
Elevated Seawater Temperature and Infection with Neoparamoeba perurans Exacerbate Complex Gill Disease in Farmed Atlantic Salmon (Salmo salar) in British Columbia, Canada


Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Collection and Gill Scoring
2.2. Histology
2.3. Neoparamoeba Perurans Quantitative PCR (qPCR)
2.4. Statistical Analysis
3. Results
3.1. Temperature and Salinity
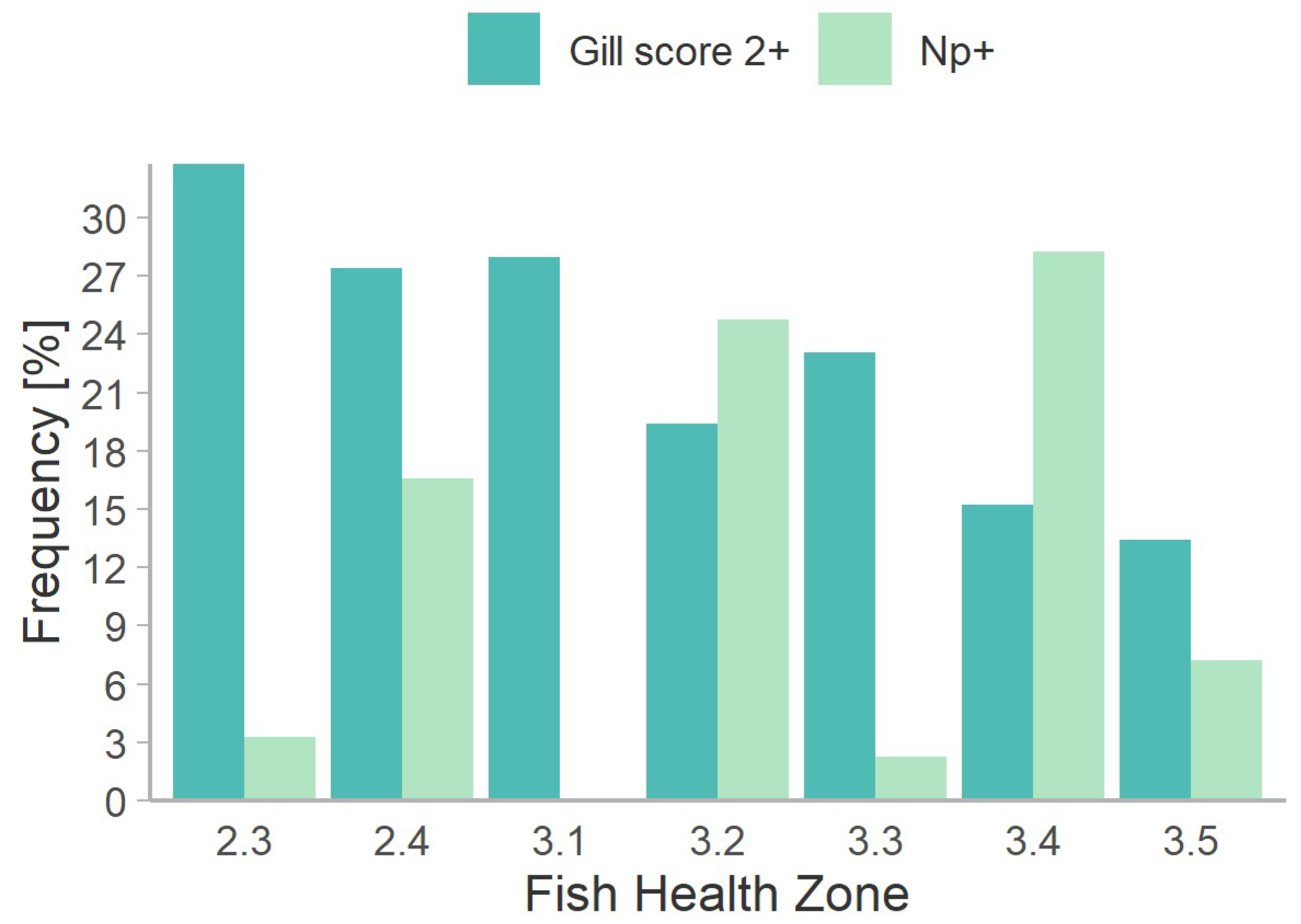
3.2. Neoparamoeba perurans in Fish Health Audits and Individual Fish
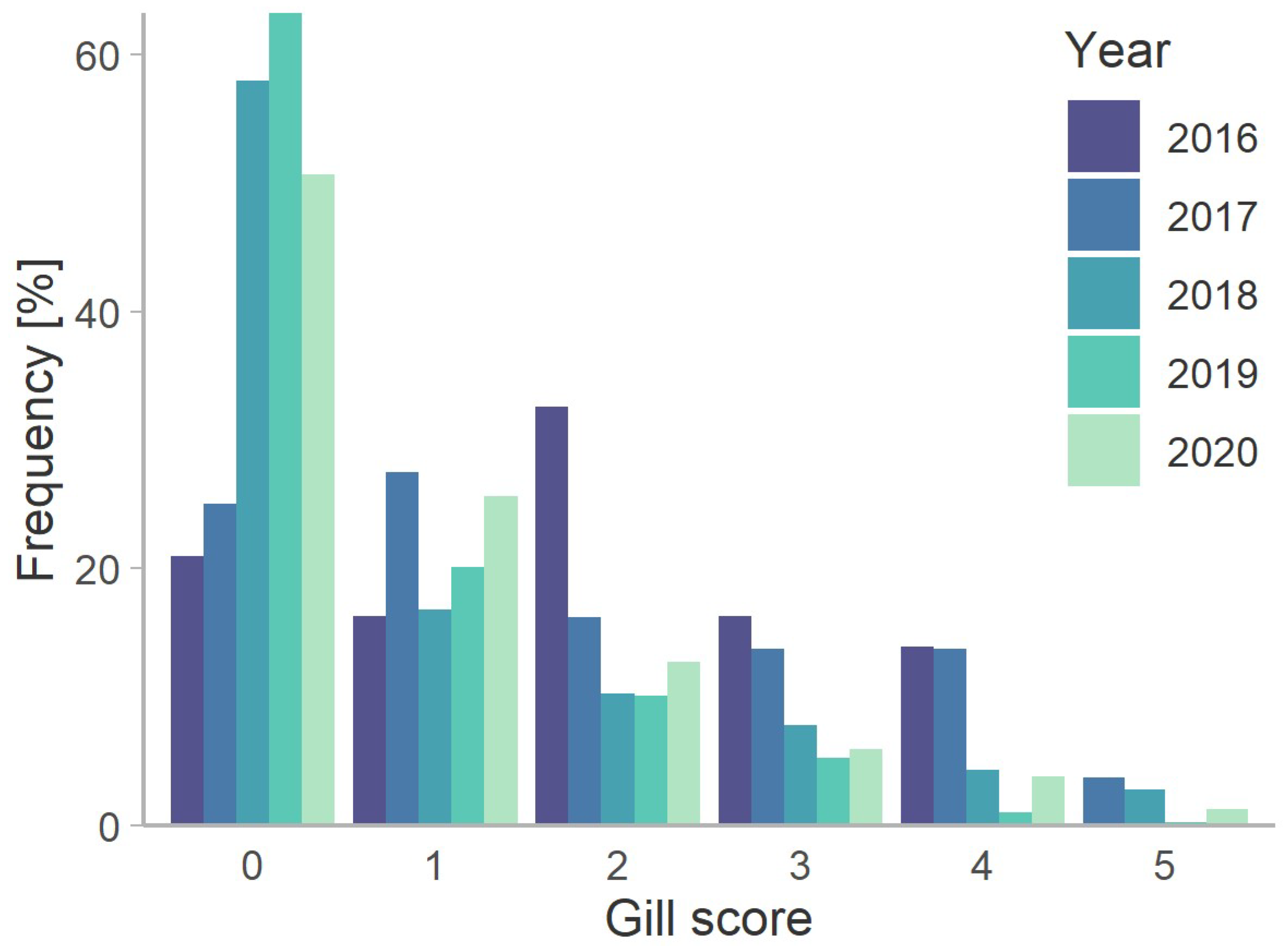
3.3. Amoebic Gill Disease and Gill Damage
3.4. Days at Sea
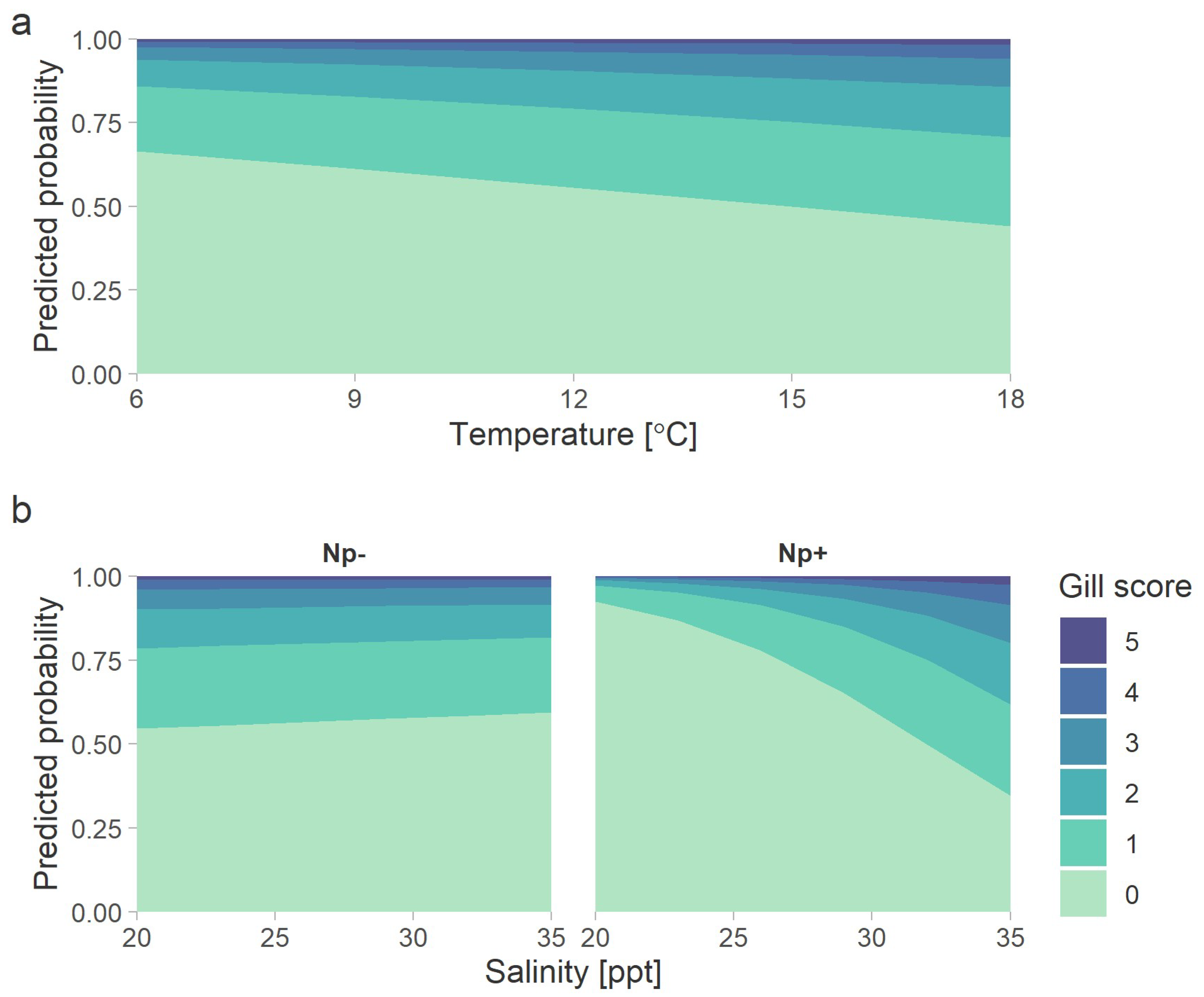
3.5. Logistic Model
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boerlage, A.S.; Ashby, A.; Herrero, A.; Reevers, A.; Gunn, G.J.; Rodger, H.D. Epidemiology of marine gill diseases in Atlantic salmon (Salmo salar) aquaculture: A review. Rev. Aquacult. 2020, 12, 2140–2159. [Google Scholar] [CrossRef]
- Young, N.D.; Crosbie, P.; Adams, M.; Nowak, B.; Morrison, R. Neoparamoeba perurans n. sp., an agent of amoebic gill disease of Atlantic salmon (Salmo salar). Int. J. Parasitol. 2007, 37, 1469–1481. [Google Scholar] [CrossRef] [PubMed]
- Crosbie, P.B.; Bridle, A.; Cadoret, K.; Nowak, B. In Vitro cultured Neoparamoeba perurans causes amoebic gill disease in Atlantic salmon and fulfills Koch’s postulates. Int. J. Parasitol. 2012, 42, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Oldham, T.; Rodger, H.; Nowak, B.F. Incidence and distribution of amoebic gill disease (AGD)—An epidemiological review. Aquaculture 2016, 457, 35–42. [Google Scholar] [CrossRef]
- Nowak, B.F. Neoparamoeba perurans. In Fish Parasites, Pathobiology and Protection; Woo, P.T.K., Buchmann, K., Eds.; CAB International: Surrey, UK, 2012; pp. 1–18. [Google Scholar]
- Adams, M.; Crosbie, P.; Nowak, B. Preliminary success using hydrogen peroxide to treat Atlantic salmon, Salmo salar L., affected with experimentally induced amoebic gill disease (AGD). J. Fish Dis. 2012, 35, 389–848. [Google Scholar] [CrossRef]
- Munday, B.; Zilberg, D.; Findlay, V. Gill disease of marine fish caused by infection with Neoparamoeba pemaquidensis. J. Fish Dis. 2001, 24, 497–507. [Google Scholar] [CrossRef]
- Nowak, B.F.; Archibald, J.M. Opportunistic but lethal: The mystery of paramoebae. Trends Parasitol. 2018, 34, 404–419. [Google Scholar] [CrossRef]
- Clark, A.; Nowak, B.F. Field investigations of amoebic gill disease in Atlantic salmon, Salmo salar L., in Tasmania. J. Fish Dis. 1999, 22, 433–443. [Google Scholar] [CrossRef]
- Adams, M.B.; Ellard, K.; Nowak, B.F. Gross pathology and its relationship with histopathology of amoebic gill disease (AGD) in farmed Atlantic salmon Salmo salar L. J. Fish Dis. 2004, 27, 151–161. [Google Scholar] [CrossRef]
- Bustos, P.A.; Young, N.D.; Rozas, M.A.; Bohle, H.M.; Ildefonso, R.S.; Morrison, R.N.; Nowak, B.F. Amoebic gill disease (AGD) in Atlantic salmon (Salmo salar) farmed in Chile. Aquaculture 2011, 310, 281–288. [Google Scholar] [CrossRef]
- Rodger, H.D.; McArdle, J.F. An outbreak of amoebic gill disease in Ireland. Vet. Record 1996, 139, 348–349. [Google Scholar] [CrossRef]
- Steinum, T.; Kvellestad, A.; Rønneberg, L.; Nilsen, H.; Asheim, A.; Fjell, K.; Nygård, S.; Olsen, A.; Dale, O. First cases of amoebic gill disease (AGD) in Norwegian seawater farmed Atlantic salmon Salmo salar L., and phylogeny of the causative amoeba using 18S cDNA sequences. J. Fish Dis. 2008, 31, 205–214. [Google Scholar] [CrossRef]
- Mouton, A.; Crosbie, P.; Cadoret, K.; Nowak, B. First record of amoebic gill disease caused by Neoparamoeba perurans in South Africa. J. Fish Dis. 2013, 37, 407–409. [Google Scholar] [CrossRef]
- Stagg, H.E.B.; Hall, M.; Wallace, I.S.; Pert, C.C.; Garcia Perez, S.; Collins, C. Detection of Paramoeba perurans in Scottish marine wild fish populations. Bull. Eur. Assoc. Fish Pathol. 2015, 35, 217–226. [Google Scholar]
- Kent, M.L.; Sawyer, T.; Hedrick, R. Paramoeba pemaquidensis (Sarcomastigophora: Paramoebidae) infestation of the gills of coho salmon Oncorhynchus kisutch reared in sea water. Dis. Aquat. Org. 1988, 5, 163–169. [Google Scholar] [CrossRef]
- Nowak, B.F.; Bryan, J.; Jones, S. Do salmon lice, Lepeophtheirus salmonis, have a role in the epidemiology of amoebic gill disease caused by Neoparamoeba perurans? J. Fish Dis. 2010, 33, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Douglas-Helders, M.; Saksida, S.; Raverty, S.; Nowak, B.F. Temperature as a risk factor for outbreaks of amoebic gill disease in famed Atlantic salmon (Salmo salar). Bull. Eur. Assoc. Fish Pathol. 2001, 21, 114–116. [Google Scholar]
- Young, N.D.; Dyková, I.; Snekvik, K.; Nowak, B.F.; Morrison, R.N. Neoparamoeba perurans is a cosmopolitan aetiological agent of amoebic gill disease. Dis. Aquat. Org. 2008, 78, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Report of the Working Group on Pathology and Diseases of Marine Organisms (WGPDMO); ICES CM 2015/SSGEPI:01; Helsinki, Finland, 24–28 February 2015; ICES: Copenhagen, Denmark, 2015; p. 124.
- Interim Report of the Working Group on Pathology and Diseases of Marine Organisms (WGPDMO); ICES CM 2016/SSGEPI:07; 17–20 February 2016, VA, USA; ICES: Copenhagen, Denmark, 2016; p. 18.
- DFO 2020 Fish Health Events Graph|DFO Public Reporting on Aquaculture|Pacific Region|Fisheries and Oceans Canada. Available online: dfo-mpo.gc.ca (accessed on 20 April 2022).
- Herrero, A.; Thompson, K.D.; Ashby, A.; Rodger, H.D.; Dagleish, M.P. Complex gill disease: An emerging syndrome in farmed Atlantic salmon (Salmo salar L.). J. Comp. Pathol. 2018, 163, 23–28. [Google Scholar] [CrossRef]
- Fringuelli, E.; Gordon, A.; Rodger, H.; Welsh, M.; Graham, D. Detection of Neoparamoeba perurans by duplex quantitative Taqman real-time PCR in formalin-fixed, paraffin-embedded Atlantic salmonid gill tissues. J. Fish Dis. 2012, 35, 711–724. [Google Scholar] [CrossRef]
- Christensen, R.H.B. “Ordinal—Regression Models for Ordinal Data.” R Package Version 2019.12-10. Available online: https://CRAN.R-project.org/package=ordinal (accessed on 20 April 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 20 April 2022).
- Taylor, R.S.; Muller, W.J.; Cook, M.T.; Kube, P.D.; Elliot, N.G. Gill observations in Atlantic salmon (Salmo salar, L.) during repeated amoebic gill disease (AGD) field exposure and survival challenge. Aquaculture 2009, 290, 1–8. [Google Scholar] [CrossRef]
- Bloecher, N.; Powell, M.; Hytterød, S.; Gjessing, M.; Wiik-Nielsen, J.; Mohammad, S.N.; Johansen, J.; Hansen, H.; Floerl, O.; Gjervre, A.-G. Effects of cnidarian biofouling on salmon gill health and development of amoebic gill disease. PLoS ONE 2018, 13, e0199842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Król, E.; Noguera, P.; Shaw, S.; Costelloe, E.; Gajardo, K.; Valdenegro, V.; Bickerdike, R.; Douglas, A.; Martin, S.A.M. Integration of transcriptome, gross morphology and histopathology in the gill of farmed Atlantic salmon (Salmo salar): Lessons from multi-site sampling. Front. Genet. 2020, 11, 610. [Google Scholar] [CrossRef]
- Downes, J.K.; Yatabe, T.; Marcos-Lopez, M.; Rodger, H.D.; MacCarthy, E.; O’Connor, I.; Collins, E.; Ruane, N.M. Investigation of co-infections with pathogens associated with gill disease in Atlantic salmon during an amoebic gill disease outbreak. J. Fish Dis. 2018, 41, 1217–1227. [Google Scholar] [CrossRef]
- Gjessing, M.C.; Steinum, T.; Olsen, A.B.; Lie, K.I.; Tavornpanich, S.; Colquhoun, D.J.; Gjevre, A.-G. Histopathological investigation of complex gill disease in sea farmed Atlantic salmon. PLoS ONE 2019, 14, e0222926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, E.J.; Rodger, H.D.; McAllen, R.; Doyle, T.K. Gill disorders in marine farmed salmon: Investigating the role of hydrozoan jellyfish. Aquacult. Env. Interac. 2011, 1, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Baxter, E.J.; Sturt, M.M.; Ruane, N.M.; Doyle, T.K.; McAllen, R.; Harman, L.; Rodger, H.D. Gill damage to Atlantic salmon (Salmo salar) caused by the common jellyfish Aurelia aurita under experimental challenge. PLoS ONE 2011, 6, e18529. [Google Scholar] [CrossRef]
- Baxter, E.J.; Sturt, M.M.; Ruane, N.M.; Doyle, T.K.; McAllen, R.; Rodger, H.D. Biofouling of the hydroid Ectopleura larynx on aquaculture nets in Ireland: Implications for finfish health. Fish Vet. J. 2012, 13, 18–30. [Google Scholar]
- Marcos-Lopez, M.; Mitchell, S.O.; Rodgers, H.D. Pathology and mortality associated with the mauve stinger jellyfish Pelagia noctiluca in farmed Atlantic salmon Salmo salar L. J. Fish Dis. 2016, 39, 111–115. [Google Scholar] [CrossRef]
- Powell, M.D.; Atland, A.; Dale, T. Acute lion’s mane jellyfish, Cyanea capillata (Cnideria: Scyphozoa), exposure to Atlantic salmon (Salmo salar L.). J. Fish Dis. 2018, 41, 751–759. [Google Scholar] [CrossRef]
- Kintner, A.; Brierley, A.S. Cryptic hydrozoan blooms pose risks to gill health in farmed North Atlantic salmon (Salmo salar). J. Mar. Biol. Assoc. UK 2019, 99, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Rodger, H.D. Gill disorders: An emerging problem for farmed Atlantic salmon (Salmo salar) in the marine environment? Fish Vet. J. 2007, 9, 38–48. [Google Scholar]
- Østevik, L.; Stormoen, M.; Nødtvedt, A.; Alarcón, M.; Lie, K.-I.; Skagøy, A.; Rodger, H. Assessment of acute effects of in situ net cleaning on gill health of farmed Atlantic salmon (Salmo salar L). Aquaculture 2021, 545, 737203. [Google Scholar] [CrossRef]
- Benedicenti, O.; Pottinge, T.G.; Collins, C.; Secombes, C.J. Effects of temperature on amoebic gill disease development: Does it play a role? J. Fish Dis. 2019, 42, 1241–1258. [Google Scholar] [CrossRef] [PubMed]
- Foyle, K.L.; Hess, S.; Powell, M.D.; Herbert, N.A. What is gill health and what is its role in marine fish aquaculture in the face of a changing climate? Front. Mar. Sci. 2020, 7, 400. [Google Scholar] [CrossRef]
- Tan, C.K.; Nowak, B.F.; Hodson, S.L. Biofouling as a reservoir of Neoparamoeba pemaquidensis (Page, 1970), the causative agent of amoebic gill disease in Atlantic salmon. Aquaculture 2002, 210, 49–58. [Google Scholar] [CrossRef]
- Hellebø, A.; Stene, A.; Aspehaug, V. PCR survey for Paramoeba perurans in fauna, environmental samples and fish associated with marine farming sites for Atlantic salmon (Salmo salar L.). J. Fish Dis. 2017, 40, 661–670. [Google Scholar] [CrossRef]
- Floerl, O.; Sunde, L.M.; Bloecher, N. Potential environmental risks associated with biofouling management in aquaculture. Aquacult. Env. Interac. 2016, 8, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Kent, M.L.; Traxler, G.S.; Kieser, D.; Richard, J.; Dawe, S.C.; Shaw, R.W.; Prosperi-Porta, G.; Ketcheson, J.; Evelyn, T.P.T. Survey of salmonid pathogens in ocean-caught fishes in British Columbia. J. Aquat. Anim. Health 1998, 10, 211–219. [Google Scholar] [CrossRef]
- DiLorenzo, E.; Mantua, N. Multi-year persistence of the 2014/15 North Pacific marine heatwave. Nat. Clim. Chang. 2016, 6, 1042–1047. [Google Scholar] [CrossRef]
- Chen, Z.; Shi, J.; Li, C. Two types of warm blobs in the Northeast Pacific and their potential effect on the El Niño. Int. J. Climatol. 2021, 41, 2810–2827. [Google Scholar] [CrossRef]
- Yang, Q.; Cokelet, E.D.; Stabeno, P.J.; Li, L.; Hollowed, A.B.; Pallson, W.A.; Bond, N.A.; Barbeaux, S.J. How “The Blob” affected groundfish distributions in the Gulf of Alaska. Fish. Oceanog. 2019, 28, 434–453. [Google Scholar] [CrossRef]
- Traving, S.J.; Kellogg, C.T.E.; Ross, T.; McLaughlin, R.; Kieft, B.; Ho, G.Y.; Peña, A.; Krzywinski, M.; Robert, M.; Hallam, S.J. Prokaryotic responses to a warm temperature anomaly in northeast subarctic Pacific waters. Comm. Biol. 2021, 4, 1217. [Google Scholar] [CrossRef] [PubMed]
- Tate, H.M.; Studholme, K.R.; Domalik, A.D.; Drever, M.C.; Romero, L.M.; Gormally, B.M.G.; Hobson, K.A.; Hipfner, J.M.; Crossin, G.T. Interannual measures of nutritional stress during a marine heatwave (the Blob) differ between two North Pacific seabird species. Conserv. Physiol. 2021, 9, coab090. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Score | Filament Damage | Extent (%) |
|---|---|---|
| 0 | None visible | 0 |
| 1 | Limited thickening, or very few affected | <10 |
| 2 | Frequent thickening of tips | 10–25 |
| 3 | Thickening on most tips, extending to ≤50% of length | 25–50 |
| 4 | Thickening on most, to more than 50% of length | 50–75 |
| 5 | Most or all thickened along entire length | 75–100 |
| Score | N (%) | Pathology 1 | |||||
|---|---|---|---|---|---|---|---|
| GLH/GLF | PAP | GFB | GLT | BST | GTH | ||
| <2 | 1437 (76.4) | 39 (2.7) | 8 (0.6) | 15 (1.0) | 9 (0.6) | 8 (0.6) | 8 (0.6) |
| ≥2 | 444 (23.6) | 55 (12.4) | 11 (2.5) | 22 (5.0) | 6 (1.4) | 6 (1.4) | 4 (0.9) |
| Days at Sea | Nt | N. perurans | Gill Score | ||
|---|---|---|---|---|---|
| Np | Mean Ct | N ≥ 2 | Mean | ||
| 0–199 | 171 | 5 (2.9) | 36.45 | 23 (13.4) | 0.47 |
| 200–299 | 103 | 9 (8.7) | 28.27 | 9 (8.7) | 0.38 |
| 300–399 | 112 | 13 (11.6) | 32.28 | 24 (21.4) | 0.72 |
| 400–499 | 159 | 8 (5.0) | 34.67 | 52 (32.7) | 1.12 |
| 500–599 | 106 | 3 (2.3) | 33.37 | 40 (37.7) | 1.26 |
| 600–699 | 25 | 5 (20.0) | 33.36 | 5 (16.0) | 1.00 |
| 700–799 | 3 | 3 (100.0) | 30.04 | 0 | 1.00 |
| Term | Estimate | Std. Error | 95% Conf. Interval | p Value |
|---|---|---|---|---|
| Gill score thresholds | ||||
| 0|1 | 0.31 | 0.10 | 0.12–0.50 | |
| 1|2 | 1.42 | 0.10 | 1.22–1.63 | |
| 2|3 | 2.34 | 0.12 | 2.11–2.56 | |
| 3|4 | 3.32 | 0.14 | 3.05–3.60 | |
| 4|5 | 4.63 | 0.21 | 4.21–5.04 | |
| N. perurans | −0.27 | 0.25 | −0.77–0.22 | |
| Salinity (scaled) | −0.03 | 0.07 | −0.16–0.09 | |
| Temperature (scaled) | 0.19 | 0.08 | 0.05–0.34 | 0.01 |
| N. perurans × Salinity (scaled) | 0.59 | 0.26 | 0.08–1.09 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, S.R.M.; Price, D. Elevated Seawater Temperature and Infection with Neoparamoeba perurans Exacerbate Complex Gill Disease in Farmed Atlantic Salmon (Salmo salar) in British Columbia, Canada. Microorganisms 2022, 10, 1039. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051039
Jones SRM, Price D. Elevated Seawater Temperature and Infection with Neoparamoeba perurans Exacerbate Complex Gill Disease in Farmed Atlantic Salmon (Salmo salar) in British Columbia, Canada. Microorganisms. 2022; 10(5):1039. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051039
Chicago/Turabian StyleJones, Simon R. M., and Derek Price. 2022. "Elevated Seawater Temperature and Infection with Neoparamoeba perurans Exacerbate Complex Gill Disease in Farmed Atlantic Salmon (Salmo salar) in British Columbia, Canada" Microorganisms 10, no. 5: 1039. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051039