
The Interactions between Arbuscular Mycorrhizal Fungi and Trichoderma longibrachiatum Enhance Maize Growth and Modulate Root Metabolome under Increasing Soil Salinity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the T. longibrachiatum MF Inoculant
2.2. Preparation of the Soil
2.3. Microcosm Experiment
2.3.1. Experiment 1
2.3.2. Experiment 2
2.4. Harvest and Sample Preparation
2.5. Determination of Colonization and Hyphal Length Density of AM Fungi
2.6. The Population of T. longibrachiatum MF
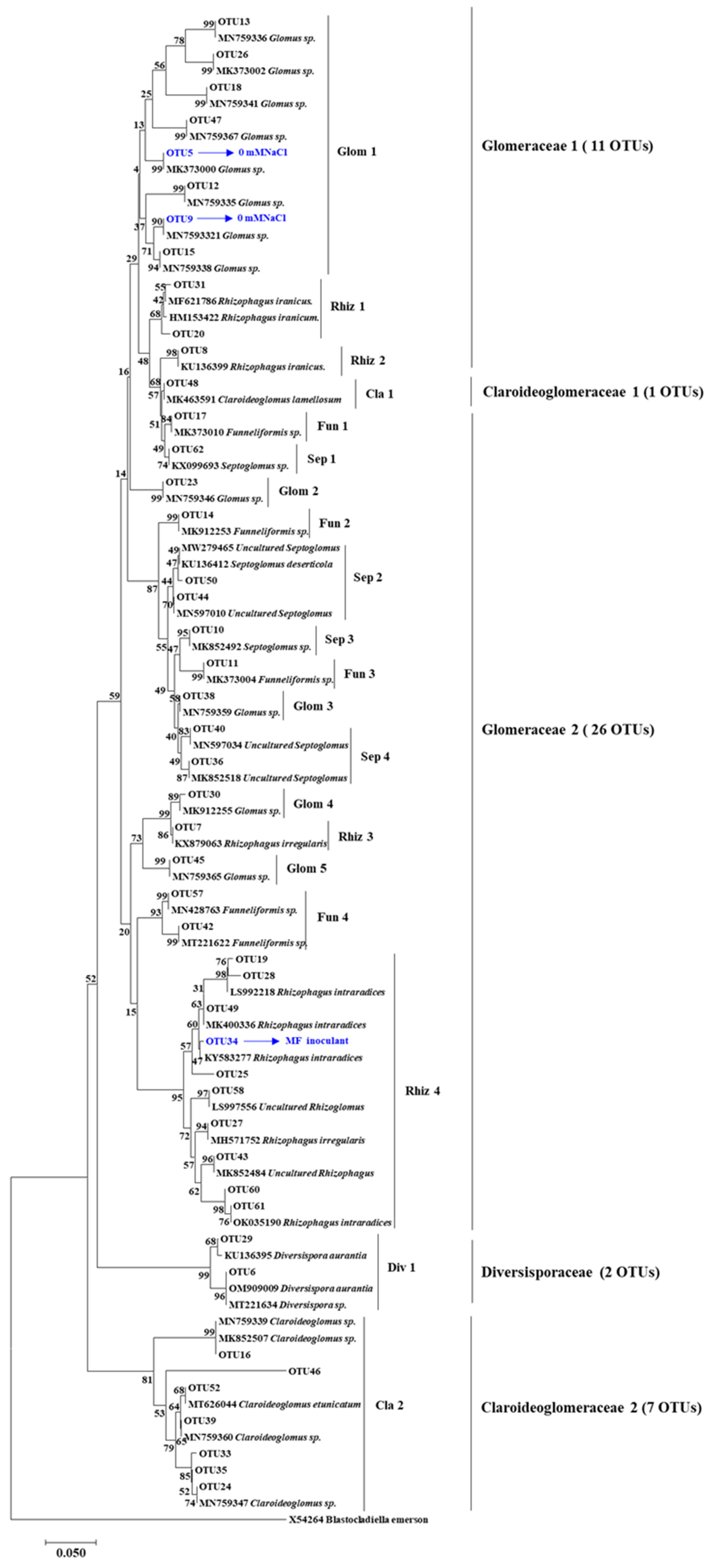
2.7. AM Fungal Community Structure Analysis
2.8. Profiling of Metabolites
2.9. Data Analysis
3. Results
3.1. Experiment 1
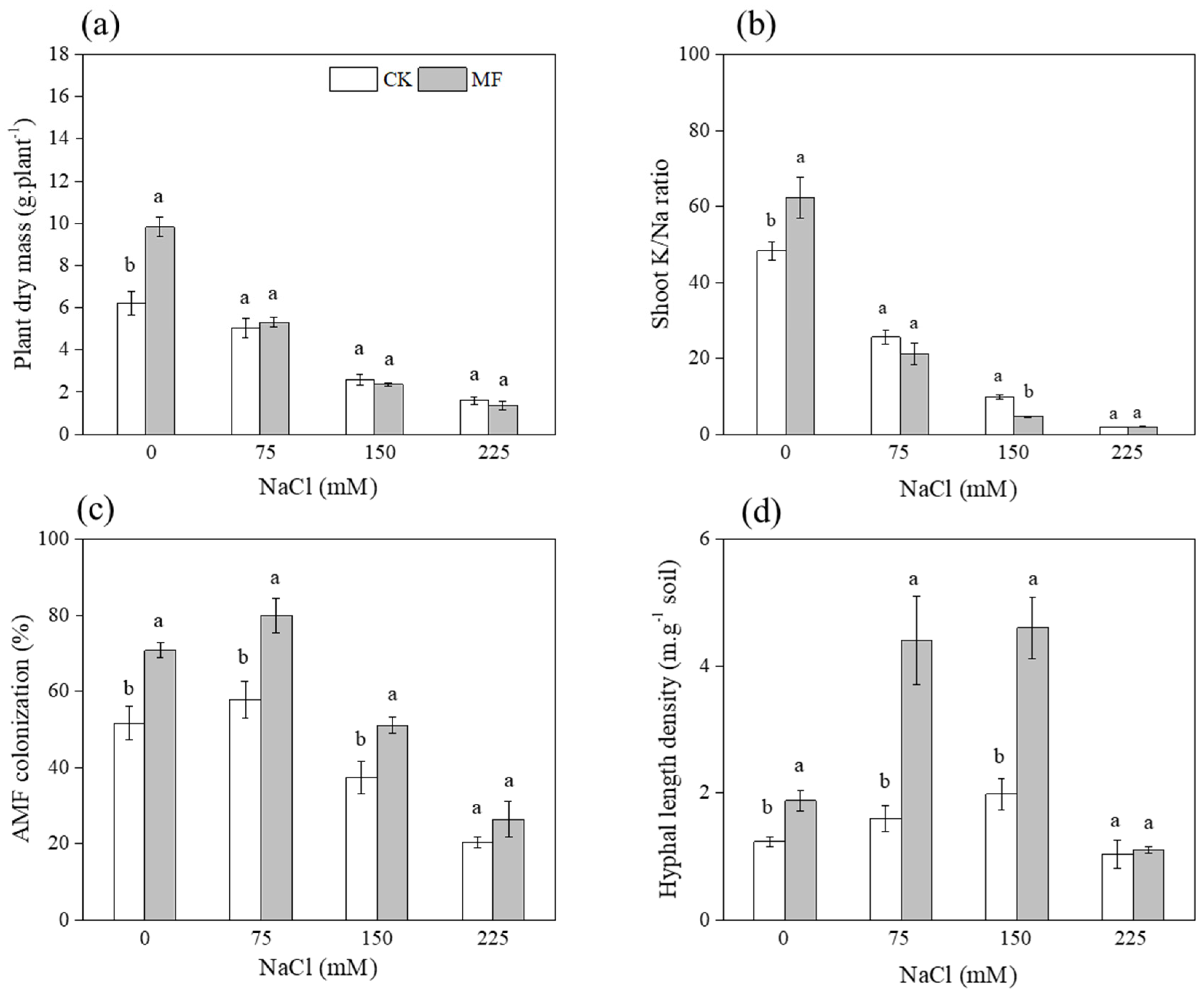
3.1.1. T. longibrachiatum MF, AM Fungi and Maize Performance
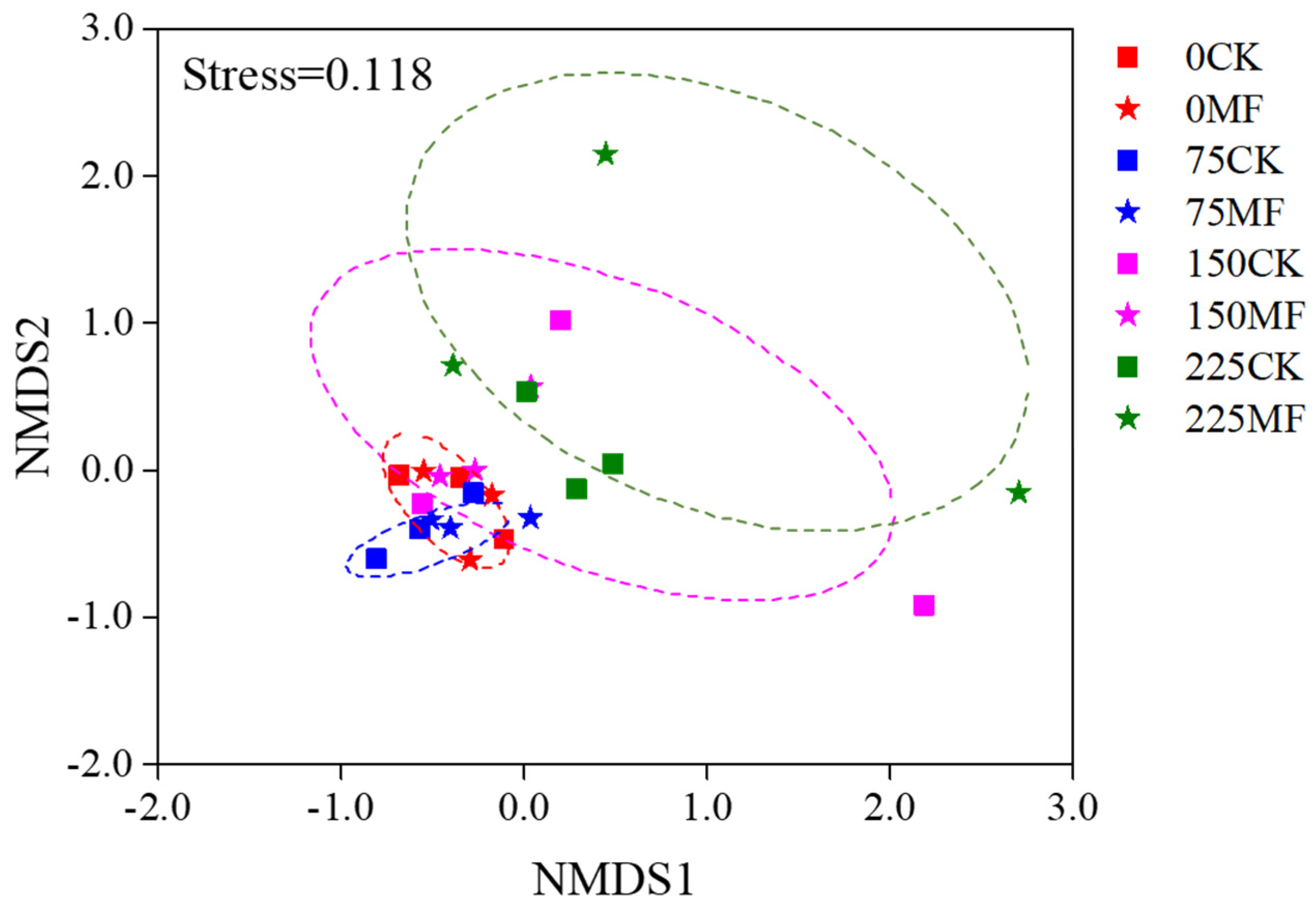
3.1.2. AM Fungal Structure in Maize Roots
3.2. Experiment 2
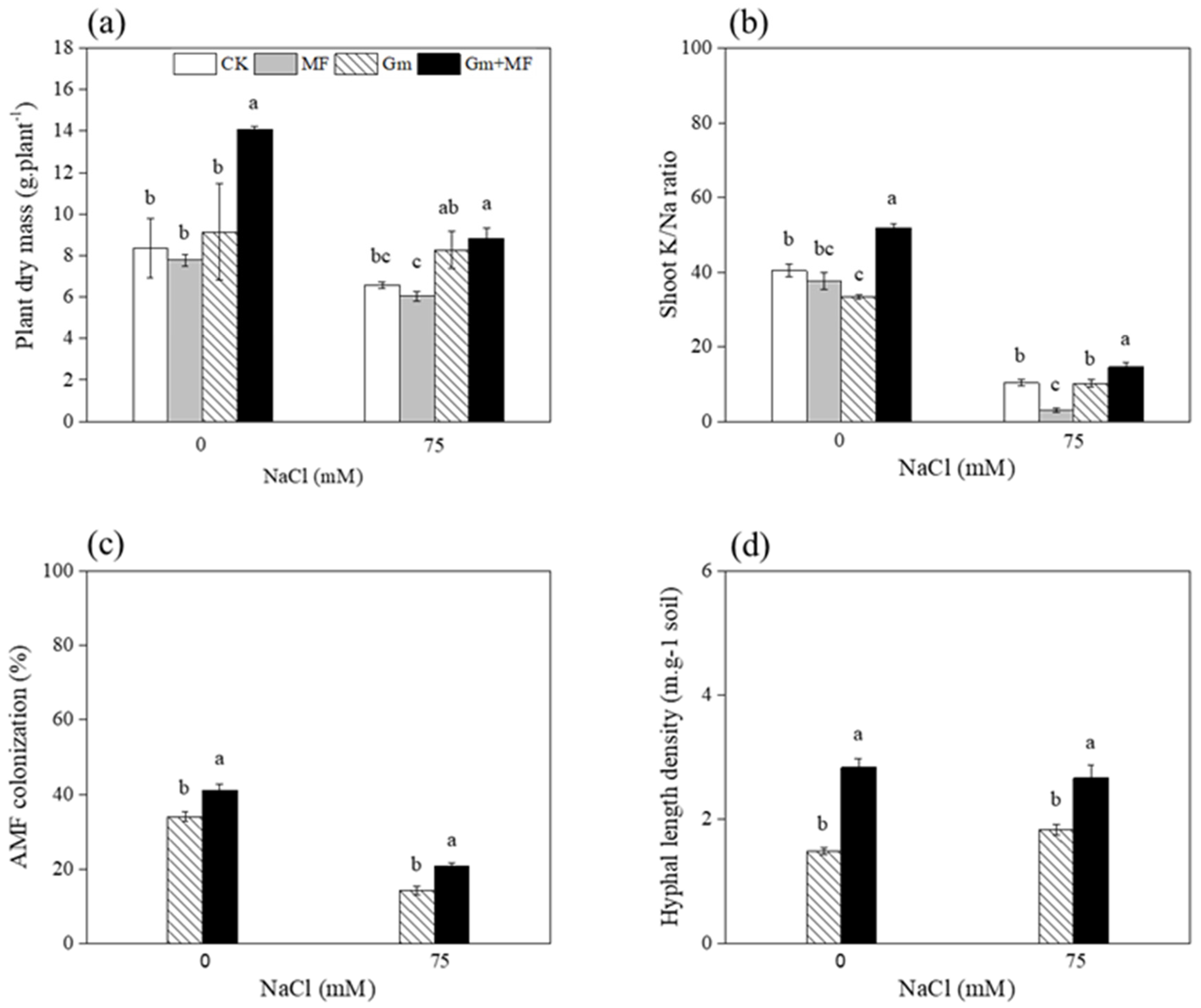
3.2.1. T. longibrachiatum MF, AM Fungi and Maize Performance
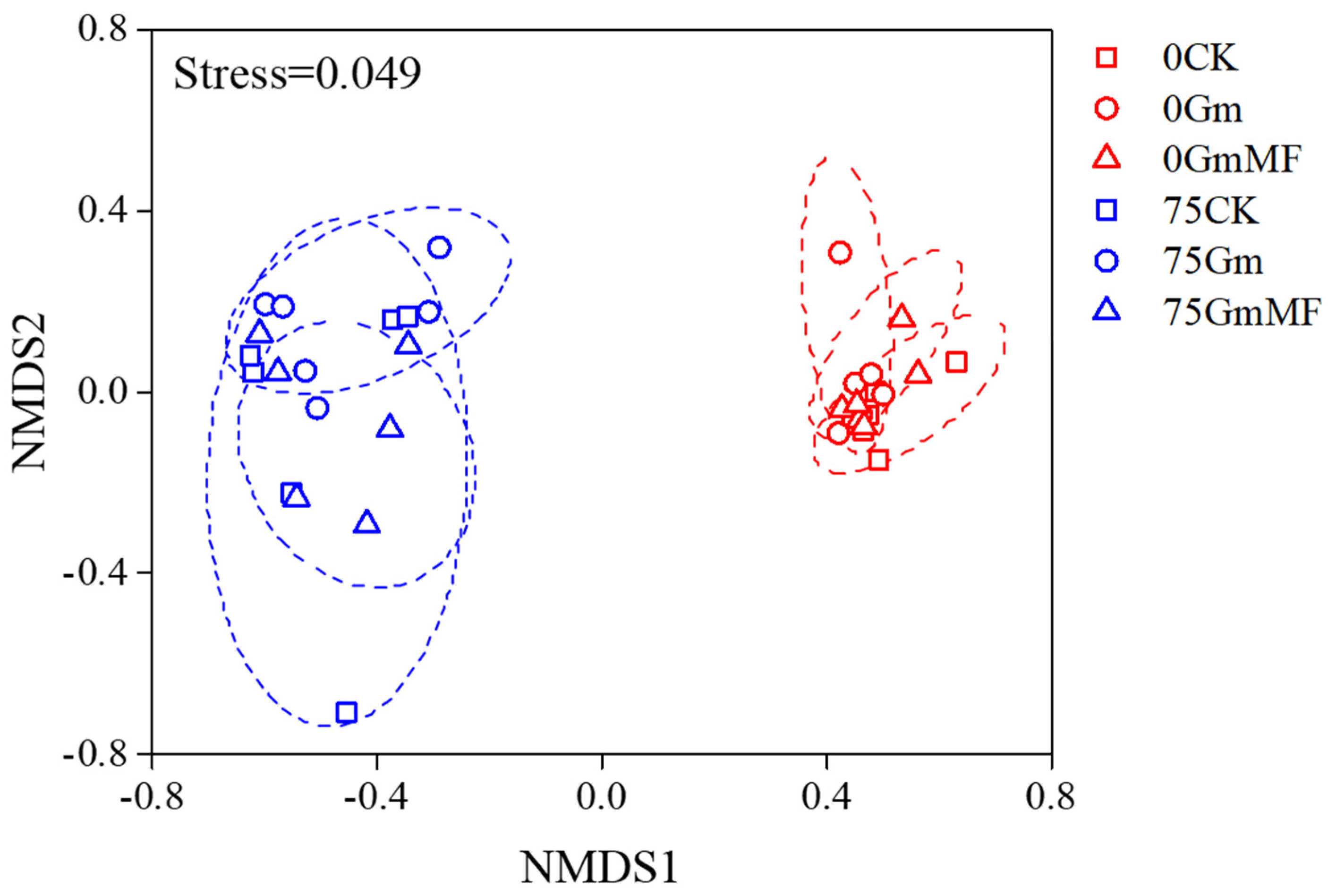
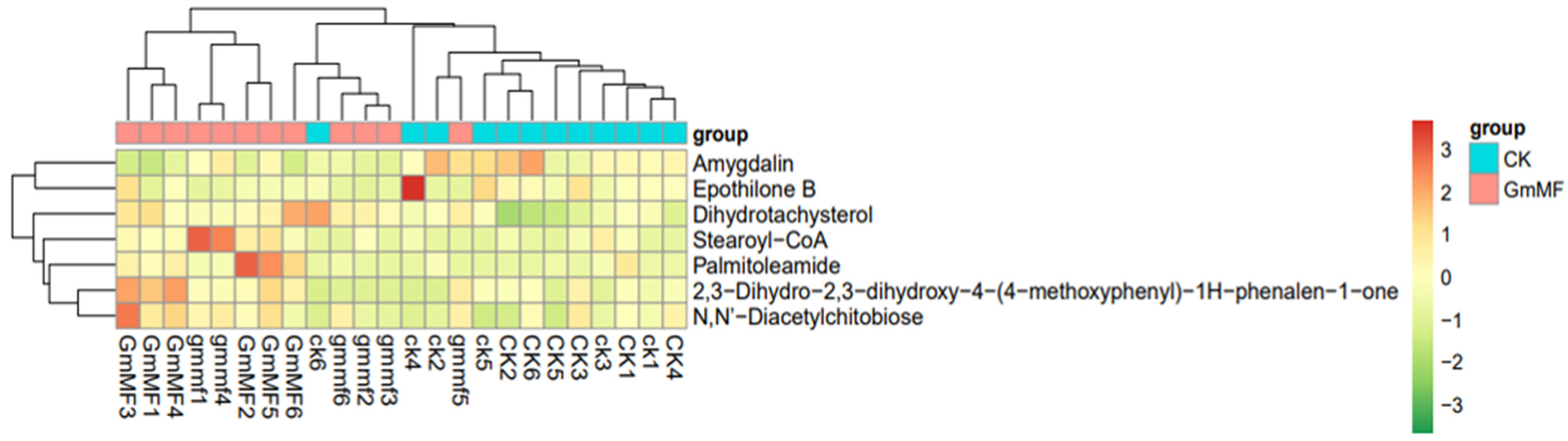
3.2.2. Metabolic Profiles in Maize Roots
4. Discussion
4.1. T. longibrachiatum MF Improved AM Fungi Growth
4.2. The Dual Inoculation Enhanced Plant Growth
4.3. The Dual Inoculation Modulated Root Metabolome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [Green Version]
- Hidri, R.; Barea, J.M.; Metoui-Ben, M.O.; Abdelly, C.; Azcon, R. Impact of microbial inoculation on biomass accumulation by Sulla carnosa provenances, and in regulating nutrition, physiological and antioxidant activities of this species under non-saline and saline conditions. J. Plant Physiol. 2016, 201, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Vimal, S.R.; Singh, J.S.; Arora, N.K.; Singh, S. Soil-plant-microbe interactions in stressed agriculture management: A review. Pedosphere 2017, 27, 177–192. [Google Scholar] [CrossRef]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushio, M.; Nakaoka, S.; Onoda, Y. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Hidri, R.; Mahmoud, M.B.; Debez, A.; Abdelly, C.; Azcon, R. Modulation of c:n:p stoichiometry is involved in the effectiveness of a PGPR and am fungus in increasing salt stress tolerance of sulla carnosa tunisian provenances. App. Soil Ecol. 2019, 143, 161–172. [Google Scholar] [CrossRef]
- Ghaffari, M.R.; Ghabooli, M.; Khatabi, B.; Hajirezaei, M.R.; Schweizer, P.; Salekdeh, G.H. Metabolic and transcriptional response of central metabolism affected by root endophytic fungus Piriformospora indica under salinity in barley. Plant Mol. Biol. 2016, 90, 699–717. [Google Scholar] [CrossRef] [PubMed]
- Gul Jan, F.; Hamayun, M.; Hussain, A.; Jan, G.; Iqbal, A.; Khan, A.; Lee, I.-J. An endophytic isolate of the fungus yarrowia lipolytica produces metabolites that ameliorate the negative impact of salt stress on the physiology of maize. BMC Microbial. 2019, 19, 3. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Xiao, Y.; Wang, Y.F.; Liu, Z.H.; Yang, K.J. Trichoderma affects the physiochemical characteristics and bacterial community composition of saline-alkaline maize rhizosphere soils in the cold-region of Heilongjiang province. Plant Soil 2019, 436, 211–227. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbial. 2004, 1, 43–56. [Google Scholar] [CrossRef]
- Bae, H.; Roberts, D.P.; Lim, H.S.; Strem, M.D.; Park, S.C.; Ryu, C.M. Endophytic trichoderma isolates from tropical environments delay disease onset and induce resistance against phytophthora capsici in hot pepper using multiple mechanisms. Mol. Plant Microbial. Interact. 2011, 24, 336–351. [Google Scholar] [CrossRef] [Green Version]
- Wazny, R.; Rozpadek, P.; Jedrzejczyk, R.J.; Sliwa, M.; Stojakowska, A.; Anielska, T.; Turnau, K. Does co-inoculation of lactuca serriola with endophytic and arbuscular mycorrhizal fungi improve plant growth in a polluted environment? Mycorrhiza 2018, 3, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, S.M.; Ahmadm, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotech. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Li, L.; Lindström, K.; Räsänen, L. A synergistic interaction between salt tolerant Pseudomonas and Mezorhizobium strains improves growth and symbiotic performance ofliquorice (Glycyrrhiza uralensis Fish.) under salt stress. Apply Microbial. Biotechnol. 2016, 100, 2829–2841. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcon, C.; Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef] [Green Version]
- Astrit, B.; Glenda, S.; Boris, R. AMF inoculation enhances growth and improves the nutrient uptake rates of transplanted, salt-stressed tomato seedlings. Sustainability 2015, 12, 15967–15981. [Google Scholar] [CrossRef] [Green Version]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Marulanda-Aguirre, A.; Azcon, R.; Ruiz-Lozano, J.M.; Aroca, R. Differential effects of a Bacillus megaterium strain on Lactucasativa plant growth depending on the origin of the arbuscular mycorrhizal fungus coinoculated: Physiologic and biochemical traits. J. Plant Growth Regul. 2008, 27, 10–18. [Google Scholar] [CrossRef]
- Toro, M.; Azcón, R.; Barea, J.M. Improvement of arbuscular mycorrhiza development by inoculation of soil with phosphate-solubilizing rhizobacteria to improve rock phosphate bioavailability (32P) and nutrient cycling. Appl. Environ. Microbial. 1997, 63, 4408–4412. [Google Scholar] [CrossRef] [Green Version]
- Lagos, C.; Larsen, J.; Correa, E.S.; Almonacid, L.; Héctor, H.; Fuentes, A.; Arriagada, C. Dual inoculation with mycorrhizal and saprotrophic fungi suppress the maize growth and development under phenanthrene exposure. J. Soil Sci. Plant Nutr. 2018, 3, 721–734. [Google Scholar] [CrossRef] [Green Version]
- Chandanie, W.A.; Kubota, M.; Hyakumachi, M. Interactions between the arbuscular mycorrhizal fungus Glomus mosseae and plant growth promoting fungi and their significance for enhancing plant growth and suppressing damping-off of cucumber (Cucumis sativus L.). Appl. Soil Ecol. 2009, 41, 336–341. [Google Scholar] [CrossRef]
- Singh, S.; Kapoor, K.K. Inoculation with phosphate-solubilizing microorganisms and a vesicular-arbuscular mycorrhizal fungus improves dry matter yield and nutrient uptake by wheat grown in a sandy soil. Biol. Fertil. Soil 1999, 28, 139–144. [Google Scholar] [CrossRef]
- Osorio, N.W.; Habte, M. Effect of a phosphate-solubilizing fungus and an arbuscular mycorrhizal fungus on leucaena seedlings in tropical soils with contrasting phosphate sorption capacity. Plant Soil 2015, 389, 375–385. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt tolerance mechanisms. Trends Plant Sci. 2014, 6, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Rivero, J.; Álvarez, D.; Flors, V.; Azcon-Aguilar, C. Root metabolic plasticity underlies functional diversity in mycorrhiza-enhanced stress tolerance in tomato. New Phytol. 2018, 220, 1322–1336. [Google Scholar] [CrossRef] [Green Version]
- Rodziewicz, P.; Swarcewicz, B.; Chmielewska, K.; Wojakowska, A.; Stobiecki, M. Influence of abiotic stresses on plant proteome and metabolome changes. Acta Physiol. Plant. 2014, 36, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Wang, J.J.; Xu, S.; Zhao, W.; Liu, H.Y.; Li, Q.Q.; Huang, Z.Y. Screening, identification and salt-tolerant characteristics of phosphate-solubilizing fungi. Microbiol. China 2018, 10, 2142–2151. [Google Scholar] [CrossRef]
- Yuan, S.F.; Li, M.Y.; Fang, Z.Y.; Liu, Y.; Shi, W.; Pan, B.; Wu, K.; Shi, J.X.; Shen, B.; Shen, Q.R. Biological control of tobacco bacterial wilt using Trichoderma harzianum amended bioorganic fertilizer and the arbuscular mycorrhizal fungi Glomus Mosseae. Biol. Control 2016, 92, 164–171. [Google Scholar] [CrossRef]
- Yang, R.; Qin, Z.F.; Wang, J.J.; Xu, S.; Zhao, W.; Zhang, X.X.; Huang, Z.Y. Salinity changes root occupancy by arbuscular mycorrhizal fungal species. Pedobiol. J. Soil Biol. 2020, 81, 150665. [Google Scholar] [CrossRef]
- Utobo, E.B.; Ogbodo, E.N.; Nwogbaga, A.C. Techniques for extraction and quantification of arbuscular rmycorrhizal fungi. Libyan Agric. Res. Center Int. 2011, 2, 68–78. [Google Scholar]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.M.; Reinhardt, D.R.; Jastrow, J.D. External hyphal production of vesicular arbuscular mycorrhizal fungi in pasture and tallgrass prairie communities. Oecologia 1995, 103, 17–23. [Google Scholar] [CrossRef]
- Efthymiou, A.K.; Jensen, B.; Jakobsen, I. The roles of mycorrhiza and penicillium inoculants in phosphorus uptake by biochar-amended wheat. Soil Biol. Biochem. 2018, 127, 168–177. [Google Scholar] [CrossRef]
- Lumini, E.; Orgiazzi, A.; Borriello, R.; Bonfante, P.; Bianciotto, V. Disclosing arbuscular mycorrhizal fungal biodiversity in soil through a land-use gradient using a pyrosequencing approach. Environ. Microbial. 2010, 12, 2165–2179. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- de Caceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Kohler, J.; Hernandez, J.A.; Caravaca, F.; Roldan, A. Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 2009, 65, 245–252. [Google Scholar] [CrossRef]
- Kanchan Singh, N. Organic amendments to soil inoculated arbuscular mycorrhizal fungi and Pseudomonas fluorescens treatments reduce the development of root-rot disease and enhance the yield of Phaseolus vulgaris L. Eur. J. Soil Biol. 2011, 47, 288–295. [Google Scholar] [CrossRef]
- Ruíz-Sánchez, M.; Armada, E.; Munoz, Y.; de Salamone, I.E.G.; Aroca, R.; Ruíz-Lozano, J.M.; Azcón, R. Azospirillum and arbuscular mycorrhizal colonization enhance rice growth and physiological traits under wellwatered and drought conditions. J. Plant Physiol. 2011, 168, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Almethyeb, M.; Ruppel, S.; Paulsen, H.M.; Vassilev, N.; Eichler-Löbermann, B. Single and combined applications of arbuscular mycorrhizal fungi and Enterobacter radicincitans affect nutrient uptake of faba bean and soil biological characteristics. Agric. Forest Res. 2013, 3, 229–234. [Google Scholar] [CrossRef]
- Garg, N.; Chandel, S. Role of arbuscular mycorrhiza in arresting reactive oxygen species (ROS) and strengthening antioxidant defense in Cajanus cajan (L.) Millsp. Nodules under salinity (NaCl) and cadmium (Cd) stress. Plant Growth Regul. 2015, 75, 521–534. [Google Scholar] [CrossRef]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J.M. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fertil. Soi. 2003, 1, 1–16. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kim, K.; Subramanian, P.; Senthilkumar, M.; Anandham, R.; Sa, T. Arbuscular mycorrhizal fungi and associated bacteria isolated from saltaffected soil enhances the tolerance of maize to salinity in coastal reclamation soil. Agric. Ecosyst. Environ. 2016, 23, 233–239. [Google Scholar] [CrossRef]
- Van Geel, M.; Jacquemyn, H.; Plue, J.; Saar, L.; Ceulemans, T. Abiotic rather than biotic filtering shapes the arbuscular mycorrhizal fungal communities of European seminatural grasslands. New Phytol. 2017, 220, 1262–1272. [Google Scholar] [CrossRef] [Green Version]
- ÖPik, M.; Moora, M.; Liira, J.; Kõljalg, U.; Zobel, M.; Seb, R. Divergent arbuscular mycorrhizal fungal communities colonize roots of Pulsatilla spp. in boreal scots pine forest and grassland soils. New Phytol. 2003, 3, 581–593. [Google Scholar] [CrossRef] [Green Version]
- Yamato, M.; Ikeda, S.; Iwase, K. Community of arbuscular mycorrhizal fungi in a coastal vegetation on okinawa island and effect of the isolated fungi on growth of sorghum under salt-treated conditions. Mycorrhiza 2008, 5, 241–249. [Google Scholar] [CrossRef]
- Pang, G.; Cai, F.; Li, R.; Zhao, Z.; Li, R.; Gu, X.; Shen, Q.R.; Chen, W. Trichoderma-enriched organic fertilizer can mitigate microbiome degeneration of monocropped soil to maintain better plant growth. Plant Soil 2017, 416, 181–192. [Google Scholar] [CrossRef]
- Helena, M.; Sofia, I.A.P.; Alberto, V.; Paula, M.L.; Castro, A.P.G.C.M. Synergistic effects of arbuscular mycorrhizal fungi and plant growth-promoting bacteria benefit maize growth under increasing soil salinity. J. Environ. Manag. 2020, 257, 109982. [Google Scholar] [CrossRef]
- Ashrafi, E.; Zahedi, M.; Razmjoo, J. Co-inoculations of arbuscular mycorrhizal fungi and rhizobia under salinity in alfalfa. Soil Sci. Plant Nutr. 2014, 60, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Osorio, N.W.; Habte, M. Synergistic effect of a phosphate-solubilizing fungus and an arbuscular mycorrhizal fungus on leucaena seedlings in an oxisol fertilized with rock phosphate. Botany 2013, 4, 274–281. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Sui, X.; Fan, X.X.; Jia, T.T.; Song, F.Q. Arbuscular mycorrhizal symbiosis modulates antioxidant response and ion distribution in salt-stressed Elaeagnus angustifolia seedlings. Front. Microbial. 2018, 9, 652. [Google Scholar] [CrossRef] [Green Version]
- Amanifar, S.; Toghranegar, Z. The efficiency of arbuscular mycorrhiza for improving tolerance of valeriana officinalis L. and enhancing valerenic acid accumulation under salinity stress. Ind. Crops Prod. 2020, 147, 112234. [Google Scholar] [CrossRef]
- Abeer, H.; Elsayed, F.A.; Abdulaziz, A.A.; Asma, A.A.; Stephan, W.; Dilfuza, E. The Interaction between Arbuscular Mycorrhizal Fungi and Endophytic Bacteria Enhances Plant Growth of Acacia gerrardii under Salt Stress. Front. Microbial. 2016, 7, 868. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.J.; Zhang, N.L.; Zhang, L.L.; Zhang, X.L.; Wu, A.P.; Huang, J.Y.; Yu, S.Q.; Wang, Y.H. Mediation of arbuscular mycorrhizal fungi on growth and biochemical parameters of ligustrum vicaryi in response to salinity. Physiol. Mol. Plant Pathol. 2020, 112, 101522. [Google Scholar] [CrossRef]
- Ntambi, J.M.; Miyazaki, M. Regulation of stearoyl-CoA desaturases and role in metabolism. Prog. Lipid Res. 2004, 2, 91–104. [Google Scholar] [CrossRef]
- Shilpha, J.; Satish, L.; Kavikkuil, M.; Joe Virgin Largia, M.; Ramesh, M. Methyl jasmonate elicits the solasodine production and anti-oxidant activity in hairy root cultures of Solanum trilobatum L. Ind. Crops Prod. 2015, 71, 54–64. [Google Scholar] [CrossRef]
- Riemann, M.; Dhakarey, R.; Hazman, M.; Miro, B.; Kohli, A.; Nick, P. Exploring jasmonates in the hormonal network ofdrought and salinity responses. Front. Plant Sci. 2015, 6, 1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Salinity (S) | Inoculants (I) | Interaction S×I |
|---|---|---|---|
| Experiment 1 | |||
| Plant dry mass | 146.43 *** | 12.68 ** | 14.63 *** |
| Shoot K+/Na+ ratio | 199.86 *** | 0.42 ns | 6.97 ** |
| AMF colonization rate | 57.06 *** | 32.60 *** | 1.74 ns |
| HLD | 20.53 *** | 41.30 *** | 8.34 ** |
| Experiment 2 | |||
| Plant dry mass | 10.85 ** | 7.60 ** | 1.75 ns |
| Shoot K+/Na+ ratio | 1282.87 *** | 43.47 *** | 12.61 *** |
| AMF colonization rate | 233.02 *** | 26.52 ** | 0.54 ns |
| HLD | 0.41 ns | 63.82 *** | 3.69 ns |
| ID | Metabolite Name | CK vs. Gm | CK vs. Gm + MF | Gm vs. Gm + MF |
|---|---|---|---|---|
| meta_804 | Amygdalin | down | down | |
| meta_445 | Caproic acid | down | ||
| meta_430 | 6,10,14-Trimethyl-5,9,13-pentadecatrien-2-one | up | ||
| meta_291 | Epothilone B | down | ||
| meta_544 | Tylosin | up | ||
| meta_188 | Pro Met Met Thr | down | ||
| meta_196 | Chamazulene | down | ||
| meta_701 | 2,3-Dihydro-2,3-dihydroxy-4- (4-methoxyphenyl) -1H-phenalen-1-one | up | ||
| meta_696 | Stearoyl-CoA | up | ||
| meta_753 | Dihydrotachysterol | up | ||
| meta_789 | N,N′-Diacetylchitobiose | up | ||
| meta_627 | 3-Hydroxydodecanoic acid | up | ||
| meta_432 | Palmitoleamide | up | ||
| meta_239 | C16 Sphinganine | up | down | |
| meta_747 | N-Acetyl-a-neuraminic acid | up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Qin, Z.; Wang, J.; Zhang, X.; Xu, S.; Zhao, W.; Huang, Z. The Interactions between Arbuscular Mycorrhizal Fungi and Trichoderma longibrachiatum Enhance Maize Growth and Modulate Root Metabolome under Increasing Soil Salinity. Microorganisms 2022, 10, 1042. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051042
Yang R, Qin Z, Wang J, Zhang X, Xu S, Zhao W, Huang Z. The Interactions between Arbuscular Mycorrhizal Fungi and Trichoderma longibrachiatum Enhance Maize Growth and Modulate Root Metabolome under Increasing Soil Salinity. Microorganisms. 2022; 10(5):1042. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051042
Chicago/Turabian StyleYang, Rong, Zefeng Qin, Jingjing Wang, Xiaoxia Zhang, Song Xu, Wei Zhao, and Zhiyong Huang. 2022. "The Interactions between Arbuscular Mycorrhizal Fungi and Trichoderma longibrachiatum Enhance Maize Growth and Modulate Root Metabolome under Increasing Soil Salinity" Microorganisms 10, no. 5: 1042. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051042