
Comparison of the Microbiome of Artisanal Homemade and Industrial Feta Cheese through Amplicon Sequencing and Shotgun Metagenomics

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cheese Samples
2.2. DNA Extraction
2.3. Sequencing and Bioinformatics Analysis
3. Results and Discussion
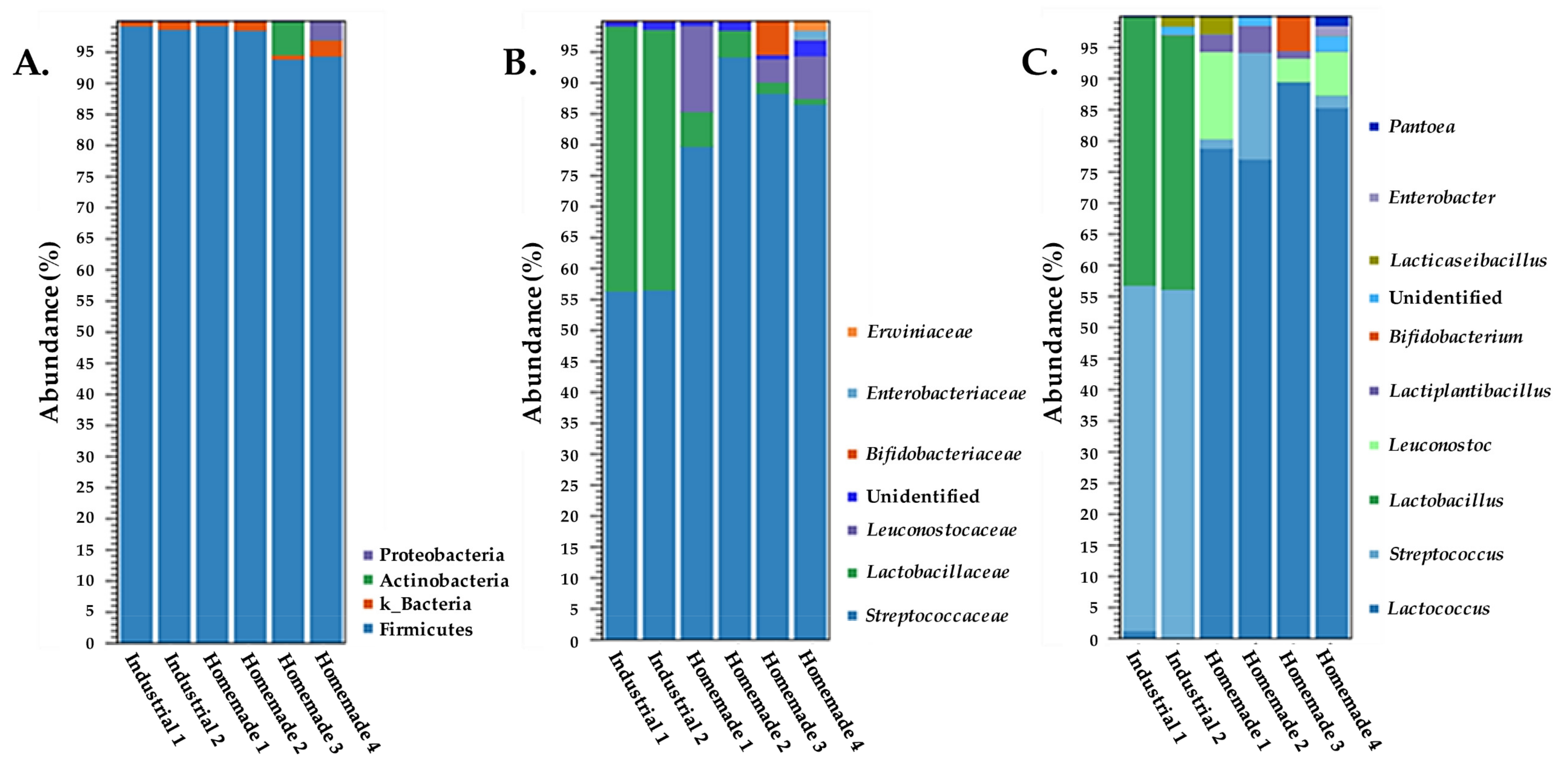
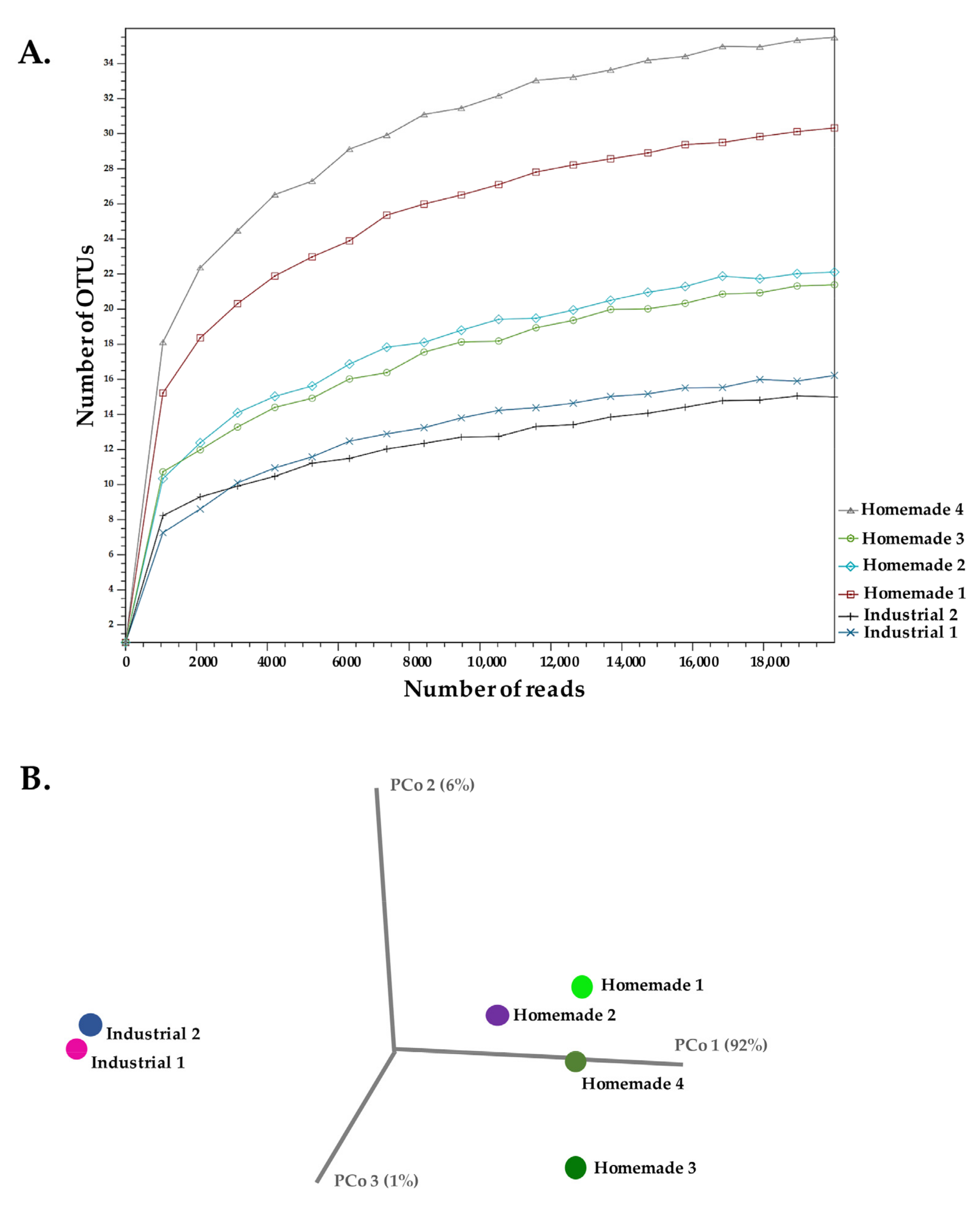
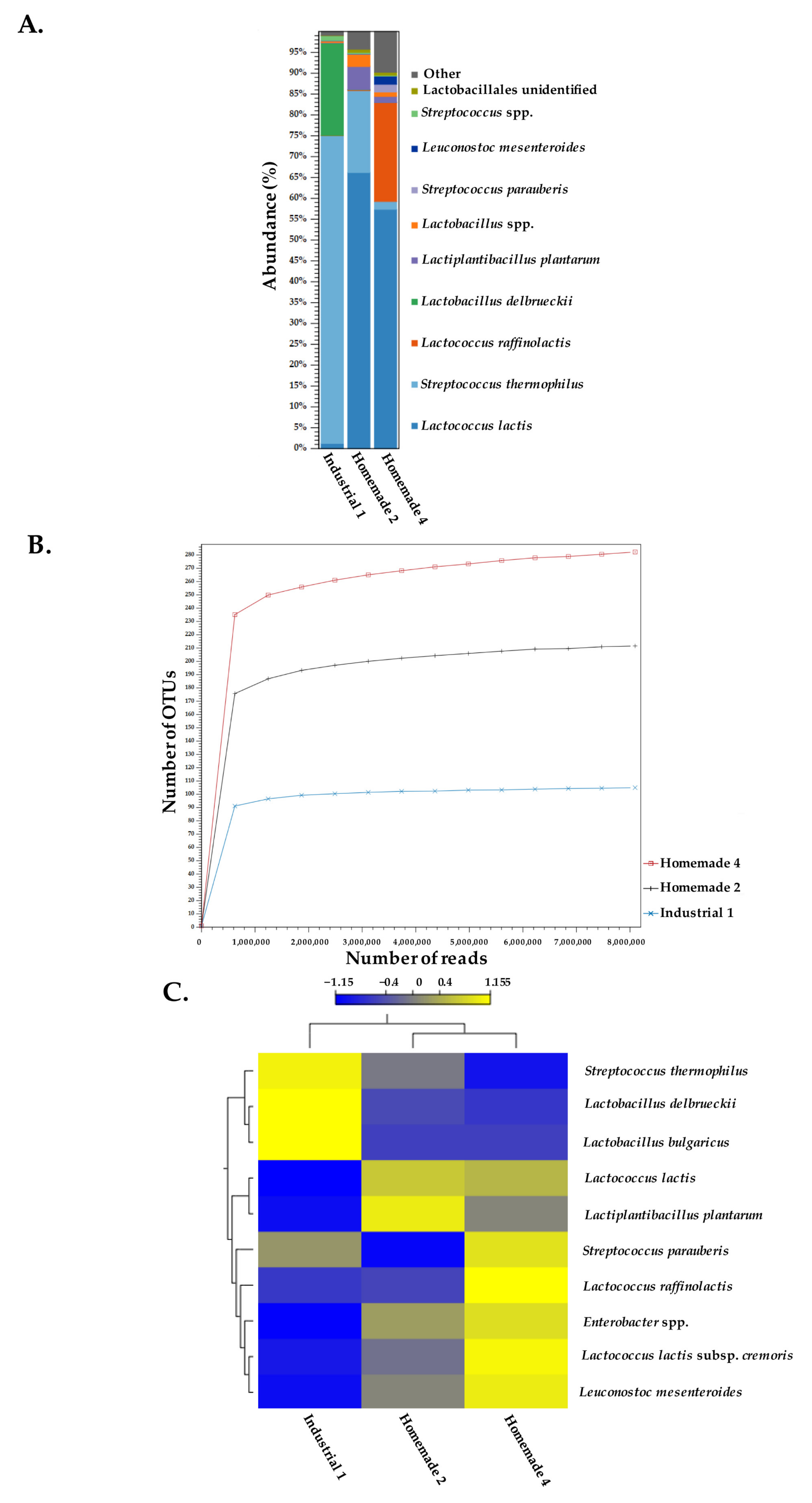
3.1. 16S rDNA Amplicon Sequencing
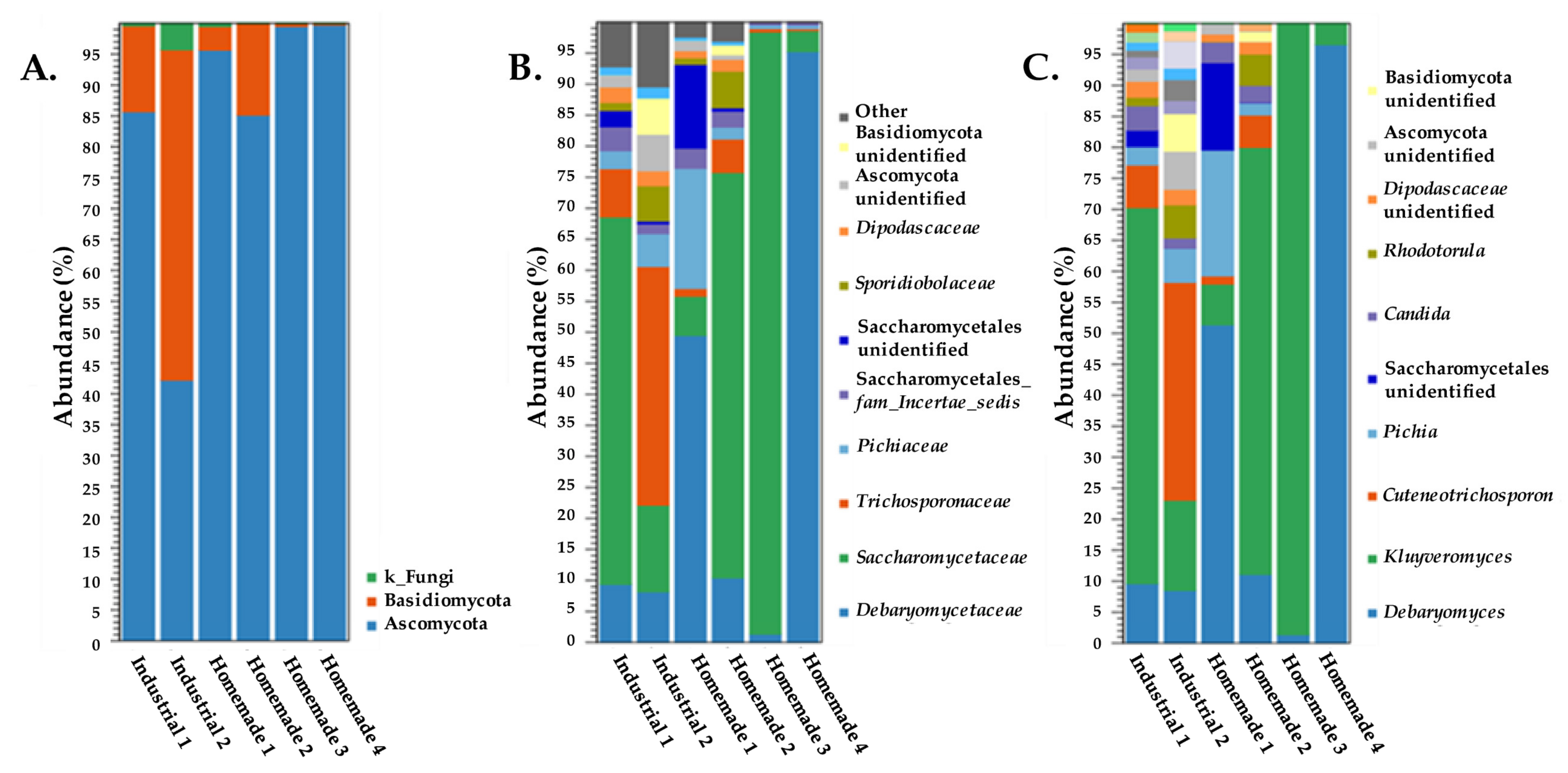
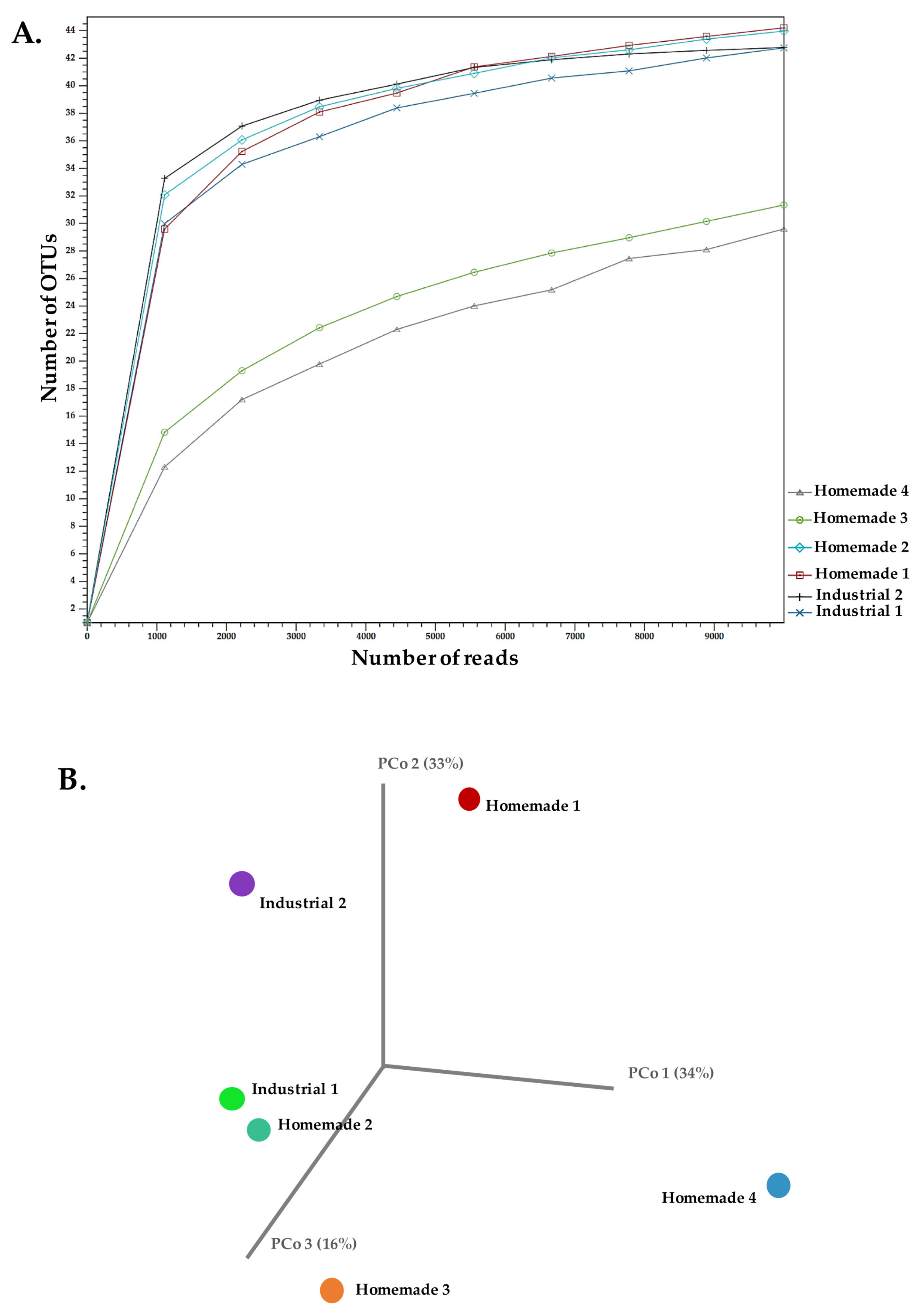
3.2. ITS Amplicon Sequencing
3.3. Species Identification Using Shotgun Metagenomic Sequence Reads of Feta Cheese Samples
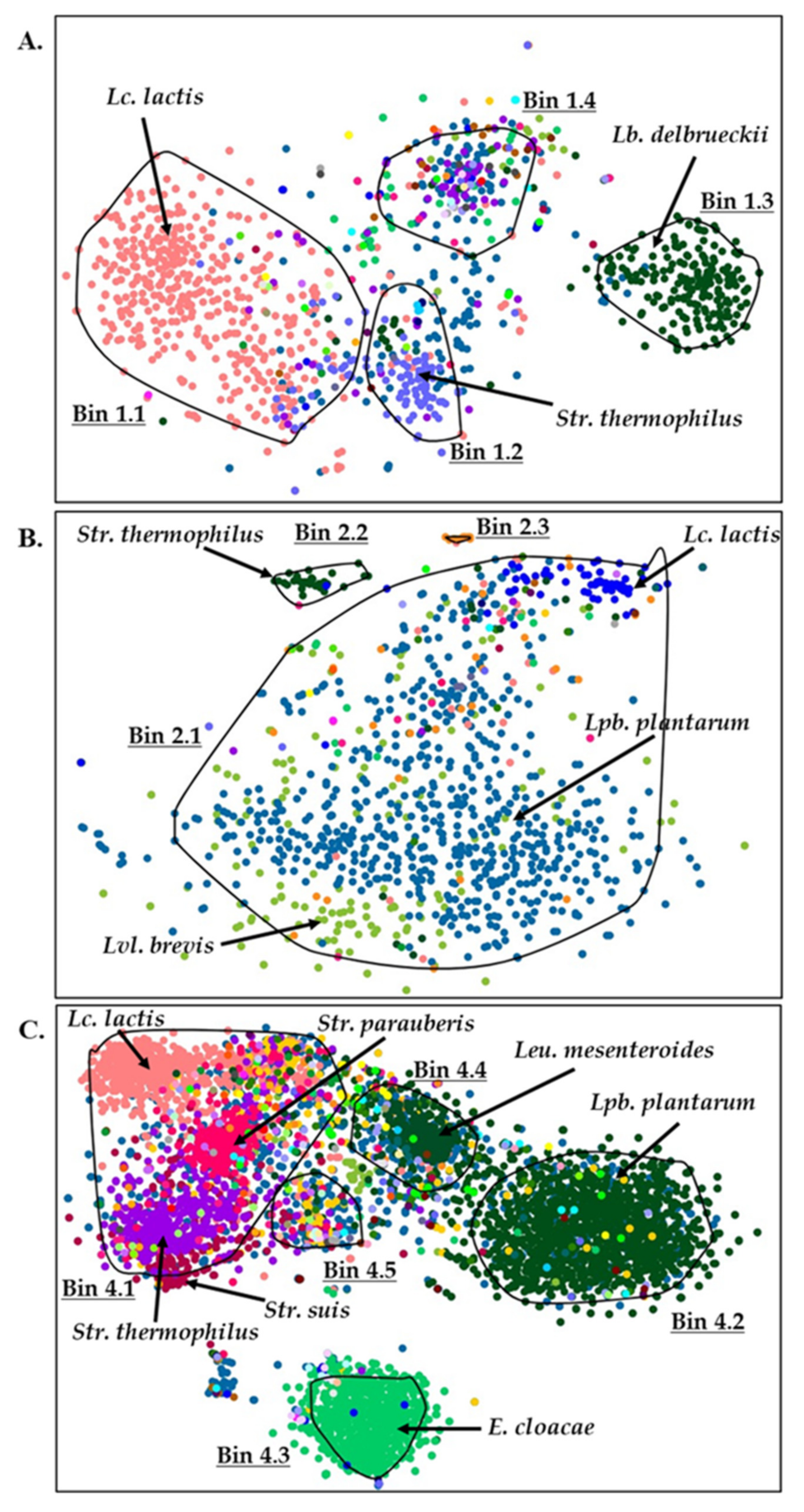
3.4. Binning Metagenomic Scaffolds of Feta Cheese Samples
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Manolopoulou, E.; Sarantinopoulos, P.; Zoidou, E.; Aktypis, A.; Moschopoulou, E.; Kandarakis, I.G.; Anifantakis, E.M. Evolution of microbial populations during traditional Feta cheese manufacture and ripening. Int. J. Food Microbiol. 2003, 82, 153–161. [Google Scholar] [CrossRef]
- Tzanetakis, N.; Litopoulou-Tzanetaki, E. Changes in numbers and kinds of lactic acid bacteria in Feta and Teleme, two greek cheeses from ewe’s milk. J. Dairy Sci. 1992, 75, 1389–1393. [Google Scholar] [CrossRef]
- Anifantakis, E.M.; Moatsou, G. Feta and other Balkan cheeses. In Brined Cheeses; Tamime, A.Y., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 43–76. [Google Scholar] [CrossRef]
- Bintsis, T.; Papademas, P. Microbiological quality of white-brined cheeses: A review. Int. J. Dairy Technol. 2002, 55, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Pappas, C.P.; Kondyli, E.; Voutsinas, L.P.; Mallatou, H. Effects of starter level, draining time and aging on the physicochemical, organoleptic and rheological properties of Feta cheese. Int. J. Dairy Technol. 1996, 49, 73–78. [Google Scholar] [CrossRef]
- Kandarakis, I.; Moatsou, G.; Georgala, A.I.K.; Kaminarides, S.; Anifantakis, E. Effect of draining temperature on the biochemical characteristics of Feta cheese. Food Chem. 2001, 72, 369–378. [Google Scholar] [CrossRef]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N.; Vafopoulou-Mastrojiannaki, A. Effect of the type of lactic starter on microbiologicalchemical and sensory characteristics of Feta cheese. Food Microbiol. 1993, 10, 31–41. [Google Scholar] [CrossRef]
- Pappa, H.C.; Anyfantakis, E.M. Effect of concentrated starter cultures on the manufacture of Feta cheese. Milchwissenschaft 2001, 56, 325–329. [Google Scholar]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N. The microfloras of traditional Greek cheeses. Microbiol. Spectr. 2014, 2, Cm-0009-2012. [Google Scholar] [CrossRef] [Green Version]
- Özer, B.H. Microflora of white-brined cheeses. In Encyclopedia of Food Microbiology; Robinson, R.K., Ed.; Elsevier: Oxford, UK, 1999; pp. 397–403. [Google Scholar]
- Pappas, C.P.; Kondyli, E.; Voutsinas, L.P.; Mallatou, H. Effects of salting method and storage time on composition and quality of Feta cheese. Int. J. Dairy Technol. 1996, 49, 113–118. [Google Scholar] [CrossRef]
- Bintsis, T.; Litopoulou-Tzanetaki, E.; Davies, R.; Robinson, R.K. Microbiology of brines used to mature Feta cheese. Int. J. Dairy Technol. 2000, 53, 106–112. [Google Scholar] [CrossRef]
- Rantsiou, K.; Urso, R.; Dolci, P.; Comi, G.; Cocolin, L. Microflora of Feta cheese from four Greek manufacturers. Int. J. Food Microbiol. 2008, 126, 36–42. [Google Scholar] [CrossRef]
- Fadda, M.E.; Cosentino, S.; Deplano, M.; Palmas, F. Yeast populations in Sardinian feta cheese. Int. J. Food Microbiol. 2001, 69, 153–156. [Google Scholar] [CrossRef]
- Kaminarides, S.E.; Laskos, N.S. Yeasts in factory brine of Feta cheese. Aust. J. Dairy Technol. 1992, 47, 68–71. [Google Scholar]
- Psomas, E.; Andrighetto, C.; Litopoulou-Tzanetaki, E.; Lombardi, A.; Tzanetakis, N. Some probiotic properties of yeast isolates from infant faeces and Feta cheese. Int. J. Food Microbiol. 2001, 69, 125–133. [Google Scholar] [CrossRef]
- Westall, S.; Filtenborg, O. Yeast occurrence in Danish feta cheese. Food Microbiol. 1998, 15, 215–222. [Google Scholar] [CrossRef]
- Bintsis, T.; Robinson, R.K. A study of the effects of adjunct cultures on the aroma compounds of Feta-type cheese. Food Chem. 2004, 88, 435–441. [Google Scholar] [CrossRef]
- Sarantinopoulos, P.; Kalantzopoulos, G.; Tsakalidou, E. Effect of Enterococcus faecium on microbiological, physicochemical and sensory characteristics of Greek Feta cheese. Int. J. Food Microbiol. 2002, 76, 93–105. [Google Scholar] [CrossRef]
- Angelopoulou, A.; Alexandraki, V.; Georgalaki, M.; Anastasiou, R.; Manolopoulou, E.; Tsakalidou, E.; Papadimitriou, K. Production of probiotic Feta cheese using Propionibacterium freudenreichii subsp. shermanii as adjunct. Int. Dairy J. 2017, 66, 135–139. [Google Scholar] [CrossRef]
- Dimitrellou, D.; Kandylis, P.; Sidira, M.; Koutinas, A.A.; Kourkoutas, Y. Free and immobilized Lactobacillus casei ATCC 393 on whey protein as starter cultures for probiotic Feta-type cheese production. J. Dairy Sci. 2014, 97, 4675–4685. [Google Scholar] [CrossRef]
- Mantzourani, I.; Terpou, A.; Alexopoulos, A.; Chondrou, P.; Galanis, A.; Bekatorou, A.; Bezirtzoglou, E.; Koutinas, A.A.; Plessas, S. Application of a novel potential probiotic Lactobacillus paracasei strain isolated from kefir grains in the production of feta-type cheese. Microorganisms 2018, 6, 121. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, O.S.; Argyri, A.A.; Varzakis, E.E.; Tassou, C.C.; Chorianopoulos, N.G. Greek functional Feta cheese: Enhancing quality and safety using a Lactobacillus plantarum strain with probiotic potential. Food Microbiol. 2018, 74, 21–33. [Google Scholar] [CrossRef]
- Terpou, A.; Mantzourani, I.; Galanis, A.; Kanellaki, M.; Bezirtzoglou, E.; Bekatorou, A.; Koutinas, A.A.; Plessas, S. Employment of L. paracasei K5 as a novel potentially probiotic freeze-dried starter for Feta-type cheese production. Microorganisms 2018, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Vafopoulou-Mastrojiannaki, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Effect of Pediococcus pentosaceus on ripening changes of Feta cheese. Microbiol. Aliment. Nutr. 1990, 8, 53–62. [Google Scholar]
- Afshari, R.; Pillidge, C.J.; Dias, D.A.; Osborn, A.M.; Gill, H. Cheesomics: The future pathway to understanding cheese flavour and quality. Crit. Rev. Food Sci. Nutr. 2020, 60, 33–47. [Google Scholar] [CrossRef]
- Cao, Y.; Fanning, S.; Proos, S.; Jordan, K.; Srikumar, S. A Review on the Applications of Next Generation Sequencing Technologies as Applied to Food-Related Microbiome Studies. Front. Microbiol. 2017, 8, 1829. [Google Scholar] [CrossRef]
- De Filippis, F.; Parente, E.; Ercolini, D. Metagenomics insights into food fermentations. Microb. Biotechnol. 2017, 10, 91–102. [Google Scholar] [CrossRef]
- Kamilari, E.; Tomazou, M.; Antoniades, A.; Tsaltas, D. High throughput sequencing technologies as a new toolbox for deep analysis, characterization and potentially authentication of protection designation of origin cheeses? Int. J. Food Sci. 2019, 2019, 5837301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kergourlay, G.; Taminiau, B.; Daube, G.; Champomier Vergès, M.-C. Metagenomic insights into the dynamics of microbial communities in food. Int. J. Food Microbiol. 2015, 213, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Yeluri Jonnala, B.R.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Sequencing of the cheese microbiome and its relevance to industry. Front. Microbiol. 2018, 9, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michailidou, S.; Pavlou, E.; Pasentsis, K.; Rhoades, J.; Likotrafiti, E.; Argiriou, A. Microbial profiles of Greek PDO cheeses assessed with amplicon metabarcoding. Food Microbiol. 2021, 99, 103836. [Google Scholar] [CrossRef] [PubMed]
- Papadakis, P.; Konteles, S.; Batrinou, A.; Ouzounis, S.; Tsironi, T.; Halvatsiotis, P.; Tsakali, E.; Van Impe, J.F.M.; Vougiouklaki, D.; Strati, I.F.; et al. Characterization of bacterial microbiota of P.D.O. Feta cheese by 16S metagenomic analysis. Microorganisms 2021, 9, 2377. [Google Scholar] [CrossRef]
- Spyrelli, E.D.; Stamatiou, A.; Tassou, C.C.; Nychas, G.-J.E.; Doulgeraki, A.I. Microbiological and metagenomic analysis to assess the effect of container material on the microbiota of Feta cheese during ripening. Fermentation 2020, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Bozoudi, D.; Kotzamanidis, C.; Hatzikamari, M.; Tzanetakis, N.; Menexes, G.; Litopoulou-Tzanetaki, E. A comparison for acid production, proteolysis, autolysis and inhibitory properties of lactic acid bacteria from fresh and mature Feta PDO Greek cheese, made at three different mountainous areas. Int. J. Food Microbiol. 2015, 200, 87–96. [Google Scholar] [CrossRef]
- Bozoudi, D.; Torriani, S.; Zdragas, A.; Litopoulou-Tzanetaki, E. Assessment of microbial diversity of the dominant microbiota in fresh and mature PDO Feta cheese made at three mountainous areas of Greece. LWT Food Sci. Technol. 2016, 72, 525–533. [Google Scholar] [CrossRef]
- Pitcher, D.G.; Saunders, N.A.; Owen, R.J. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol. 1989, 8, 151–156. [Google Scholar] [CrossRef]
- Delforno, T.P.; Belgini, D.R.B.; Hidalgo, K.J.; Centurion, V.B.; Lacerda-Júnior, G.V.; Duarte, I.C.S.; Varesche, M.B.A.; Oliveira, V.M. Anaerobic reactor applied to laundry wastewater treatment: Unveiling the microbial community by gene and genome-centric approaches. Int. Biodeterior. Biodegrad. 2020, 149, 104916. [Google Scholar] [CrossRef]
- Papademas, P.; Aspri, M.; Mariou, M.; Dowd, S.E.; Kazou, M.; Tsakalidou, E. Conventional and omics approaches shed light on Halitzia cheese, a long-forgotten white-brined cheese from Cyprus. Int. Dairy J. 2019, 98, 72–83. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef]
- Laczny, C.C.; Kiefer, C.; Galata, V.; Fehlmann, T.; Backes, C.; Keller, A. BusyBee Web: Metagenomic data analysis by bootstrapped supervised binning and annotation. Nucleic Acids Res. 2017, 45, W171–W179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, F.; Bagchi, S.; Chaterji, S.; Gerlach, W.; Grama, A.; Harrison, T.; Paczian, T.; Trimble, W.L.; Wilke, A. MG-RAST version 4—lessons learned from a decade of low-budget ultra-high-throughput metagenome analysis. Brief. Bioinform. 2019, 20, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Geronikou, A.; Srimahaeak, T.; Rantsiou, K.; Triantafillidis, G.; Larsen, N.; Jespersen, L. Occurrence of yeasts in white-brined cheeses: Methodologies for identification, spoilage potential and good manufacturing practices. Front. Microbiol. 2020, 11, 2454. [Google Scholar] [CrossRef] [PubMed]
- Genigeorgis, C.; Carniciu, M.; Dutulescu, D.; Farver, T.B. Growth and survival of Listeria monocytogenes in market cheeses stored at 4 to 30 °C. J. Food Prot. 1991, 54, 662–668. [Google Scholar] [CrossRef]
- Papageorgiou, D.K.; Marth, E.H. Fate of Listeria monocytogenes during the manufacture, ripening and storage of Feta cheese. J. Food Prot. 1989, 52, 82–87. [Google Scholar] [CrossRef]
- Govaris, A.; Papageorgiou, D.K.; Papatheodorou, K. Behavior of Escherichia coli O157:H7 during the manufacture and ripening of Feta and Telemes cheeses. J. Food Prot. 2002, 65, 609–615. [Google Scholar] [CrossRef]
- Pexara, A.; Solomakos, N.; Sergelidis, D.; Govaris, A. Fate of enterotoxigenic Staphylococcus aureus and staphylococcal enterotoxins in Feta and Galotyri cheeses. J. Dairy Res. 2012, 79, 405–413. [Google Scholar] [CrossRef]
- Erkmen, O. Survival of virulent Yersinia enterocolitica during the manufacture and storage of Turkish Feta cheese. Int. J. Food Microbiol. 1996, 33, 285–292. [Google Scholar] [CrossRef]
- Maipa, V.; Papadopoulou, C.; Pappas, C.; Dimitriou, D.; Voutsinas, L.; Malatou, H. Survival of Salmonella enteritidis during the manufacture of Feta cheese made of pasteurized ewe’s milk. Zent. Mikrobiol. 1993, 148, 66–73. [Google Scholar] [CrossRef]
- Riffon, R.; Sayasith, K.; Khalil, H.; Dubreuil, P.; Drolet, M.; Lagacé, J. Development of a rapid and sensitive test for identification of major pathogens in bovine mastitis by PCR. J. Clin. Microbiol. 2001, 39, 2584–2589. [Google Scholar] [CrossRef] [Green Version]
- Davin-Regli, A.; Pagès, J.-M. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Front. Microbiol. 2015, 6, 392. [Google Scholar] [CrossRef] [Green Version]
- Gaya, P.; Medina, M.; Nuñez, M. Accelerated decrease of Enterobacteriaceae counts during ripening of raw milk Manchego cheese by lactic culture inoculation. J. Food Prot. 1983, 46, 305–308. [Google Scholar] [CrossRef]
- Maifreni, M.; Frigo, F.; Bartolomeoli, I.; Innocente, N.; Biasutti, M.; Marino, M. Identification of the Enterobacteriaceae in Montasio cheese and assessment of their amino acid decarboxylase activity. J. Dairy Res. 2013, 80, 122–127. [Google Scholar] [CrossRef]
- Morales, P.; Feliu, I.; Fernández-García, E.; Nuñez, M. Volatile compounds produced in cheese by Enterobacteriaceae strains of dairy origin. J. Food Prot. 2004, 67, 567–573. [Google Scholar] [CrossRef]
- Tornadijo, M.E.; García, M.C.; Fresno, J.M.; Carballo, J. Study of Enterobacteriaceae during the manufacture and ripening of San Simón cheese. Food Microbiol. 2001, 18, 499–509. [Google Scholar] [CrossRef]
- De Filippis, F.; La Storia, A.; Stellato, G.; Gatti, M.; Ercolini, D. A selected core microbiome drives the early stages of three popular italian cheese manufactures. PLoS ONE 2014, 9, e89680. [Google Scholar] [CrossRef]
- Fortina, M.G.; Ricci, G.; Acquati, A.; Zeppa, G.; Gandini, A.; Manachini, P.L. Genetic characterization of some lactic acid bacteria occurring in an artisanal protected denomination origin (PDO) Italian cheese, the Toma piemontese. Food Microbiol. 2003, 20, 397–404. [Google Scholar] [CrossRef]
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus suis: An emerging zoonotic pathogen. Lancet Infect. Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef]
- de Garnica, M.L.; Sáez-Nieto, J.A.; González, R.; Santos, J.A.; Gonzalo, C. Diversity of gram-positive catalase-negative cocci in sheep bulk tank milk by comparative 16S rDNA sequence analysis. Int. Dairy J. 2014, 34, 142–145. [Google Scholar] [CrossRef]
- Zhang, R.; Huo, W.; Zhu, W.; Mao, S. Characterization of bacterial community of raw milk from dairy cows during subacute ruminal acidosis challenge by high-throughput sequencing. J. Sci. Food Agric. 2015, 95, 1072–1079. [Google Scholar] [CrossRef]
- Pirisi, A.; Comunian, R.; Urgeghe, P.P.; Scintu, M.F. Sheep’s and goat’s dairy products in Italy: Technological, chemical, microbiological, and sensory aspects. Small Rumin. Res. 2011, 101, 102–112. [Google Scholar] [CrossRef]
- Porcellato, D.; Skeie, S.B. Bacterial dynamics and functional analysis of microbial metagenomes during ripening of Dutch-type cheese. Int. Dairy J. 2016, 61, 182–188. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Button, J.E.; Santarelli, M.; Dutton, R.J. Cheese rind communities provide tractable systems for in situ and in vitro studies of microbial diversity. Cell 2014, 158, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Nalbantoglu, U.; Cakar, A.; Dogan, H.; Abaci, N.; Ustek, D.; Sayood, K.; Can, H. Metagenomic analysis of the microbial community in kefir grains. Food Microbiol. 2014, 41, 42–51. [Google Scholar] [CrossRef]
- Yasir, M.; Bibi, F.; Hashem, A.M.; Azhar, E.I. Comparative metagenomics and characterization of antimicrobial resistance genes in pasteurized and homemade fermented Arabian laban. Food Res. Int. 2020, 137, 109639. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 16S rDNA Amplicon | ITS Amplicon | |||
|---|---|---|---|---|
| Samples | Number of Reads * | Number of OTUs | Number of Reads * | Number of OTUs |
| Industrial 1 | 34,683 | 19 | 31,811 | 49 |
| Industrial 2 | 32,556 | 19 | 12,026 | 44 |
| Homemade 1 | 22,334 | 33 | 25,127 | 48 |
| Homemade 2 | 24,466 | 26 | 50,218 | 50 |
| Homemade 3 | 37,619 | 26 | 134,137 | 45 |
| Homemade 4 | 25,693 | 41 | 280,150 | 50 |
| Sample | Bin | Completeness * | Contamination * | Strain Heterogeneity * |
|---|---|---|---|---|
| Industrial 1 | 1.1 | 82.0 | 19.8 | 43.5 |
| 1.2 | 90.1 | 6.3 | 14.3 | |
| 1.3 | 95.5 | 1.8 | 50.0 | |
| 1.4 | 93.7 | 18.0 | 20.0 | |
| Homemade 2 | 2.1 | 95.5 | 103.6 | 4.6 |
| 2.2 | 95.5 | 0.9 | 0.0 | |
| 2.3 | 0.0 | 0.0 | 0.0 | |
| Homemade 4 | 4.1 | 95.5 | 232.4 | 21.0 |
| 4.2 | 75.7 | 35.1 | 65.2 | |
| 4.3 | 47.8 | 3.6 | 25.0 | |
| 4.4 | 92.8 | 9.0 | 0.0 | |
| 4.5 | 93.7 | 4.5 | 40.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadimitriou, K.; Anastasiou, R.; Georgalaki, M.; Bounenni, R.; Paximadaki, A.; Charmpi, C.; Alexandraki, V.; Kazou, M.; Tsakalidou, E. Comparison of the Microbiome of Artisanal Homemade and Industrial Feta Cheese through Amplicon Sequencing and Shotgun Metagenomics. Microorganisms 2022, 10, 1073. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051073
Papadimitriou K, Anastasiou R, Georgalaki M, Bounenni R, Paximadaki A, Charmpi C, Alexandraki V, Kazou M, Tsakalidou E. Comparison of the Microbiome of Artisanal Homemade and Industrial Feta Cheese through Amplicon Sequencing and Shotgun Metagenomics. Microorganisms. 2022; 10(5):1073. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051073
Chicago/Turabian StylePapadimitriou, Konstantinos, Rania Anastasiou, Marina Georgalaki, Rimi Bounenni, Argiro Paximadaki, Christina Charmpi, Voula Alexandraki, Maria Kazou, and Effie Tsakalidou. 2022. "Comparison of the Microbiome of Artisanal Homemade and Industrial Feta Cheese through Amplicon Sequencing and Shotgun Metagenomics" Microorganisms 10, no. 5: 1073. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051073