
Movement of Salmonella serovar Typhimurium and E. coli O157:H7 to Ripe Tomato Fruit Following Various Routes of Contamination


Abstract
:
1. Introduction
2. Experimental Section
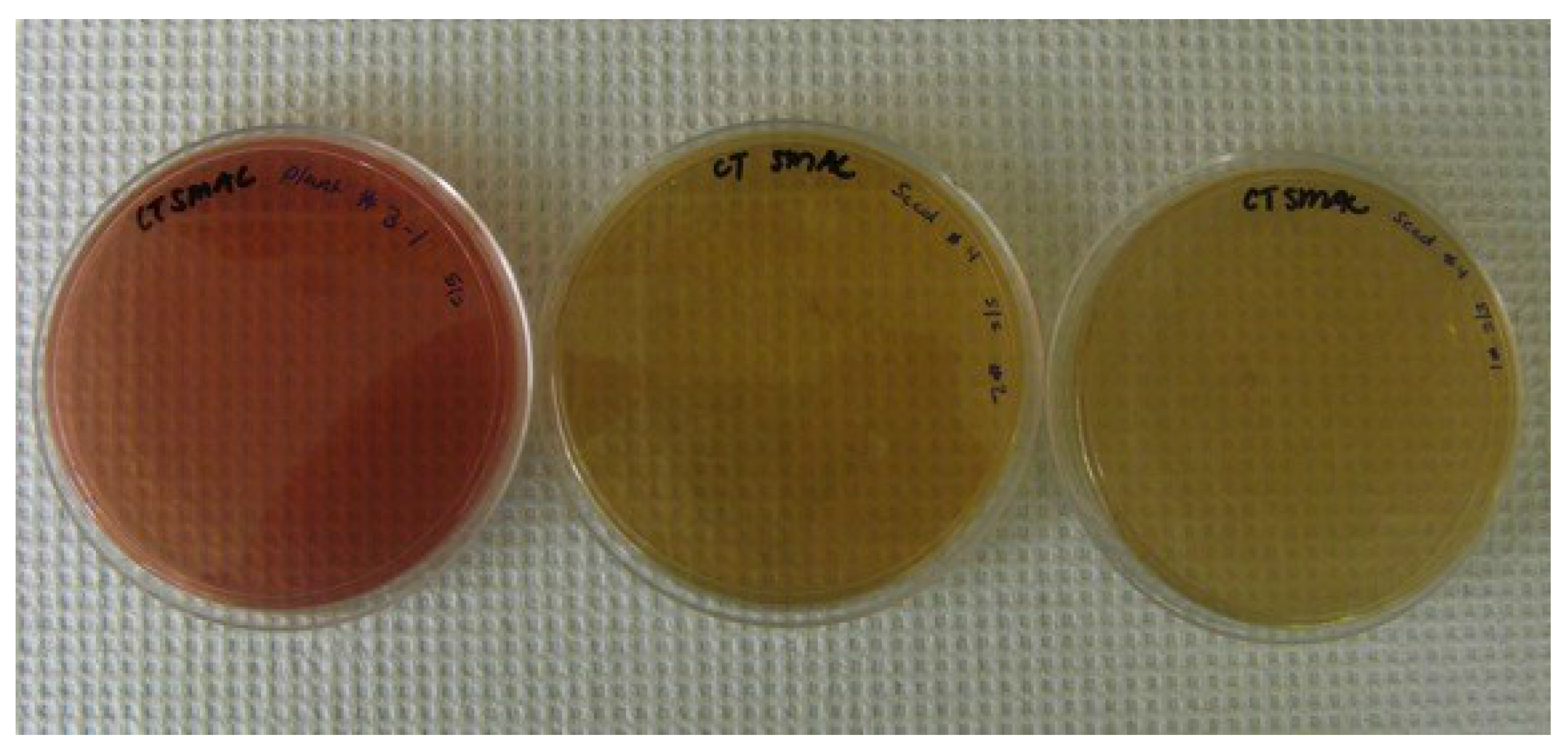
2.1. Bacteria
2.2. Tomato Varieties
2.3. Inoculation of Plants and Growing Conditions
2.3.1. Treatment for Seed-Contaminated Plants
2.3.2. Treatment for Soil-Contaminated Plants
2.3.3. Treatment for Leaf-Contaminated Plants
2.3.4. Treatment for Control Plants
2.4. Sampling of Ripe Tomato Fruit
2.5. PCR for the Detection of GFP
2.6. Confirmation of E. coli O157:H7-GFP
2.7. Persistence of E. coli O157:H7-GFP and Salmonella Typhimurium-GFP
2.8. Statistical Analysis
3. Results and Discussion
3.1. Contaminating Tomato Plants with E. coli O157:H7-GFP and Salmonella Typhimurium-GFP Affected Plant Health and Tomato Production



3.2. Effects of Routes of Contamination by E. coli O157:H7-GFP



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Contamination Type | Total Tomatoes Produced (and Sampled) | % GFP Positive |
|---|---|---|---|
| Micro-Tom | Seed | 72 | 54.2% |
| Soil | 57 | 0% | |
| Leaf | 49 | 38.8% | |
| Tommy Toe | Seed | 0 | - |
| Soil | 3 | 66.7% | |
| Leaf | 0 | - | |
| Red Currant | Seed | 5 | 60.0% |
| Soil | 0 | - | |
| Leaf | 0 | - | |
| Sweet Quartz | Seed | 3 | 100% |
| Soil | 0 | - | |
| Leaf | 0 | - | |
| Roma | Seed | 25 | 40% |
| Soil | 12 | 50% | |
| Leaf | 14 | 21.4% | |
| Elfin | Seed | 0 | - |
| Soil | 0 | - | |
| Leaf | 2 | 0% | |
| Total sampled | - | 242 | 35.1% |

3.3. Effects of Routes of Contamination by Salmonella Typhimurium-GFP
3.4. Persistence of E. coli O157:H7-GFP and Salmonella Typhimurium-GFP

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- U.S. Food and Drug Administration (FDA). Guidance for Industry: Guide to Minimize Microbial Food Safety Hazards of Tomatoes; Draft Guidance 2009; FDA: Silver Spring, MA, USA, 2013.
- Reller, M.E.; Nelson, J.M.; Molbak, K.; Ackman, D.M.; Schoonmaker-Bopp, D.J.; Root, T.P.; Mintz, E.D. A large, multiple-restaurant outbreak of infection with Shigella flexneri serotype 2a traced to tomatoes. Clin. Infect. Dis. 2006, 42, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Outbreaks of Salmonella infections associated with eating roma tomatoes—United States and Canada, 2004. MMWR Morb. Mortal. Wkly. Rep. 2005, 54, 325–328. [Google Scholar]
- Barak, J.D.; Liang, A.S. Role of soil, crop debris, and a plant pathogen in Salmonella enterica contamination of tomato plants. PLoS ONE 2008, 3, e1657. [Google Scholar] [CrossRef] [PubMed]
- Orozco, L.; Rico-Romero, L.; Escartin, E.F. Microbiological profile of greenhouses in a farm producing hydroponic tomatoes. J. Food Prot. 2008, 71, 60–65. [Google Scholar] [PubMed]
- Guo, X.; Chen, J.; Brackett, R.E.; Beuchat, L.R. Survival of Salmonellae on and in tomato plants from the time of inoculation at flowering and early stages of fruit development through fruit ripening. Appl. Environ. Microbiol. 2001, 67, 4760–4764. [Google Scholar] [CrossRef] [PubMed]
- Rathinasabapathi, B. Survival of Salmonella Montevideo on tomato leaves and mature green tomatoes. J. Food Prot. 2004, 67, 2277–2279. [Google Scholar] [PubMed]
- Iturriaga, M.H.; Tamplin, M.L.; Escartin, E.F. Colonization of tomatoes by Salmonella Montevideo is affected by relative humidity and storage temperature. J. Food Prot. 2007, 70, 30–34. [Google Scholar] [PubMed]
- Shi, X.; Namvar, A.; Kostrzynska, M.; Hora, R.; Warriner, K. Persistence and growth of different Salmonella serovars on pre- and postharvest tomatoes. J. Food Prot. 2007, 70, 2725–2731. [Google Scholar] [PubMed]
- Koodie, L.; Dhople, A.M. Acid tolerance of Escherichia coli O157:H7 and its survival in apple juice. Microbios 2001, 104, 167–175. [Google Scholar] [PubMed]
- Weagant, S.D.; Bryant, J.L.; Bark, D.H. Survival of Escherichia coli O157: H7 in mayonnaise and mayonnaise-based sauces at room and refrigerated temperatures. J. Food Prot. 1994, 57, 629–631. [Google Scholar]
- Eribo, B.; Ashenafi, M. Behavior of Escherichia coli O157:H7 in tomato and processed tomato products. Food Res. Int. 2003, 36, 823–830. [Google Scholar] [CrossRef]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157:H7 outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Ongoing multistate outbreak of Escherichia coli serotype O157:H7 infections associated with consumption of fresh spinach—United States, September 2006. In MMWR Morb. Mortal. Wkly. Rep.; 2006; 55, pp. 1045–1046. [Google Scholar]
- Slayton, R.B.; Turabelidze, G.; Bennett, S.D.; Schwensohn, C.A.; Yaffee, A.Q.; Khan, F.; Butler, C.; Trees, E.; Ayers, T.L.; Davis, M.L.; et al. Outbreak of shiga toxin-producing Escherichia coli (STEC) O157:H7 associated with romaine lettuce consumption, 2011. PLoS ONE 2013, 8, e55300. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Cevallos-Cevallos, J.M.; Vallad, G.E.; van Bruggen, A.H. Organically managed soils reduce internal colonization of tomato plants by Salmonella enterica serovar Typhimurium. Phytopathology 2013, 103, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Cevallos-Cevallos, J.M.; Gu, G.; Danyluk, M.D.; Dufault, N.S.; van Bruggen, A.H. Salmonella can reach tomato fruits on plants exposed to aerosols formed by rain. Int. J. Food Microbiol. 2012, 158, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Hu, J.; Cevallos-Cevallos, J.M.; Richardson, S.M.; Bartz, J.A.; van Bruggen, A.H. Internal colonization of Salmonella enterica serovar Typhimurium in tomato plants. PLoS ONE 2011, 6, e27340. [Google Scholar] [CrossRef] [PubMed]
- Fratamico, P.M.; Deng, M.Y.; Strobaugh, T.P.; Palumbo, S.A. Construction and characterization of Escherichia coli O157:H7 strains expressing firefly luciferase and green fluorescent protein and their use in survival studies. J. Food Prot. 1997, 60, 1167–1173. [Google Scholar]
- Kudva, I.T.; Blanch, K.; Hovde, C.J. Analysis of Escherichia coli O157:H7 survival in ovine or bovine manure and manure slurry. Appl. Environ. Microbiol. 1998, 64, 3166–3174. [Google Scholar] [PubMed]
- Malorny, B.; Hoorfar, J.; Bunge, C.; Helmuth, R. Multicenter validation of the analytical accuracy of Salmonella PCR: Towards an international standard. Appl. Environ. Microbiol. 2003, 69, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Rahn, K.; de Grandis, S.A.; Clarke, R.C.; McEwen, S.A.; Galan, J.E.; Ginocchio, C.; Curtiss, R., III; Gyles, C.L. Amplification of an invA gene sequence of Salmonella Typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell. Probes 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Deering, A.J.; Pruitt, R.E. Purdue University: West Lafayette, IN, USA, Unpublished work. 2015.
- Errampalli, D.; Leung, K.; Cassidy, M.B.; Kostrzynska, M.; Blears, M.; Lee, H.; Trevors, J.T. Applications of the green fluorescent protein as a molecular marker in environmental microorganisms. J. Microbiol. Methods 1999, 35, 187–199. [Google Scholar] [CrossRef]
- Klerks, M.M.; van Gent-Pelzer, M.; Franz, E.; Zijlstra, C.; van Bruggen, A.H. Physiological and molecular responses of Lactuca sativa to colonization by Salmonella enterica serovar Dublin. Appl. Environ. Microbiol. 2007, 73, 4905–4914. [Google Scholar] [CrossRef] [PubMed]
- Klement, Z.; Goodman, R.N. The hypersensitive reaction to infection by bacterial plant pathogens. Annu. Rev. Phytopathol. 1967, 5, 17–44. [Google Scholar] [CrossRef]
- Warriner, K.; Ibrahim, F.; Dickinson, M.; Wright, C.; Waites, W.M. Internalization of human pathogens within growing salad vegetables. Biotechnol. Genet. Eng. Rev. 2003, 20, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Allard, S.; Reynolds, S.; Millner, P.; Arce, G.; Blodgett, R.J.; Brown, E.W. Colonization and internalization of Salmonella enterica in tomato plants. Appl. Environ. Microbiol. 2013, 79, 2494–2502. [Google Scholar] [CrossRef] [PubMed]
- Barak, J.D.; Kramer, L.C.; Hao, L.Y. Colonization of tomato plants by Salmonella enterica is cultivar dependent, and type 1 trichomes are preferred colonization sites. Appl. Environ. Microbiol. 2011, 77, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Cevallos-Cevallos, J.M.; Gu, G.; Danyluk, M.D.; van Bruggen, A.H. Adhesion and splash dispersal of Salmonella enterica Typhimurium on tomato leaflets: Effects of rdar morphotype and trichome density. Int. J. Food Microbiol. 2012, 160, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Cevallos-Cevallos, J.M.; van Bruggen, A.H. Ingress of Salmonella enterica Typhimurium into tomato leaves through hydathodes. PLoS ONE 2013, 8, e53470. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Multistate outbreaks of Salmonella infections associated with raw tomatoes eaten in restaurants—United States, 2005–2006. In MMWR Morb. Mortal. Wkly. Rep.; 2007; 56, pp. 909–911. [Google Scholar]
- Noel, J.T.; Arrach, N.; Alagely, A.; McClelland, M.; Teplitski, M. Specific responses of Salmonella enterica to tomato varieties and fruit ripeness identified by in vivo expression technology. PLoS ONE 2010, 5, e12406. [Google Scholar] [CrossRef] [PubMed]
- Hintz, L.D.; Boyer, R.R.; Ponder, M.A.; Williams, R.C.; Rideout, L.C. Recovery of Salmonella enterica Newport introduced through irrigation water from tomato (Lycopersicum esculentum) fruit, roots, stems, and leaves. HortSci. 2010, 45, 675–678. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deering, A.J.; Jack, D.R.; Pruitt, R.E.; Mauer, L.J. Movement of Salmonella serovar Typhimurium and E. coli O157:H7 to Ripe Tomato Fruit Following Various Routes of Contamination. Microorganisms 2015, 3, 809-825. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040809
Deering AJ, Jack DR, Pruitt RE, Mauer LJ. Movement of Salmonella serovar Typhimurium and E. coli O157:H7 to Ripe Tomato Fruit Following Various Routes of Contamination. Microorganisms. 2015; 3(4):809-825. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040809
Chicago/Turabian StyleDeering, Amanda J., Dan R. Jack, Robert E. Pruitt, and Lisa J. Mauer. 2015. "Movement of Salmonella serovar Typhimurium and E. coli O157:H7 to Ripe Tomato Fruit Following Various Routes of Contamination" Microorganisms 3, no. 4: 809-825. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040809