
Monitoring the Starvation–Survival Response of Edwardsiella piscicida and E. tarda in Freshwater Microcosms, at Various Temperatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Preparation of the Filtered and Sterilized Lake Water (FSLW) and Inoculation of Microcosms
2.3. Cell Counts, Viability Assessment and Staining Methods
2.4. Resuscitation of VBNC Cells
2.5. Virulence of the Original Edwardsiella Strains and of Its Resuscitated Cultures for European Eel
2.6. Data Analysis and Statistics
3. Results
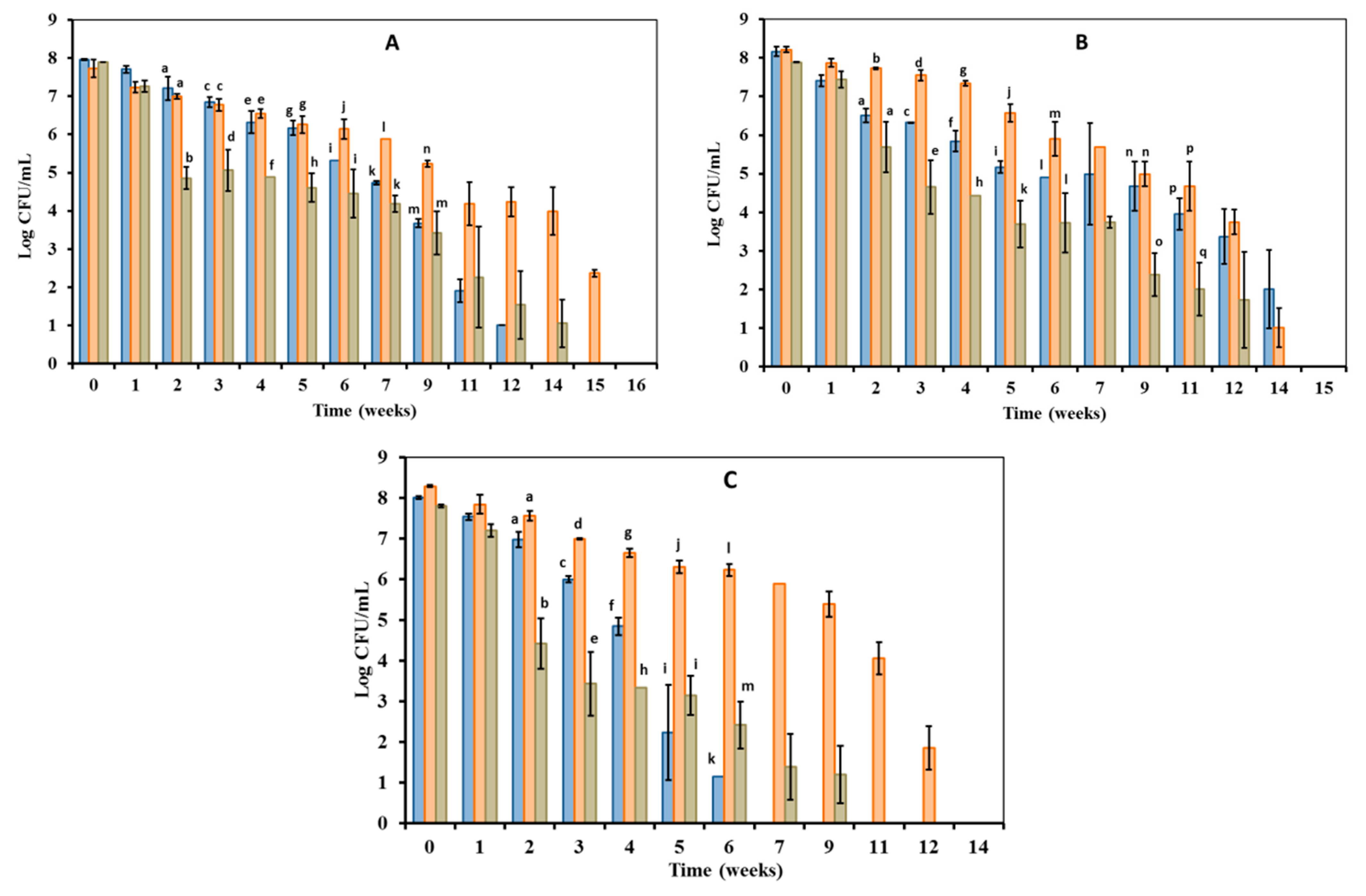
3.1. Culturability of Edwardsiella Maintained under Starvation Conditions at Various Temperatures
3.2. The VBNC State of Edwardsiella Maintained under Starvation Conditions at 7 °C and 25 °C
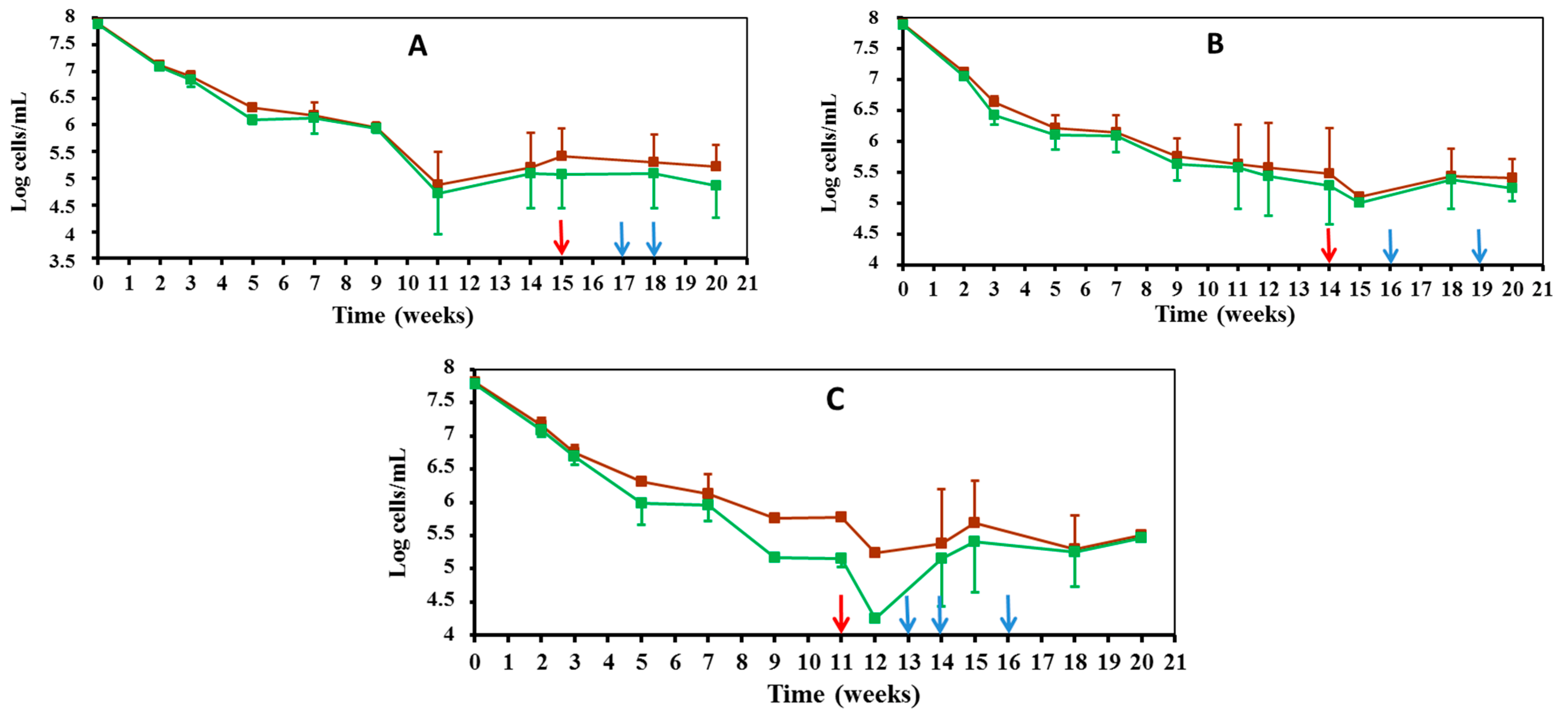
3.2.1. Direct Counts of Edwardsiella Cells during Culturable and VBNC Period
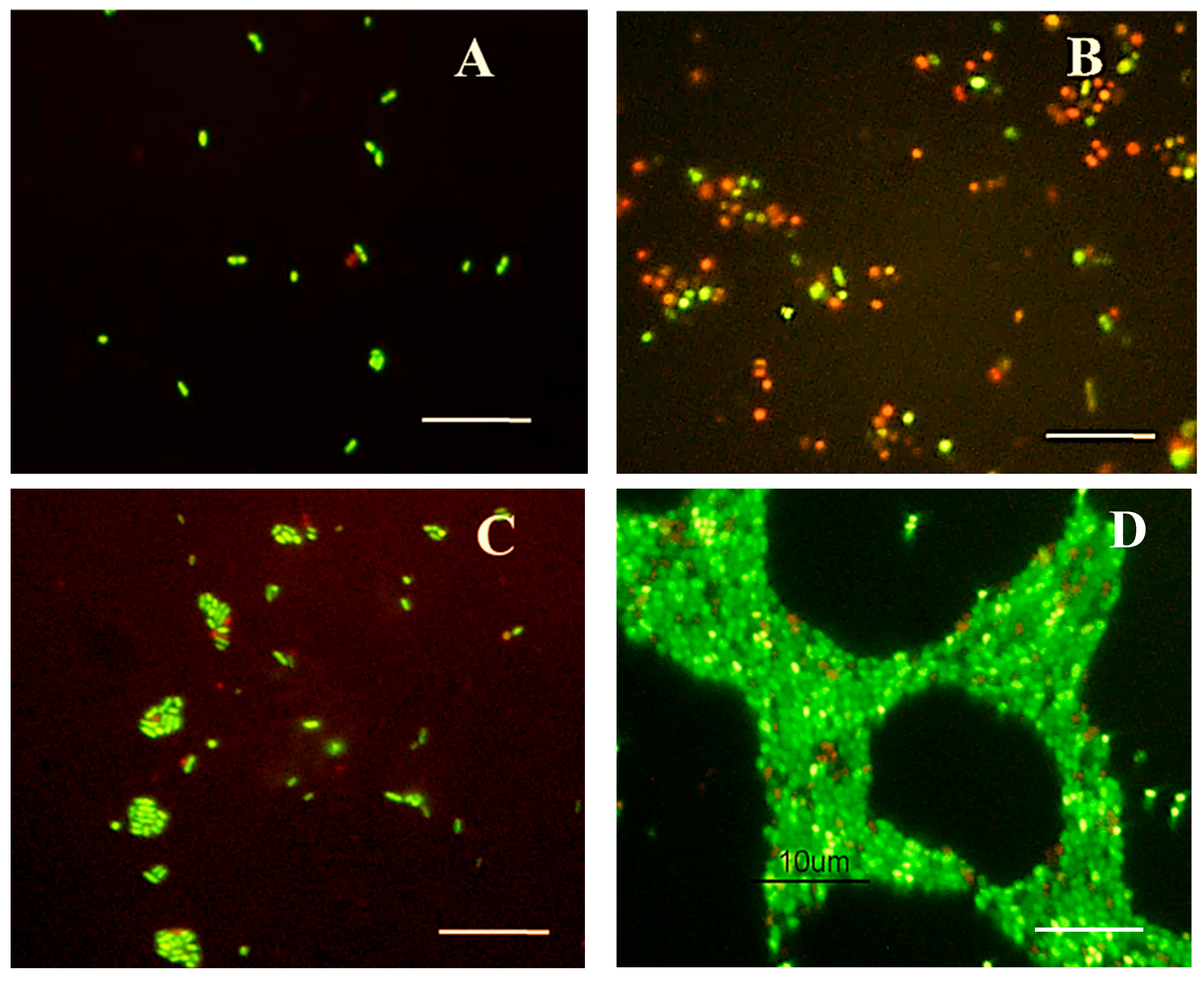
3.2.2. Morphological Changes Observed in Edwardsiella Cells during Starvation-Survival Response

3.2.3. Resuscitation of VBNC Edwardsiella Cells and Its Virulence for European Eel
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adelou, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the “Enterobacteriales”: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef]
- Hirai, Y.; Asahata-Tago, S.; Ainoda, Y.; Fijita, T.; Kikuchi, K. Edwardsiella tarda bacteremia. A rare but fatal water- and foodborne infection: Review of the literatura and clinical cases from a single centre. Can. J. Infect. Dis. Med. Microbiol. 2015, 26, 313–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, B.R.; Sahoo, P.K. Edwardsiellosis in fish: A bief review. J. Biosci. 2007, 32, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, X.-H. Edwardsiella tarda: An intringuing problem in aquaculture. Aquaculture 2014, 431, 129–135. [Google Scholar] [CrossRef]
- Reichley, S.R.; Ware, C.; Steadman, J.; Gaunt, P.S.; García, J.C.; LaFrentz, B.R.; Thachil, A.; Waldbieser, G.C.; Stine, C.B.; Arias, C.R.; et al. Phenotypic and genotypic analysis of Edwardsiella spp. isolates from different hosts and geographical origins, with an emphasis on isolates formerly classified as E. tarda and an evaluation of diagnostic methods. J. Clin. Microbiol. 2017, 55, 3466–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Damme, L.R.; Vandepitte, J. Frequent isolation of Edwardsiella tarda and Plesiomonas shigelloides from healthy Zairese freshwater fish: A possible source of sporadic diarrhea in the Tropics. Appl. Environ. Microbiol. 1980, 39, 475–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sechter, I.; Shmilovitz, M.; Altman, G.; Seligmann, R.; Kretzer, B.; Braunstein, I.; Gerichter, B. Edwardsiella tarda isolated in Israel between 1961 and 1980. J. Clin. Microbiol. 1983, 17, 669–671. [Google Scholar] [CrossRef] [Green Version]
- White, F.H.; Simpson, C.F.; Williams, I.E. Isolation of Edwardsiella tarda from aquatic animal species and surface waters in Florida. J. Wildl. Dis. 1973, 9, 204–208. [Google Scholar] [CrossRef]
- Baya, A.M.; Romalde, J.L.; Green, D.E.; Navarro, R.B.; Evans, J.; May, E.B.; Toranzo, A.E. Edwardsiellosis in wild striped bass from the Cheseapeake Bay. J. Wildl. Dis. 1997, 33, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Esteve, C.; Alcaide, E. Seasonal recovery of Edwardsiella piscicida from wild European eels and natural waters: Isolation methods, virulence and reservoirs. J. Fish. Dis. 2018, 41, 1613–1623. [Google Scholar] [CrossRef]
- Kelly, E.; Martin, P.A.J.; Gibson-Kueh, S.; Morgan, D.L.; Ebner, B.C.; Donalson, J.; Buller, N.; Crook, D.A.; Davis, A.M.; Hammer, M.P.; et al. First detection of Edwardsiella ictaluri (Proteobacteria: Enterobacteriaceae) in wild Australian catfish. J. Fish. Dis. 2018, 41, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Esteve, C.; Alcaide, E. Influence of diseases on the wild eel stock: The case of L’Albufera Lake. Aquaculture 2009, 289, 143–149. [Google Scholar] [CrossRef]
- Katharios, P.; Kokkari, C.; Dourala, N.; Smyrli, M. First report of Edwardsiellosis in cage-cultured sharpsnout sea bream, Diplodus puntazzo from the Mediterranean. BMC Vet. Res. 2015, 11, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camus, A.; Griffin, M.; Armwood, A.; Soto, E. A spontaneous outbreak of systemic Edwardsiella piscicida infection in largemounth bass Micropterus salmoides (Lacépède, 1802) in California, USA. J. Fish. Dis. 2019, 42, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Chen, J.; Zhang, X.; Li, A.; Li, Y.; Wang, Y. Retention of virulence in a viable but nonculturable Edwardsiella tarda isolate. Appl. Environ. Microbiol. 2007, 73, 1349–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biosca, E.G.; Amaro, C.; Marco-Noales, E.; Oliver, J.D. Effect of low temperature on starvation-survival of the eel pathogen Vibrio vulnificus biotype 2. Appl. Environ. Microbiol. 1996, 62, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Cai, W.; Tao, Z.; Arias, C.R. Survival of fish-pathogenic strains of Aeromonas hydrophila under starvation. J. Aquat. Anim. Health 2014, 26, 190–193. [Google Scholar] [CrossRef]
- Duodu, S.; Colquhoun, D. Monitoring the survival of fish-pathogenic Francisella in water microcosms. FEMS Microbiol. Ecol. 2010, 74, 534–541. [Google Scholar] [CrossRef] [Green Version]
- Alcaide, E.; Herraiz, S.; Esteve, C. Occurrence of Edwardsiella tarda in wild European eels Anguilla anguilla from Mediterranean Spain. Dis. Aquat. Org. 2006, 73, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Whitesides, M.D.; Oliver, J.D. Resuscitation of Vibrio vulnificus from the viable but not culturable state. Appl. Environ. Microbiol. 1997, 63, 1002–1005. [Google Scholar] [CrossRef] [Green Version]
- Official Journal of European Union. Directive 2010/63/EU of the European Parliament and of the Council, of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. 2010. Available online: https://ec.europa.eu/environment/chemicals/lab_animals/legislation_en.htm (accessed on 15 May 2022).
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbio. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Schink, S.J.; Biselli, E.; Ammar, C.; Gerland, U. Death rate of E. coli during starvation is set by maintenance cost and biomass recycling. Cell Syst. 2019, 9, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Azevedo, N.F.; Ivask, A. Propidium iodide staining underestimates viability of adherent bacterial cells. Sci. Rep. 2019, 9, 6483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, D.; Santos, M.A.; Chambel, L. Thirty years of viable but nonculturable state research: Unsolved molecular mechanisms. Crit. Rev. Microbiol. 2015, 41, 61–76. [Google Scholar] [CrossRef]
- Abayneh, T.; Colquhoun, D.J.; Sørum, H. Multi-locus sequence snalysis (MLSA) of Edwardsiella tarda isolates from fish. Vet. Microbiol. 2012, 158, 367–375. [Google Scholar] [CrossRef]
- Griffin, M.J.; Quiniou, S.M.; Cody, T.; Tabuchi, M.; Ware, C.; Cipriano, R.C.; Mauel, M.J.; Soto, E. Comparative analysis of Edwardsiella isolates from fish in the eastern United States identifies two distinct genetic taxa amongst organisms phenotypically classified as E. tarda. Vet. Microbiol. 2013, 30, 358–372. [Google Scholar] [CrossRef] [Green Version]
- Abayneh, T.; Colquhoun, D.J.; Sørum, H. Edwardsiella piscicida sp. nov., a novel species pathogenic to fish. J. Appl. Microbiol. 2012, 114, 644–654. [Google Scholar] [CrossRef]
- Shao, S.; Lai, Q.; Liu, Q.; Wu, H.; Xiao, J.; Shao, Z.; Wang, Q.; Zhang, Y. Phylogenomics characterization of a highly virulent Edwardsiella strain ET080813T encoding two distinct T3SS and three gene clusters: Propose a novel species as Edwardsiella anguillarum sp. nov. Syst. Appl. Microbiol. 2015, 38, 36–47. [Google Scholar] [CrossRef]
- Buján, N.; Mohammed, H.; Balboa, S.; Romalde, J.L.; Toranzo, A.E.; Arias, C.R.; Magariños, B. Genetic studies to re-affiliate Edwardsiella tarda fish isolates to Edwardsiella piscicida and Edwardsiella anguillarum species. Syst. Appl. Microbiol. 2018, 41, 30–37. [Google Scholar] [CrossRef]
- Leung, K.Y.; Wang, Q.; Zheng, X.; Zhuang, M.; Yang, Z.; Shao, S.; Achmon, Y.; Siame, B.A. Versatile lifestyles of Edwardsiella: Free-living, pathogen, and core bacterium of the aquatic resistome. Virulence 2022, 13, 5–18. [Google Scholar] [CrossRef]
- Sun, Y.; Li, Y.; Luo, Q.; Huang, J.; Chen, J.; Zhang, R.; Wang, X. LuS/AI-2 Quorum sensing system in Edwardsiella piscicida promores biofilm formation and pathogenicity. Infect. Immun. 2020, 88, e00907–e00919. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.A.; Ding, X.; Zhang, X.; Zhu, G. Fish borne Edwardsiella tarda eha involved in the bacterial biofilm formation, hemolytic activity, adhesion capability and pathogenicity. Arch. Microbiol. 2020, 202, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.Y.; Siame, B.A.; Tenkink, B.J.; Noort, R.J.; Mok, Y.-K. Edwardsiella tarda—Virulence mechanisms of an emerging gastroenteritis pathogen. Microb. Infect. 2012, 14, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-S.; Chowdhury, N.; Yamasaki, R.; Wood, T.K. Viable but non-culturable and persistence describe the same bacterial stress state. Environ. Microbiol. 2018, 20, 2038–2048. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, C.Y. Starvation induces phenotypic diversification and convergent evolution in Vibrio vulnificus. PLoS ONE 2014, 9, e88658. [Google Scholar] [CrossRef] [Green Version]
- Jubair, M.; Morris, G.J.J.; Ali, A. Survival of Vibrio cholera in nutrient-poor environments is associated with a novel “persister” phenotype. PLoS ONE 2012, 7, e45187. [Google Scholar] [CrossRef]
- Jubair, M.; Atanasova, K.R.; Rahman, M.; Klose, K.E.; Yasmin, M.; Yilmaz, Ö.; Morris, J.G., Jr.; Ali, A. Vibrio cholera persisted in microcosm for 700 days inhibits motility but promotes biofilm formation in nutrient-poor lake water microcosms. PLoS ONE 2014, 9, e92883. [Google Scholar] [CrossRef] [Green Version]
- Senoh, M.; Ghosh-Banerjee, J.; Ramamurthy, T.; Hamabata, T.; Kurakawa, T.; Takeda, M.; Colwell, R.R.; Balakrish Nair, G.; Takeda, Y. Conversion of viable but non culturable Vibrio cholera to the culturable state by co-culture with eukaryotic cells. Microbiol. Immunol. 2010, 54, 502–507. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | IP Challenges Dose (CFU/g of Fish) | 5 Days-Accumulative Mortality |
|---|---|---|
| Edwardsiella piscicida | ||
| V12.1 original | 1.20 × 106 | 83% |
| V12.1_7 °C_R_1 w 1 | 0.64 × 106 | 100% |
| V12.1_15 °C_R_2 w | 1.00 × 106 | 80% |
| V12.1_25 °C_R_2 w | 1.00 × 106 | 67% |
| Edwardsiella piscicida | ||
| V57.2 original | 2.30 × 105 | 80% |
| V57.2_7 °C_R_1 w | 1.42 × 106 | 100% |
| V57.2_15 °C_R_2 w | 0.70 × 106 | 80% |
| Edwardsiella tarda | ||
| V43.2 original | 1.10 × 107 | 0% |
| V43.2_7 °C_R_1 w | 2.16 × 107 | 0% |
| V43.2_15 °C_R_1 w | 2.42 × 107 | 0% |
| V43.2_25 °C_R_2 w | 1.86 × 107 | 0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esteve, C.; Alcaide, E. Monitoring the Starvation–Survival Response of Edwardsiella piscicida and E. tarda in Freshwater Microcosms, at Various Temperatures. Microorganisms 2022, 10, 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051043
Esteve C, Alcaide E. Monitoring the Starvation–Survival Response of Edwardsiella piscicida and E. tarda in Freshwater Microcosms, at Various Temperatures. Microorganisms. 2022; 10(5):1043. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051043
Chicago/Turabian StyleEsteve, Consuelo, and Elena Alcaide. 2022. "Monitoring the Starvation–Survival Response of Edwardsiella piscicida and E. tarda in Freshwater Microcosms, at Various Temperatures" Microorganisms 10, no. 5: 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051043