
Generation of PHB from Spent Sulfite Liquor Using Halophilic Microorganisms

 and
and
Abstract
:1. Introduction
- Utilization of SSL by halophilic and extremely halophilic microorganisms.
- Generation of PHB using SSL as feedstock.
- Investigating the effect of common inhibitors in wood hydrolysate on Halophiles.
1.1. Characterization and State of the Art Utilization of SSL from Chemical Pulping
1.2. Biorefinery on SSL

| Sulfite Process | ||
|---|---|---|
| pH | 1–4 | |
| temperature | 80–95 °C | |
| spruce % w/w DM | birch % w/w DM | |
| monosaccharides | ||
| arabinose | 1 | 0 |
| galactose | 5 | 1 |
| glucose | 4 | 1 |
| mannose | 12 | 6 |
| xylose | 6 | 21 |
| organic acids | ||
| acetic acid | 4 | |
| aldonic acids | 5 | |
| glucuronic acid | 2 | |
| others | ||
| lignosulfonate | 55 | |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1.3. Common Substances in Lignocellulose Hydrolysate, Known to Inhibit Microbial Growth
1.4. Halophilic Microorganisms in Biotechnology
2. Experimental Section
2.1. Selection of Strains
2.2. Media
2.3. Cultivation
2.4. Growth Measurement
2.5. HPLC Analysis
2.6. Nile Blue Staining
2.7. SSL
2.8. Inhibiting Substances in SSL
3. Results and Discussion
3.1. Strain Selection
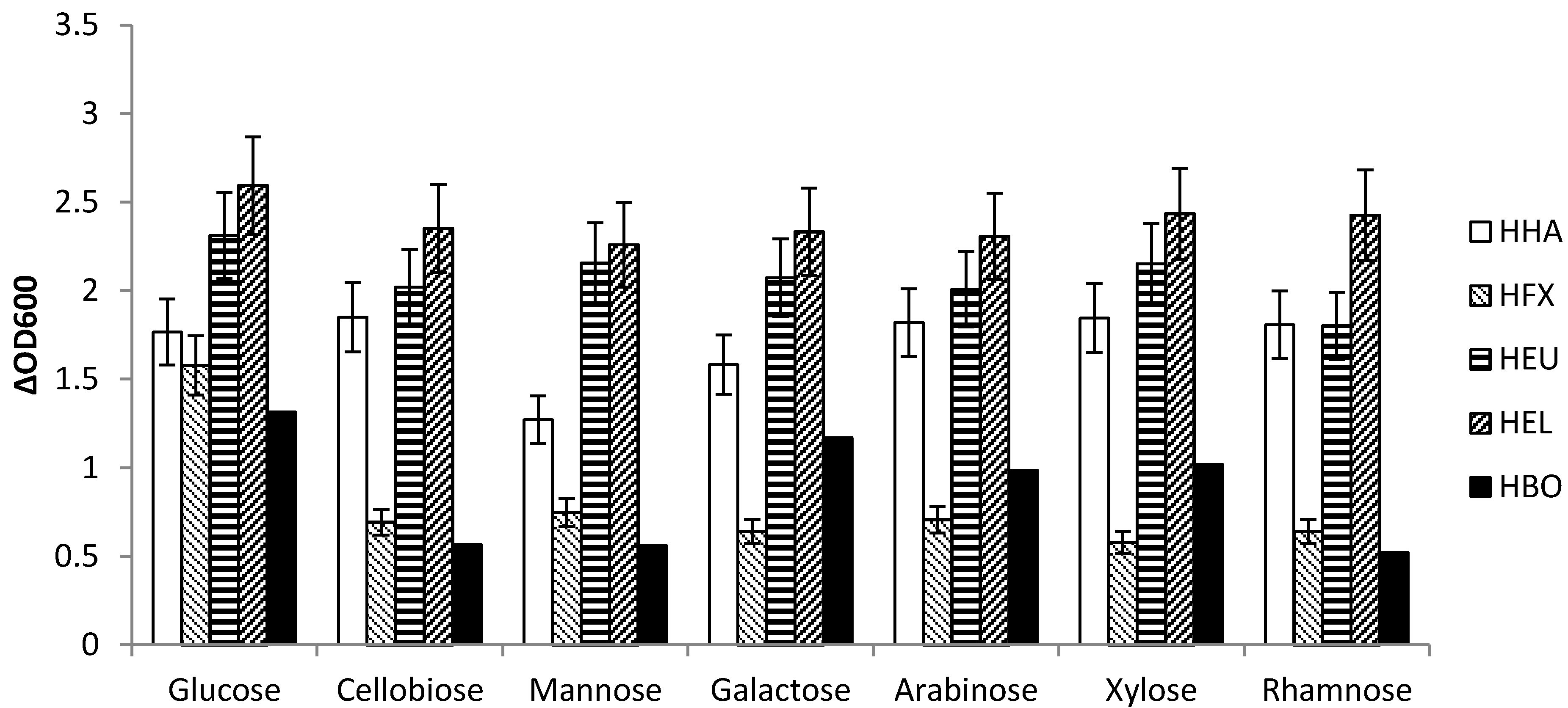
3.2. Utilization of Various Carbon Sources
3.2.1. Utilization of Single Sugars
| Name | Growth Conditions | Ar | Cb | Ga | Gl | Ma | Rh | Xy | Other | Product Yields |
|---|---|---|---|---|---|---|---|---|---|---|
| Halomonas boliviensis DSM 15516T | NaCl 0%–25% w/v, pH 6–11, 0–45 °C | y | y | y | oligo-sacharides Na- acetate, butyric acid | PHB (50%–90% w/w), ectoine (9.2 g·L−1) [35] | ||||
| Halomonas elongata DSM 2581T | NaCl 0%–20% w/v, pH 5–10; 4–45 °C | y | y | y | y | y | y | y | PHB 55% (w/v); ectoine [35,40] | |
| Halomonas eurihalina DSM-5720T | NaCl 0.5%–25% w/v pH 5–10, 4–45 °C | y | y | y | y | y | y | y | PHB [40] | |
| Halomonas halophila DSM 4770 | NaCl 2%–25% w/v, pH 5–10, 4–45 °C | y | y | y | y | y | y | y | PHB [40] | |
| Haloferax mediterranei DSM 1411 | NaCl 20%–25% (w/v) 37 °C | y | y | y | y | glycerol, starch | PHBV (18.21% ± 1.88% wt/wt) [10] |


| Strain | Arabinose | Cellobiose | Galactose | Glucose | Mannose | Rhamnose | Xylose |
|---|---|---|---|---|---|---|---|
| H. boliviensis | N | N | Y | Y | Y | N | Y |
| H. elongata | P | Y | Y | Y | Y | Y | Y |
| H. eurihalina | Y | Y | Y | Y | Y | Y | Y |
| H. halophila | Y | Y | Y | Y | P | Y | Y |
| Strain | μmax [h−1] Arabinose | μmax [h−1] Cellobiose | μmax [h−1] Galactose | μmax [h−1] Glucose | μmax [h−1] Mannose | μmax [h−1] Xylose |
|---|---|---|---|---|---|---|
| H. boliviensis | 0.27 | 0.25 | 0.29 | 0.29 | 0.24 | 0.26 |
| H. elongata | 0.27 | 0.26 | 0.27 | 0.26 | 0.27 | 0.27 |
| H. eurihalina | 0.27 | 0.27 | 0.27 | 0.27 | 0.27 | 0.27 |
| H. halophila | 0.28 | 0.3 | 0.3 | 0.29 | 0.28 | 0.29 |
| H. mediterranei | 0.17 | 0.16 | 0.17 | 0.17 | 0.17 | 0.16 |
3.2.2. Effect of Mixed Substrate



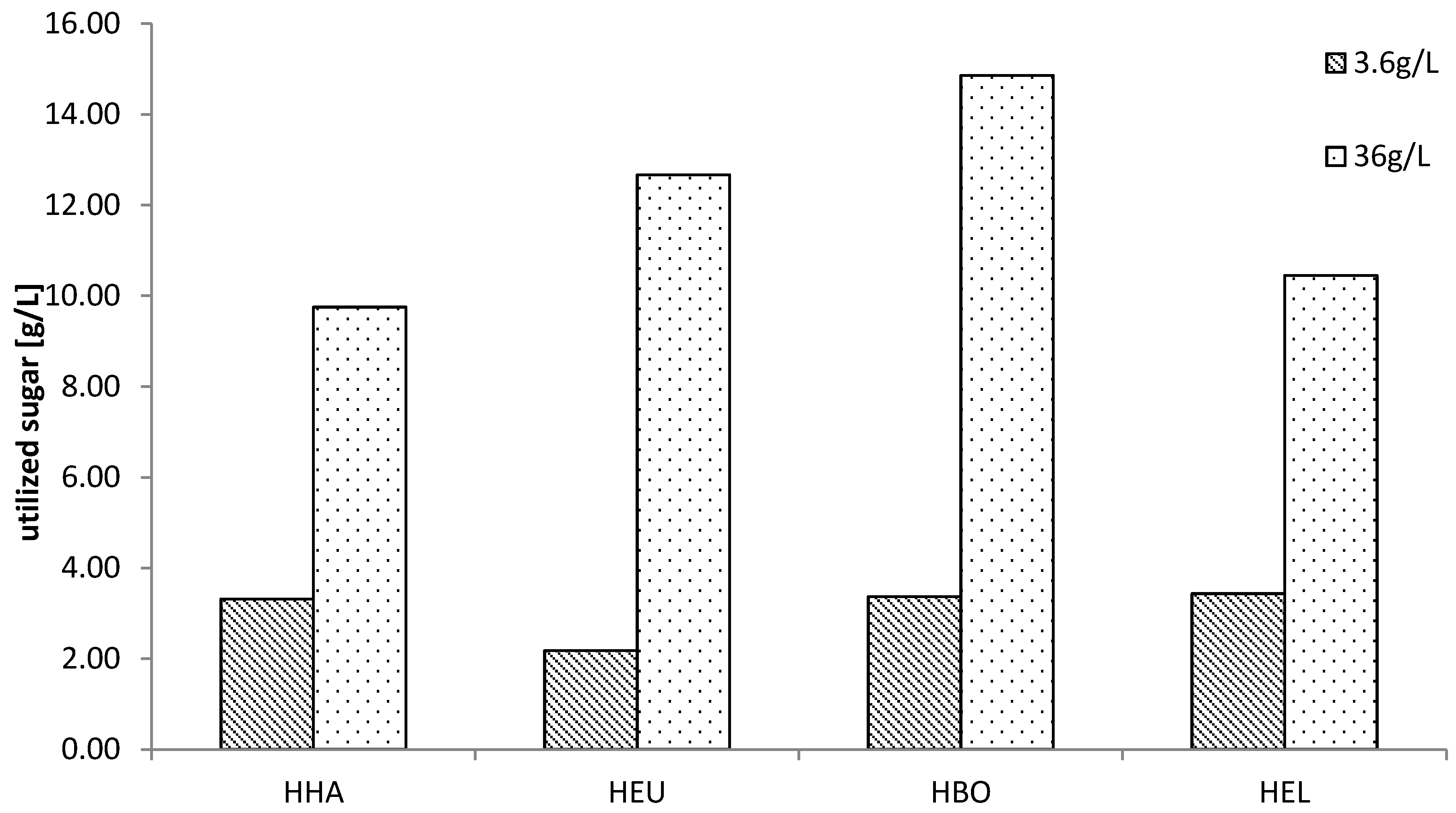
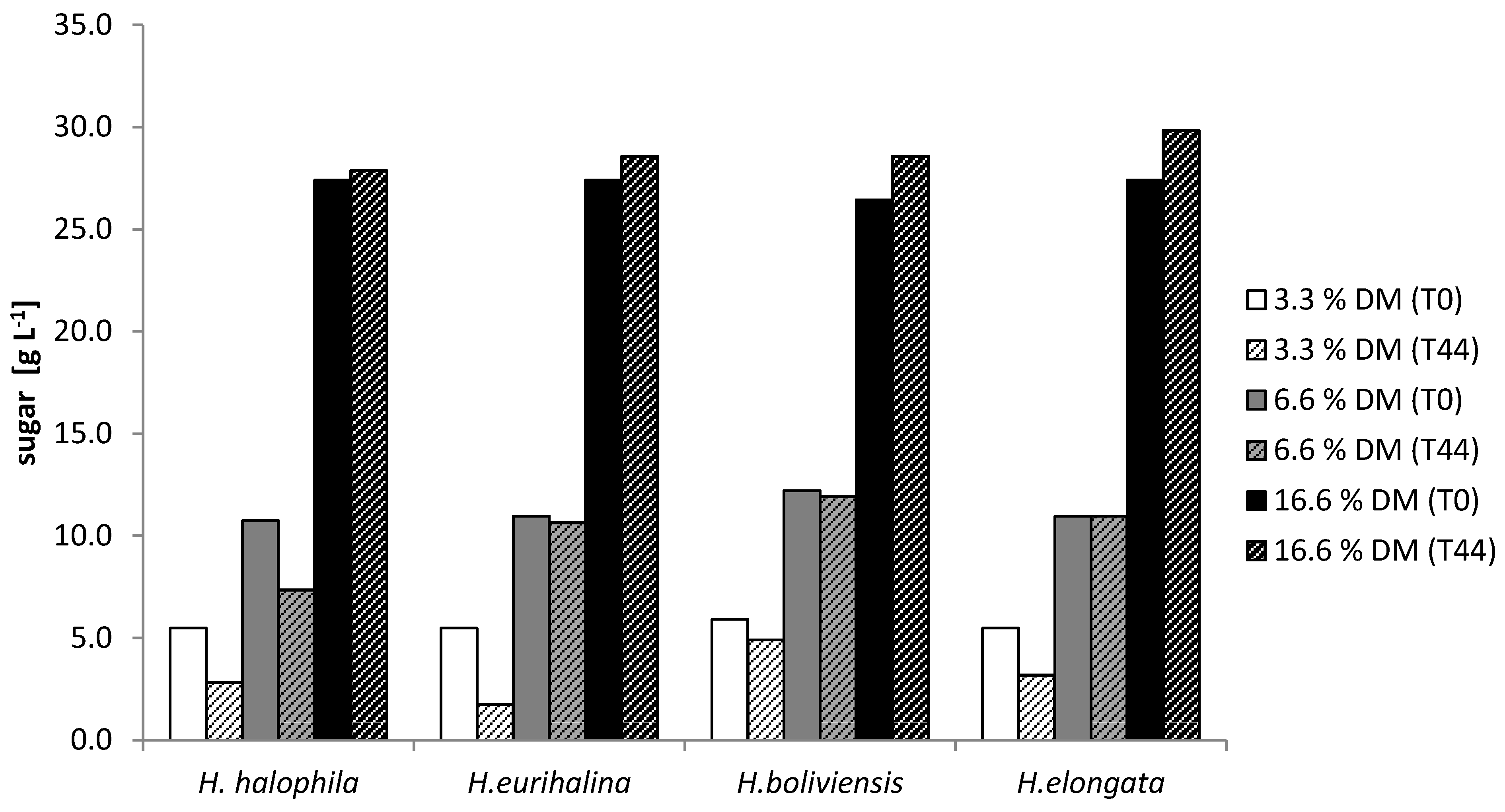
3.3. Growth Tolerance against Diluted SSL


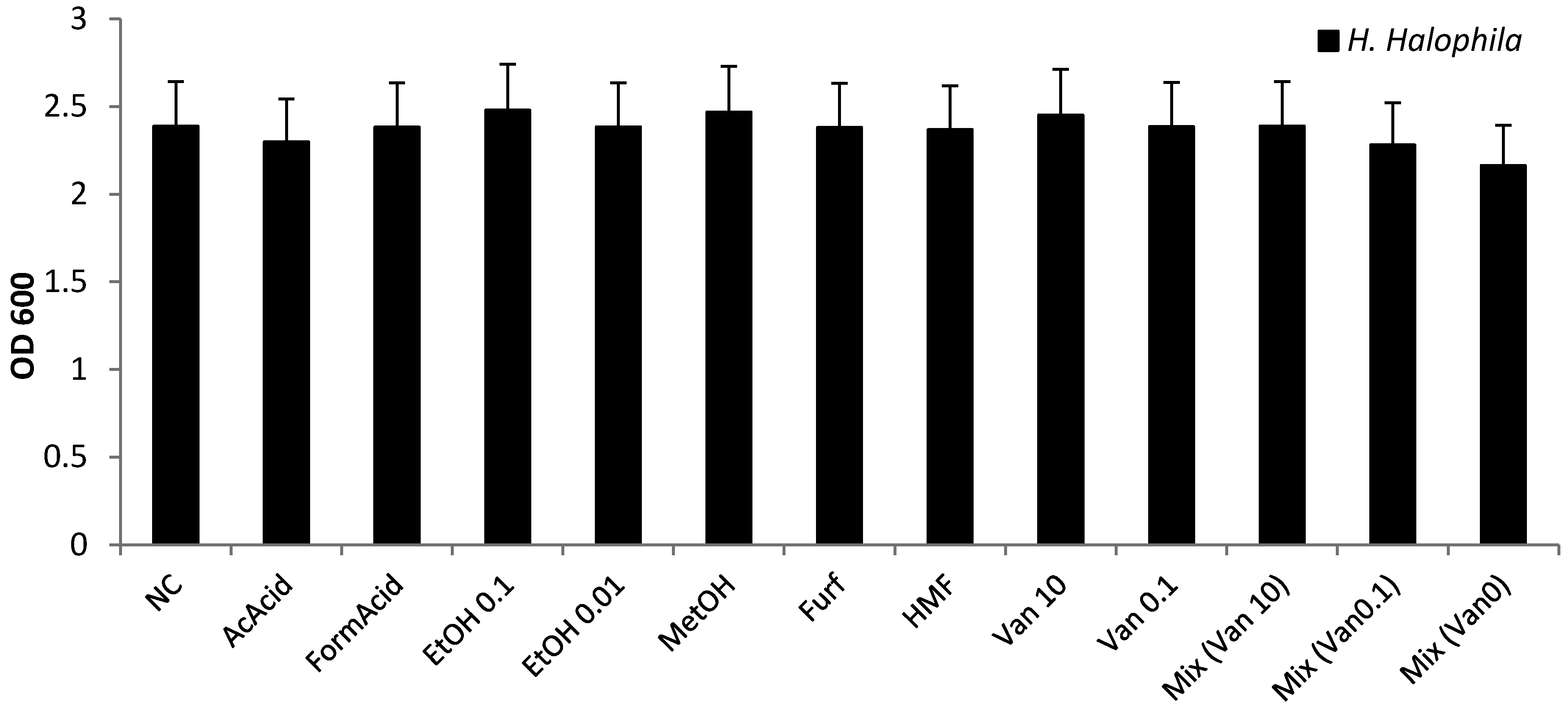
3.4. Effect of Known Inhibiting Substances in SSL on Growth

3.5. Production of PHB on SSL

4. Conclusions
- As the inhibitor experiments cannot explain the low tolerance of the halophilic microorganisms against SSL, the inhibitory effect of SSL must be caused by other, not yet investigated, factors like phenols other than vanillin or, less likely, the inorganic fraction.
- Detailed analysis of the phenolic and the inorganic fraction of pH adjusted SSL and the effect of single components on the investigated strains could be used for media adjustments that allow a much higher tolerance of the investigated halophilic strains to SSL.
- Can the Halophiles tolerate higher concentrations of SSL if they are slowly adapted to the inhibitors of this complex residual feedstock?
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sjöström, E. Wood Chemistry: Fundamentals and Applications; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Rydholm, S.A. Pulping Processes; Robert E. Krieger Publishing Company, Inc.: Malabar, FL, USA, 1985. [Google Scholar]
- Koch, G. Raw material for pulp. In Handbook of Pulp; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2008; pp. 21–68. [Google Scholar]
- Ezeji, T.; Qureshi, N.; Blaschek, H.P. Butanol production from agricultural residues: Impact of degradation products on clostridium beijerinckii growth and butanol fermentation. Biotechnol. Bioeng. 2007, 97, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Cho, D.; Lee, Y.; Um, Y.; Sang, B.-I.; Kim, Y. Detoxification of model phenolic compounds in lignocellulosic hydrolysates with peroxidase for butanol production from clostridium beijerinckii. Appl. Microbiol. Biotechnol. 2009, 83, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Ilmén, M.; Koivuranta, K.; Ruohonen, L.; Suominen, P.; Penttila, M. Efficient production of l-lactic acid from xylose by pichia stipitis. Appl. Microbiol. Biotechnol. 2007, 73, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Garde, A.; Jonsson, G.; Schmidt, A.S.; Ahring, B.K. Lactic acid production from wheat straw hemicellulose hydrolysate by lactobacillus pentosus and lactobacillus brevis. Bioresour. Technol. 2002, 81, 217–223. [Google Scholar] [CrossRef]
- Werpy, T.A.; Holladay, J.E.; White, J.F. Top Value Added Chemicals from Biomass: I. Results of Screening for Potential Candidates from Sugars and Synthesis Gas. U.S. Department of Energy: Oak Ridge, TN, USA, 2004. [Google Scholar]
- Akaraonye, E.; Keshavarz, T.; Roy, I. Production of polyhydroxyalkanoates: The future green materials of choice. J. Chem. Technol. Biotechnol. 2010, 85, 732–743. [Google Scholar] [CrossRef]
- Lu, Q.; Han, J.; Zhou, L.; Zhou, J.; Xiang, H. Genetic and biochemical characterization of the poly(3-hydroxybutyrate-co-3-hydroxyvalerate) synthase in haloferax mediterranei. J. Bacteriol. 2008, 190, 4173–4180. [Google Scholar] [CrossRef] [PubMed]
- Quillaguaman, J.; Delgado, O.; Mattiasson, B.; Hatti-Kaul, R. Poly(beta-hydroxybutyrate) production by a moderate halophile, Halomonas boliviensis LC1. Enzym. Microb. Technol. 2006, 38, 148–154. [Google Scholar] [CrossRef]
- Oren, A. Uptake and turnover of acetate in hypersaline environments. FEMS Microbiol. Ecol. 1995, 18, 75–84. [Google Scholar] [CrossRef]
- Bräsen, C.; Schönheit, P. Mechanisms of acetate formation and acetate activation in halophilic archaea. Arch. Microbiol. 2001, 175, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Gurevich, P. Diversity of lactate metabolism in halophilic archaea. Can. J. Microbiol. 1995, 41, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, I.J.; Márquez, M.C.; Xue, Y.; Ma, Y.; Cowan, D.A.; Jones, B.E.; Grant, W.D.; Ventosa, A. Bacillus chagannorensis sp. Nov., a moderate halophile from a soda lake in Inner Mongolia, China. Int. J. Syst. Evol. Microbiol. 2007, 57, 2084–2088. [Google Scholar]
- Tsu, I.H.; Huang, C.Y.; Garcia, J.L.; Patel, B.K.C.; Cayol, J.-L.; Baresi, L.; Mah, R.A. Isolation and characterization of Desulfovibrio senezii sp. Nov., a halotolerant sulfate reducer from a solar saltern and phylogenetic confirmation of desulfovibrio fructosovorans as a new species. Arch. Microbiol. 1998, 170, 313–317. [Google Scholar]
- Antón, J.; Meseguer, I.; Rodrífguez-Valera, F. Production of an extracellular polysaccharide by haloferax mediterranei. Appl. Environ. Microbiol. 1988, 54, 2381–2386. [Google Scholar] [PubMed]
- Cuadros-Orellana, S.; Pohlschröder, M.; Durrant, L.R. Isolation and characterization of halophilic archaea able to grow in aromatic compounds. Int. Biodeterior. Biodegrad. 2006, 57, 151–154. [Google Scholar] [CrossRef]
- Martinez, A.; Rodriguez, M.E.; Wells, M.L.; York, S.W.; Preston, J.F.; Ingram, L.O. Detoxification of dilute acid hydrolysates of lignocellulose with lime. Biotechnol. Progress 2001, 17, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Alriksson, B.; Cavka, A.; Jönsson, L.J. Improving the fermentability of enzymatic hydrolysates of lignocellulose through chemical in-situ detoxification with reducing agents. Bioresour. Technol. 2011, 102, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar]
- Alriksson, B.; Sjöde, A.; Nilvebrant, N.-O.; Jönsson, L. Optimal conditions for alkaline detoxification of dilute-acid lignocellulose hydrolysates. In Twenty-Seventh Symposium on Biotechnology for Fuels and Chemicals; McMillan, J., Adney, W., Mielenz, J., Klasson, K.T., Eds.; Humana Press: Totowa, NY, USA, 2006; pp. 599–611. [Google Scholar]
- Larsson, S.; Quintana-Sainz, A.; Reimann, A.; Nilvebrant, N.O.; Jonsson, L.J. Influence of lignocellulose-derived aromatic compounds on oxygen-limited growth and ethanolic fermentation by Saccharomyces cerevisiae. Appl. Biochem. Biotechnol. 2000, 84–86, 617–632. [Google Scholar]
- Martín, C.; Marcet, M.; Almazán, O.; Jönsson, L.J. Adaptation of a recombinant xylose-utilizing saccharomyces cerevisiae strain to a sugarcane bagasse hydrolysate with high content of fermentation inhibitors. Bioresour. Technol. 2007, 98, 1767–1773. [Google Scholar] [CrossRef] [PubMed]
- Olsson, L.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates for ethanol production. Enzym. Microb. Technol. 1996, 18, 312–331. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. Potential of halotolerant and halophilic microorganisms for biotechnology. Extremophiles 2001, 5, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A. Compatible solutes of halophilic eubacteria: Molecular principles, water-solute interaction, stress protection. Experientia 1993, 49, 487–496. [Google Scholar] [CrossRef]
- Fang, C.J.; Ku, K.L.; Lee, M.H.; Su, N.W. Influence of nutritive factors on C50 carotenoids production by haloferax mediterranei ATCC 33500 with two-stage cultivation. Bioresour. Technol. 2010, 101, 6487–6493. [Google Scholar] [CrossRef] [PubMed]
- Lorantfy, B.; Renkecz, T.; Koch, C.; Horvai, G.; Lendl, B.; Herwig, C. Identification of lipophilic bioproduct portfolio from bioreactor samples of extreme halophilic archaea with HPLC-MS/MS. Anal. Bioanal. Chem. 2014, 406, 2421–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margesin, R.; Schinner, F. Biodegradation and bioremediation of hydrocarbons in extreme environments. Appl. Microbiol. Biotechnol. 2001, 56, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Hermann-Krauss, C.; Koller, M.; Muhr, A.; Fasl, H.; Stelzer, F.; Braunegg, G. Archaeal production of polyhydroxyalkanoate (PHA) co- and terpolyesters from biodiesel industry-derived by-products. Archaea 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Legat, A.; Gruber, C.; Zangger, K.; Wanner, G.; Stan-Lotter, H. Identification of polyhydroxyalkanoates in halococcus and other haloarchaeal species. Appl. Microbiol. Biotechnol. 2010, 87, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, P.; Coker, J.A.; Huse, V.; DasSarma, S.; Flickinger, M.C. Halophiles, industrial applications. In Encyclopedia of Industrial Biotechnology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009. [Google Scholar]
- Lorantfy, B.; Seyer, B.; Herwig, C. Stoichiometric and kinetic analysis of extreme halophilic archaea on various substrates in a corrosion resistant bioreactor. New Biotechnol. 2014, 31, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Van, T.D. Production of Poly(3-Hydroxybutyrate) and Ectoines Using a Halophilic Bacterium. Ph.D. Thesis, Lund University, Lund, Sweden, 2009. [Google Scholar]
- Oshiki, M.; Satoh, H.; Mino, T. Rapid quantification of polyhydroxyalkanoates (PHA) concentration in activated sludge with the fluorescent dye nile blue A. Water Sci. Technol. 2011, 64, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Van-Thuoc, D.; Quillaguaman, J.; Mamo, G.; Mattiasson, B. Utilization of agricultural residues for poly(3-hydroxybutyrate) production by Halomonas boliviensis LC1. J. Appl. Microbiol. 2008, 104, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Quillaguamán, J.; Doan-Van, T.; Guzmán, H.; Guzmán, D.; Martín, J.; Everest, A.; Hatti-Kaul, R. Poly(3-hydroxybutyrate) production by Halomonas boliviensis in fed-batch culture. Appl. Microbiol. Biotechnol. 2008, 78, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Tekin, E.; Ates, M.; Kahraman, Ö. Poly-3-hydroxybutyrate-producing extreme halophilic archaeon: Haloferax sp. Ma10 isolated from çamaltı saltern, İzmir. Turk. J. Biol. 2012, 36, 303–312. [Google Scholar]
- Mata, J.A.; Martinez-Canovas, J.; Quesada, E.; Bejar, V. A detailed phenotypic characterisation of the type strains of halomonas species. Syst. Appl. Microbiol. 2002, 25, 360–375. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.; Reimann, A.; Nilvebrant, N.-O.; Jönsson, L. Comparison of different methods for the detoxification of lignocellulose hydrolyzates of spruce. Appl. biochem. Biotechnol. 1999, 77, 91–103. [Google Scholar] [CrossRef]
- Ackermann, J.U.; Babel, W. Growth-associated synthesis of poly(hydroxybutyric acid) in methylobacterium rhodesianum as an expression of an internal bottleneck. Appl. Microbiol. Biotechnol. 1997, 47, 144–149. [Google Scholar] [CrossRef]
- Keweloh, H.; Weyrauch, G.; Rehm, H.-J. Phenol-induced membrane changes in free and immobilized Escherichia coli. Appl. Microbiol. Biotechnol. 1990, 33, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Zaldivar, J.; Nielsen, J.; Olsson, L. Fuel ethanol production from lignocellulose: A challenge for metabolic engineering and process integration. Appl. Microbiol. Biotechnol. 2001, 56, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Ranatunga, T.D.; Jervis, J.; Helm, R.F.; McMillan, J.D.; Wooley, R.J. The effect of overliming on the toxicity of dilute acid pretreated lignocellulosics: The role of inorganics, uronic acids and ether-soluble organics. Enzym. Microb. Technol. 2000, 27, 240–247. [Google Scholar] [CrossRef]
- Maiorella, B.L.; Blanch, H.W.; Wilke, C.R. Feed component inhibition in ethanolic fermentation by Saccharomyces cerevisiae. Biotechnol. Bioeng. 1984, 26, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Vriesekoop, F.; Rasmusson, M.; Pamment, N.B. Respective effects of sodium and chloride ions on filament formation and growth and ethanol production in zymomonas mobilis fermentations. Lett. Appl. Microbiol. 2002, 35, 27–31. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weissgram, M.; Gstöttner, J.; Lorantfy, B.; Tenhaken, R.; Herwig, C.; Weber, H.K. Generation of PHB from Spent Sulfite Liquor Using Halophilic Microorganisms. Microorganisms 2015, 3, 268-289. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3020268
Weissgram M, Gstöttner J, Lorantfy B, Tenhaken R, Herwig C, Weber HK. Generation of PHB from Spent Sulfite Liquor Using Halophilic Microorganisms. Microorganisms. 2015; 3(2):268-289. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3020268
Chicago/Turabian StyleWeissgram, Michaela, Janina Gstöttner, Bettina Lorantfy, Raimund Tenhaken, Christoph Herwig, and Hedda K. Weber. 2015. "Generation of PHB from Spent Sulfite Liquor Using Halophilic Microorganisms" Microorganisms 3, no. 2: 268-289. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3020268