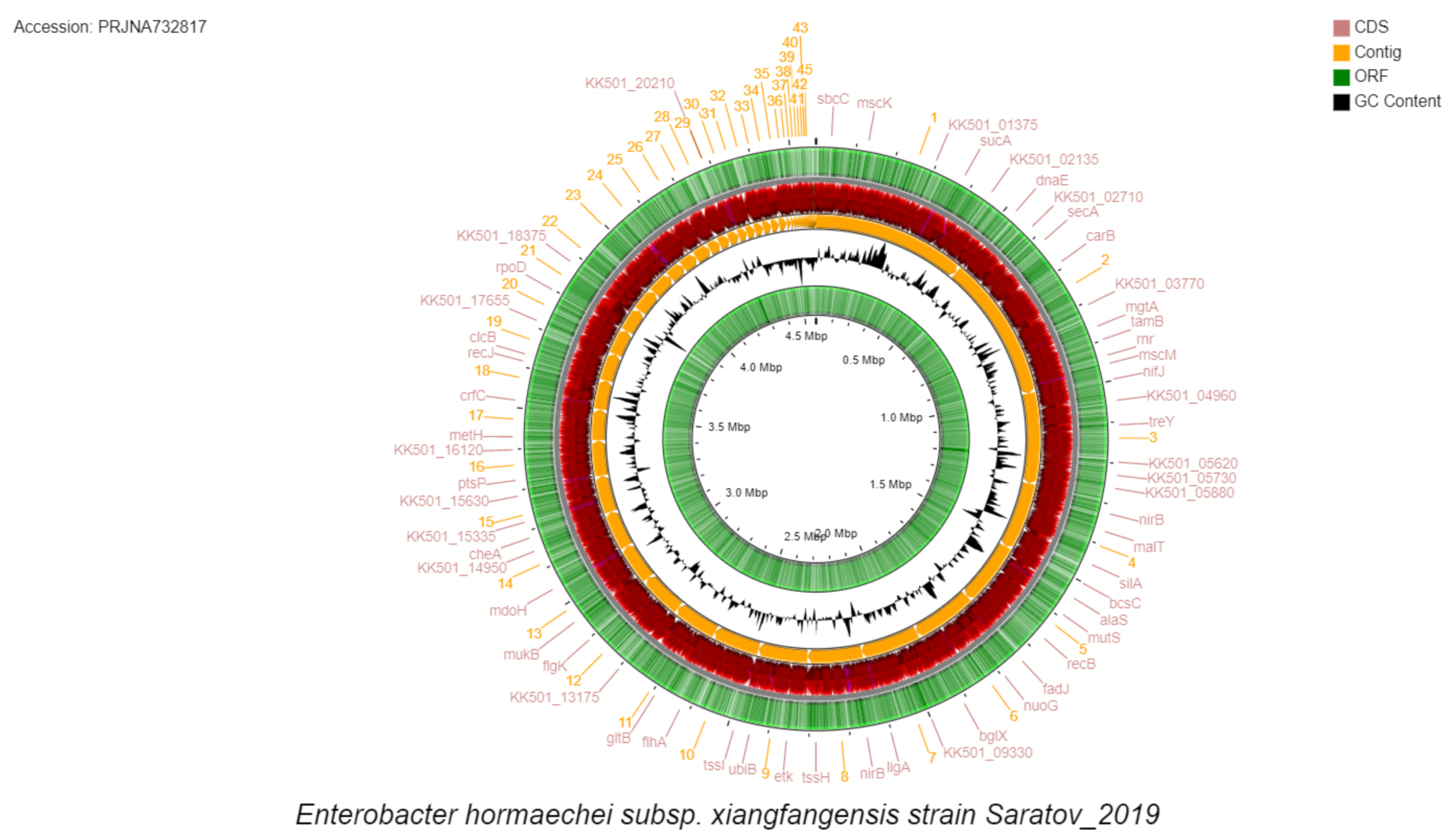
2.4. Bioinformatic Data Processing
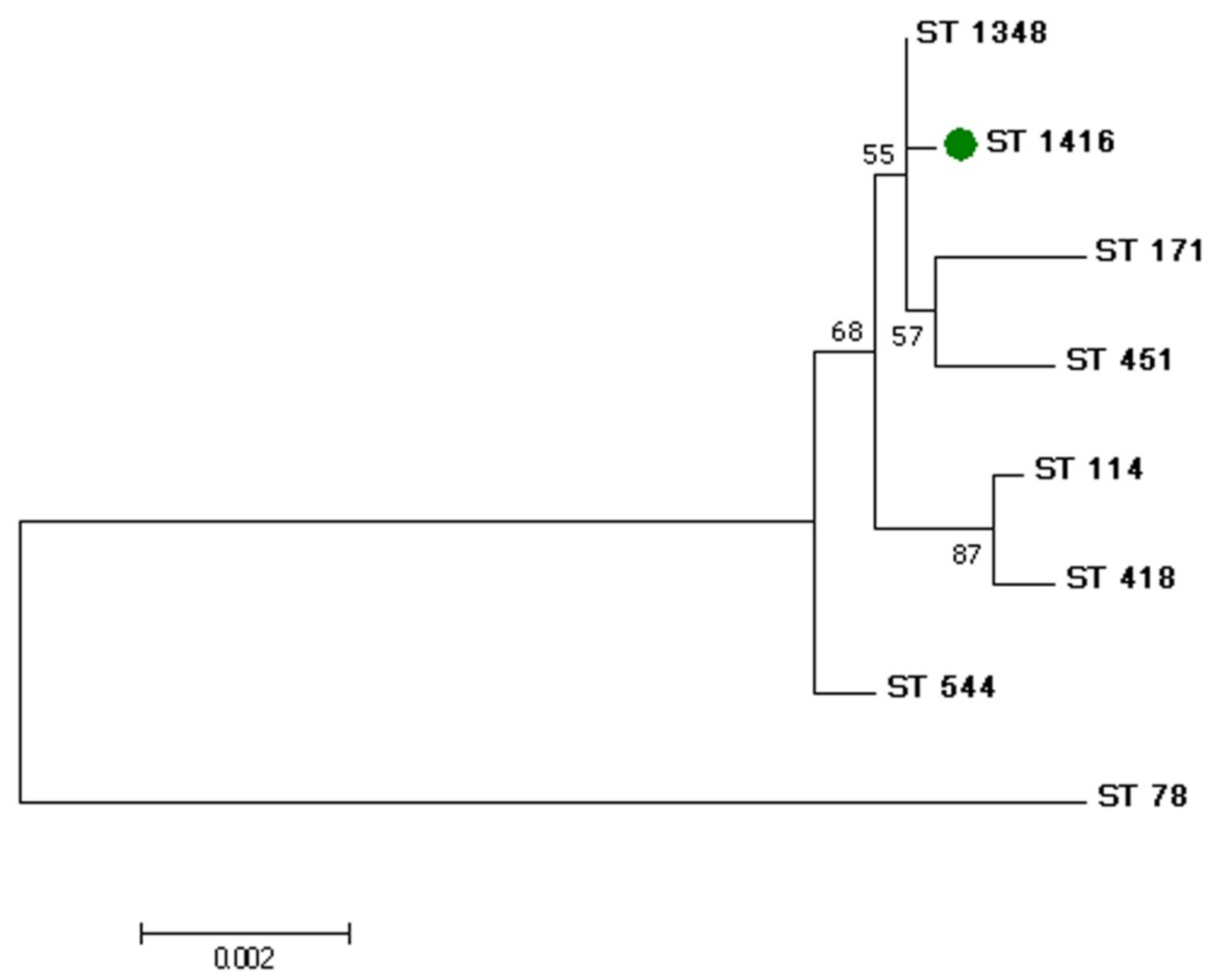
2.5. MLST-Typing
2.6. Determination of the AMR Genotype
2.7. List of Genomes Included in This Study
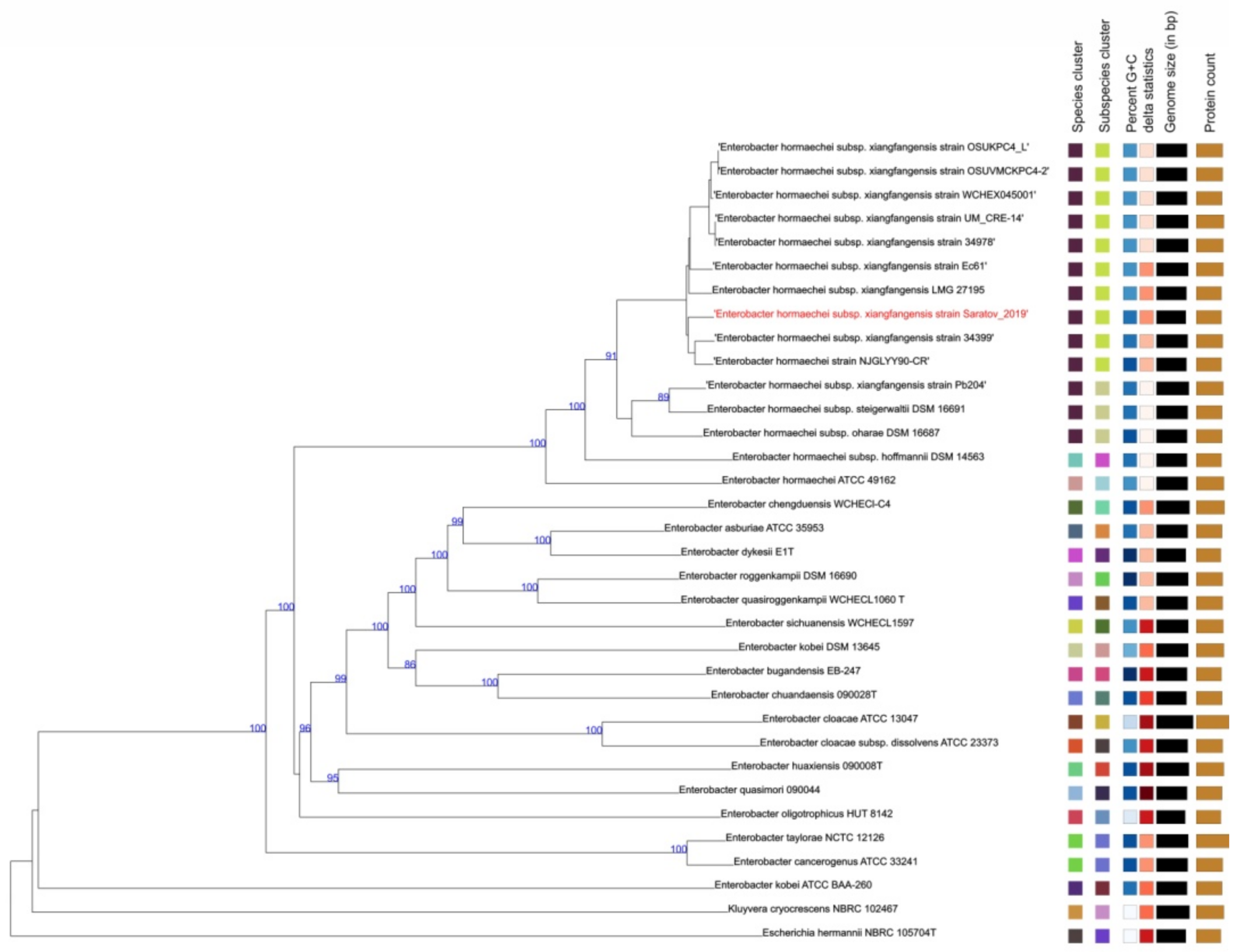
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia, J.; Rodrigues, F.; Castro, F.; Aires, A.; Marques, G.; Saavedra, M.J. Antimicrobial, Antibiofilm, and Antioxidant Properties of Boletus edulis and Neoboletus luridiformis Against Multidrug-Resistant ESKAPE Pathogens. Front. Nutr. 2021, 8, 773346. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, P.; Singh, S. Multidrug Resistance in Bacteria: A Serious Patient Safety Challenge for India. J. Lab. Physicians 2013, 5, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. Science 2017, 355, 826–830. [Google Scholar] [CrossRef]
- Medina, E.; Pieper, D.H. Tackling Threats and Future Problems of Multidrug-Resistant Bacteria. Curr. Top. Microbiol. Immunol. 2016, 398, 3–33. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Liao, W.; Cui, Y.; Quan, J.; Zhao, D.; Han, X.; Shi, Q.; Wang, Q.; Jiang, Y.; Du, X.; Li, X.; et al. High prevalence of colistin resistance and mcr-9/10 genes in Enterobacter spp. in a tertiary hospital over a decade. Int. J. Antimicrob. Agents 2022, 59, 106573. [Google Scholar] [CrossRef]
- Brenner, D.J.; McWhorter, A.C.; Kai, A.; Steigerwalt, A.G.; Farmer, J.J. Enterobacter asburiae sp. nov., a new species found in clinical specimens, and reassignment of Erwinia dissolvens and Erwinia nimipressuralis to the genus Enterobacter as Enterobacter dissolvens comb. nov. and Enterobacter nimipressuralis comb. nov. J. Clin. Microbiol. 1986, 23, 1114–1120. [Google Scholar] [CrossRef] [Green Version]
- Davin-Regli, A.; Lavigne, J.-P.; Pagès, J.-M. Enterobacter spp.: Update on Taxonomy, Clinical Aspects, and Emerging Antimicrobial Resistance. Clin. Microbiol. Rev. 2019, 32, e00002-19. [Google Scholar] [CrossRef]
- Villa, J.; Carretero, O.; Viedma, E.; Lora-Tamayo, J.; Mingorance, J.; Chaves, F. Emergence of NDM-7-producing multi-drug-resistant Enterobacter hormaechei sequence type ST-78 in Spain: A high-risk international clone. Int. J. Antimicrob. Agents 2018, 53, 533–534. [Google Scholar] [CrossRef]
- Wu, L.; Xu, T.; Ji, Y.; Song, J.; Wang, F.; Huang, J.; Zhou, K. Occurrence and Characteristics of Mcr s among Gram-Negative Bacteria Causing Bloodstream Infections of Infant Inpatients between 2006 and 2019 in China. Microbiol. Spectr. 2022, 10, e0193821. [Google Scholar] [CrossRef] [PubMed]
- Beyrouthy, R.; Barets, M.; Marion, E.; Dananché, C.; Dauwalder, O.; Robin, F.; Gauthier, L.; Jousset, A.; Dortet, L.; Guérin, F.; et al. NovelEnterobacterLineage as Leading Cause of Nosocomial Outbreak Involving Carbapenemase-Producing Strains. Emerg. Infect. Dis. 2018, 24, 1505–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- (Biohaz), E.P.O.B.H.; Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; et al. Role played by the environment in the emergence and spread of antimicrobial resistance (AMR) through the food chain. EFSA J. 2021, 19, e06651. [Google Scholar] [CrossRef]
- EMA Committee for Medicinal Products for Veterinary Use (CVMP); EFSA Panel on Biological Hazards (BIOHAZ); Murphy, D.; Ricci, A.; Auce, Z.; Beechinor, J.G.; Bergendahl, H.; Breathnach, R.; Bures, J.; Duarte Da Silva, J.P.; et al. EMA and EFSA Joint Scientific Opinion on measures to reduce the need to use antimicrobial agents in animal husbandry in the European Union, and the resulting impacts on food safety (RONAFA). EFSA J. 2017, 15, e04666. [Google Scholar] [CrossRef]
- De Almeida, C.C.; Financi, T.P.; Cardozo, M.V.; Pizauro, L.J.L.; Pereira, N.; Valmorbida, M.K.; Borzi, M.M.; Weiss, B.; De Ávila, F.A. Enterobacteriaceae in calves, cows and milking environment may act as reservoirs of virulence and antimicrobial resistance genes. Food Sci. Technol. 2020, 41, 376–380. [Google Scholar] [CrossRef]
- Shan-Shan, W.; Yun-Jia, S.; Xing-Yang, C.; Cheng-Wei, W.; Shan-Shan, G.U.; Xin, Y.; Shuang, X.; Jun-Wei, G.E.; Hong-Yan, C. Isolation, identification and phylogenetic analysis of Enterobacter hormaechei from foxes. Chin. Vet. Sci. 2017, 47, 768–772. [Google Scholar]
- Lu-Yao, L.; Liu, M.; Teng, M.; Wang, L.; Zhang, Y.; Liu, B.J.J.A.S.V.M. Study on the biological characteristics of Enterobacter hormaechei. J. Anim. Sci. Vet. Med. 2017, 36, 1–6. [Google Scholar]
- Wang, Z.; Duan, L.; Liu, F.; Hu, Y.; Leng, C.; Kan, Y.; Yao, L.; Shi, H. First report of Enterobacter hormaechei with respiratory disease in calves. BMC Vet. Res. 2020, 16, 1–4. [Google Scholar] [CrossRef]
- Harada, K.; Shimizu, T.; Mukai, Y.; Kuwajima, K.; Sato, T.; Kajino, A.; Usui, M.; Tamura, Y.; Kimura, Y.; Miyamoto, T.; et al. Phenotypic and molecular characterization of antimicrobial resistance in Enterobacter spp. isolates from companion animals in Japan. PLoS ONE 2017, 12, e0174178. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 23rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013; pp. 29–177. ISBN 1-56238-866-5. [Google Scholar]
- Mathew, A.; Joseph, I. Characterization of functionally diverse intestinal bacterial flora of Panulirus homarus (Linnaeus, 1758) along the southwest coast of India. J. Mar. Biol. Assoc. India 2019, 61, 38–43. [Google Scholar] [CrossRef]
- Akbari, M.; Bakhshi, B.; Peerayeh, S.N. Particular Distribution of Enterobacter cloacae Strains Isolated from Urinary Tract Infection within Clonal Complexes. Iran. Biomed. J. 2015, 20, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Daniels, J.B.; Chen, L.; Grooters, S.V.; Mollenkopf, D.F.; Mathys, D.A.; Pancholi, P.; Kreiswirth, B.N.; Wittum, T.E. Enterobacter cloacae Complex Sequence Type 171 Isolates Expressing KPC-4 Carbapenemase Recovered from Canine Patients in Ohio. Antimicrob. Agents Chemother. 2018, 62, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavda, K.D.; Chen, L.; Fouts, D.E.; Sutton, G.; Brinkac, L.; Jenkins, S.G.; Bonomo, R.A.; Adams, M.D.; Kreiswirth, B.N. Comprehensive Genome Analysis of Carbapenemase-Producing Enterobacter spp.: New Insights into Phylogeny, Population Structure, and Resistance Mechanisms. mBio 2016, 7, e02093-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Zhang, K.; Chen, W.; Chen, J.; Zheng, J.; Liu, C.; Cheng, L.; Zhou, W.; Shen, H.; Cao, X.; et al. Epidemiological characteristics of carbapenem-resistant Enterobacteriaceae collected from 17 hospitals in Nanjing district of China. Antimicrob. Resist. Infect. Control 2020, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, W.M.B.S.; Martins, E.R.; de Andrade, L.K.; Farzana, R.; Walsh, T.R.; Toleman, M.A.; Nogueira, M.C.L.; Gales, A.C. BKC-2, a New BKC Variant Detected in MCR-9.1-Producing Enterobacter hormaechei subsp. xiangfangensis. Antimicrob. Agents Chemother. 2021, 65, 1–6. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ben Khedher, M.; Baron, S.A.; Riziki, T.; Ruimy, R.; Raoult, D.; Diene, S.M.; Rolain, J.-M. Massive analysis of 64,628 bacterial genomes to decipher water reservoir and origin of mobile colistin resistance genes: Is there another role for these enzymes? Sci. Rep. 2020, 10, 5970. [Google Scholar] [CrossRef] [Green Version]
- Lascols, C.; Peirano, G.; Hackel, M.; Laupland, K.B.; Pitout, J.D.D. Surveillance and Molecular Epidemiology of Klebsiella pneumoniae Isolates That Produce Carbapenemases: First Report of OXA-48-Like Enzymes in North America. Antimicrob. Agents Chemother. 2013, 57, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Lutgring, J.D.; Zhu, W.; de Man, T.J.; Avillan, J.J.; Anderson, K.F.; Lonsway, D.R.; Rowe, L.A.; Batra, D.; Rasheed, J.K.; Limbago, B.M. Phenotypic and Genotypic Characterization of Enterobacteriaceae Producing Oxacillinase-48–Like Carbapenemases, United States. Emerg. Infect. Dis. 2018, 24, 700–709. [Google Scholar] [CrossRef] [Green Version]
- Chavda, K.D.; Westblade, L.F.; Satlin, M.J.; Hemmert, A.C.; Castanheira, M.; Jenkins, S.G.; Chen, L.; Kreiswirth, B.N. First Report of bla VIM-4 and mcr-9 -Coharboring Enterobacter Species Isolated from a Pediatric Patient. mSphere 2019, 4, e00629-19. [Google Scholar] [CrossRef] [Green Version]
- Ai, W.; Zhou, Y.; Wang, B.; Zhan, Q.; Hu, L.; Xu, Y.; Guo, Y.; Wang, L.; Yu, F.; Li, X. First Report of Coexistence of blaSFO–1 and blaNDM–1 β-Lactamase Genes as Well as Colistin Resistance Gene mcr-9 in a Transferrable Plasmid of a Clinical Isolate of Enterobacter hormaechei. Front. Microbiol. 2021, 12, 676113. [Google Scholar] [CrossRef] [PubMed]
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and Other Enterobacteriaceae: An Evolving Crisis of Global Dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The Role of Epidemic Resistance Plasmids and International High-Risk Clones in the Spread of Multidrug-Resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirano, G.; Matsumura, Y.; Adams, M.D.; Bradford, P.; Motyl, M.; Chen, L.; Kreiswirth, B.N.; Pitout, J.D. Genomic Epidemiology of Global Carbapenemase-Producing Enterobacter spp., 2008–2014. Emerg. Infect. Dis. 2018, 24, 1010–1019. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Naas, T.; Poirel, L. Global Spread of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain ID | ST | Allele | Reference/Source | ||||||
|---|---|---|---|---|---|---|---|---|---|
| dnaA | fusA | gyrB | leuS | pyrG | rplB | rpoB | |||
| E. hormaechei subsp. xiangfangensis strain Saratov_2019 | 1416 | 46 | 21 | 19 | 44 | 13 a | 4 | 6 | This study |
| Nameless b | 1348 | 46 | 21 | 19 | 44 | 45 | 4 | 6 | https://pubmlst.org (accessed on 15 May 2022) |
| E. hormaechei subsp. xiangfangensis LMG27195 | 544 | 10 | 21 | 9 | 44 | 45 | 4 | 33 | [22] |
| E. hormaechei subsp. xiangfangensis strain 34399 | 114 | 53 | 35 | 20 | 44 | 45 | 4 | 6 | [23] |
| E. hormaechei subsp. xiangfangensis strain 34978 | 171 | 49 | 21 | 19 | 44 | 45 | 12 | 32 | [24] |
| E. hormaechei subsp. xiangfangensis strain OSUKPC4_L | 171 | 49 | 21 | 19 | 44 | 45 | 12 | 32 | GenBank accession number: CP024908.1 |
| E. hormaechei subsp. xiangfangensis strain OSUVMCKPC4-2 | 171 | 49 | 21 | 19 | 44 | 45 | 12 | 32 | GenBank accession number: CP029246.1 |
| E. hormaechei subsp. xiangfangensis strain UM_CRE-14 | 171 | 49 | 21 | 19 | 44 | 45 | 12 | 32 | https://pubmlst.org (accessed on 15 May 2022) |
| E. hormaechei subsp. xiangfangensis strain WCHEX045001 | 171 | 49 | 21 | 19 | 44 | 45 | 12 | 32 | https://pubmlst.org (accessed on 15 May 2022) |
| E. hormaechei subsp. xiangfangensis strain Ec61 | 451 | 146 | 21 | 148 | 44 | 99 | 4 | 6 | [25] |
| E. hormaechei strain NJGLYY90-CR | 418 | 53 | 35 | 154 | 44 | 45 | 4 | 6 | [26] |
| E. hormaechei strain 1801 c | 78 | 8 | 9 | 6 | 9 | 9 | 6 | 8 | https://pubmlst.org (accessed on 15 May 2022) |
| No. | Predicted with CARD | Identified in the Strain * | Confirmation by the DDT ** | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ARO Term a | AMR Gene Family | Drug Group | Gene | Product | Locus_Tag in the Contig | Contig No. | Drug Group b | Sensitive | Resistant | |
| 1 | Escherichia coli ampH beta-lactamase | ampC-type beta-lactamase | C | ampH | D-alanyl-D-alaninecarboxypeptidase/endopeptidase AmpH | KK501_00085 | 1 | C (third-generation) | - | + |
| 2 | emrR | major facilitator superfamily (MFS) antibiotic efflux pump | F | emrR | multidrug efflux transporter EmrAB transcriptional repressor EmrR | KK501_07160 | 5 | |||
| 3 | rsmA | resistance-nodulation-cell division (RND) antibiotic efflux pump | F, D | rsmA | 16S rRNA (adenine(1518)-N(6)/adenine(1519)-N(6))- dimethyltransferase RsmA | KK501_03125 | 2 | |||
| 4 | adeF | resistance-nodulation-cell division (RND) antibiotic efflux pump | F, T | oqxB | multidrug efflux RND transporter permease subunit OqxB | KK501_09445 | 7 | |||
| 5 | oqxA | resistance-nodulation-cell division (RND) antibiotic efflux pump | F, G, T, D, Nf | oqxB | multidrug efflux RND transporter permease subunit OqxB | KK501_09445 | 7 | T | - | + |
| 6 | Klebsiella pneumoniae kpnE | major facilitator superfamily (MFS) antibiotic efflux pump | M, Ag, T, P, R | mdtJ | multidrug/spermidine efflux SMR transporter subunit MdtJ | KK501_17290 | 19 | First-generation Ag M | - | + + |
| 7 | Klebsiella pneumoniae kpnF | major facilitator superfamily (MFS) antibiotic efflux pump | M, Ag, C, T, P, R | mdtJ | multidrug/spermidine efflux SMR transporter subunit MdtJ | KK501_17290 | 19 | |||
| 8 | baeR | resistance-nodulation-cell division (RND) antibiotic efflux pump | Ag, Ac | baeR | two-component system response regulator BaeR | KK501_18050 | 21 | |||
| 9 | acrD | resistance-nodulation-cell division (RND) antibiotic efflux pump | Ag | acrD | multidrug efflux RND transporter permease AcrD | KK501_09635 | 7 | |||
| 10 | msbA | ATP-binding cassette (ABC) antibiotic efflux pump | Nm | msbA | lipid A ABC transporter ATP-binding protein/permease MsbA | KK501_13920 | 13 | Nm | - | + |
| 11 | acrR | AcrA/B complex | O | acrR | multidrug efflux transporter transcriptional repressor AcrR | KK501_00590 | 1 | O | - | + |
| 12 | fosA2 | fosfomycin thiol transferase | Ps | fosA | FosA/FosA2 family fosfomycin resistance glutathione transferase | KK501_03840 | 2 | Ps | - | + |
| 13 | Escherichia coli uhpT | antibiotic-resistant UhpT | Ps | uhpT | hexose-6-phosphate: phosphate antiporter | KK501_18665 | 22 | Ps | - | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaitsev, S.S.; Khizhnyakova, M.A.; Feodorova, V.A. First Case Report of Detection of Multidrug-Resistant Enterobacter hormaechei in Clinical Sample from an Aborted Ruminant. Microorganisms 2022, 10, 1036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051036
Zaitsev SS, Khizhnyakova MA, Feodorova VA. First Case Report of Detection of Multidrug-Resistant Enterobacter hormaechei in Clinical Sample from an Aborted Ruminant. Microorganisms. 2022; 10(5):1036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051036
Chicago/Turabian StyleZaitsev, Sergey S., Mariya A. Khizhnyakova, and Valentina A. Feodorova. 2022. "First Case Report of Detection of Multidrug-Resistant Enterobacter hormaechei in Clinical Sample from an Aborted Ruminant" Microorganisms 10, no. 5: 1036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051036