Prevalence of Resistance to β-Lactam Antibiotics and bla Genes Among Commensal Haemophilus parainfluenzae Isolates from Respiratory Microbiota in Poland

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Culture and Identification
2.3. Antimicrobial Susceptibility Testing
2.4. DNA Extraction
2.5. Amplification Experiments and Gene Detection
2.6. Statistical Analysis
3. Results
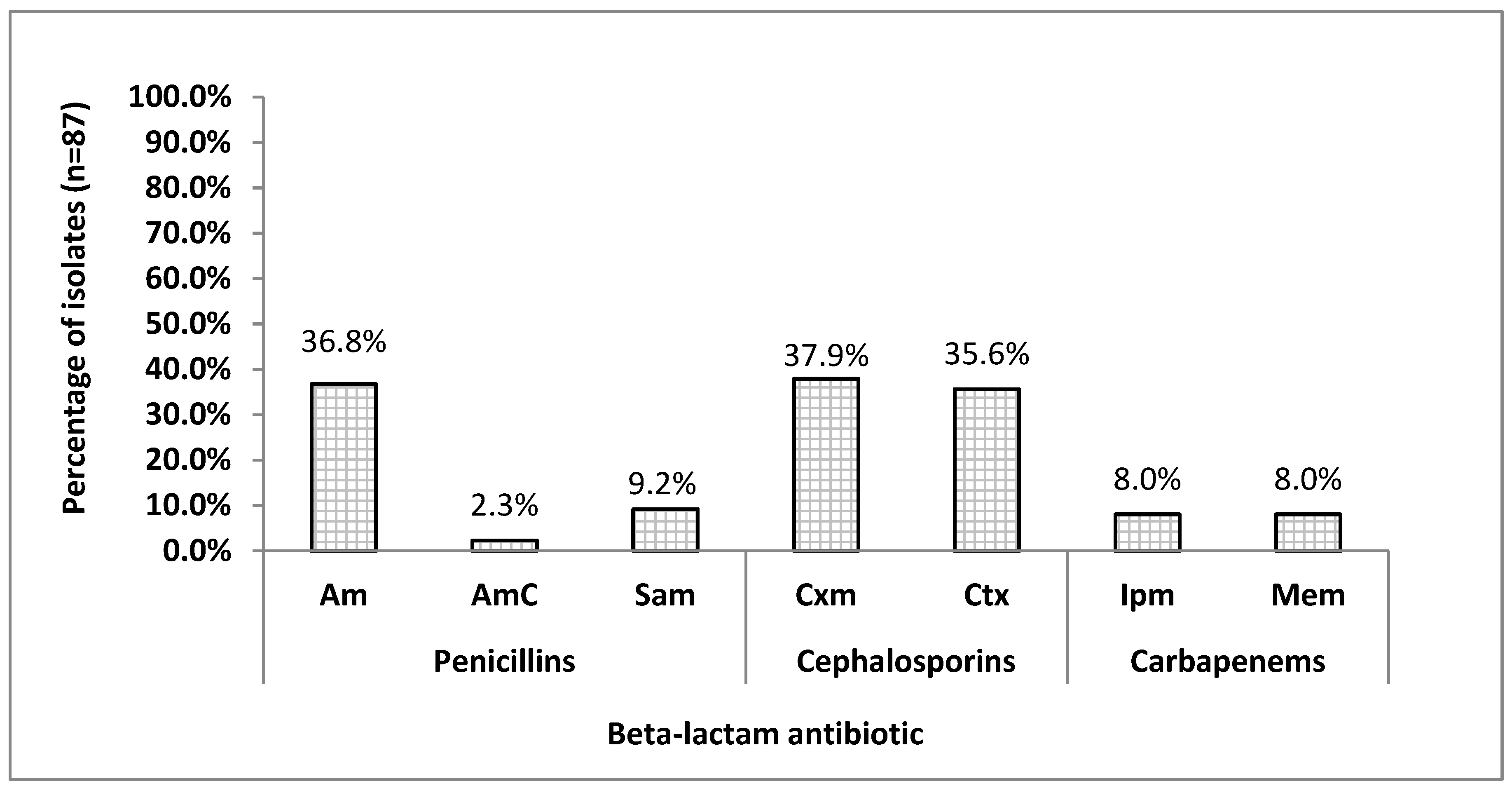
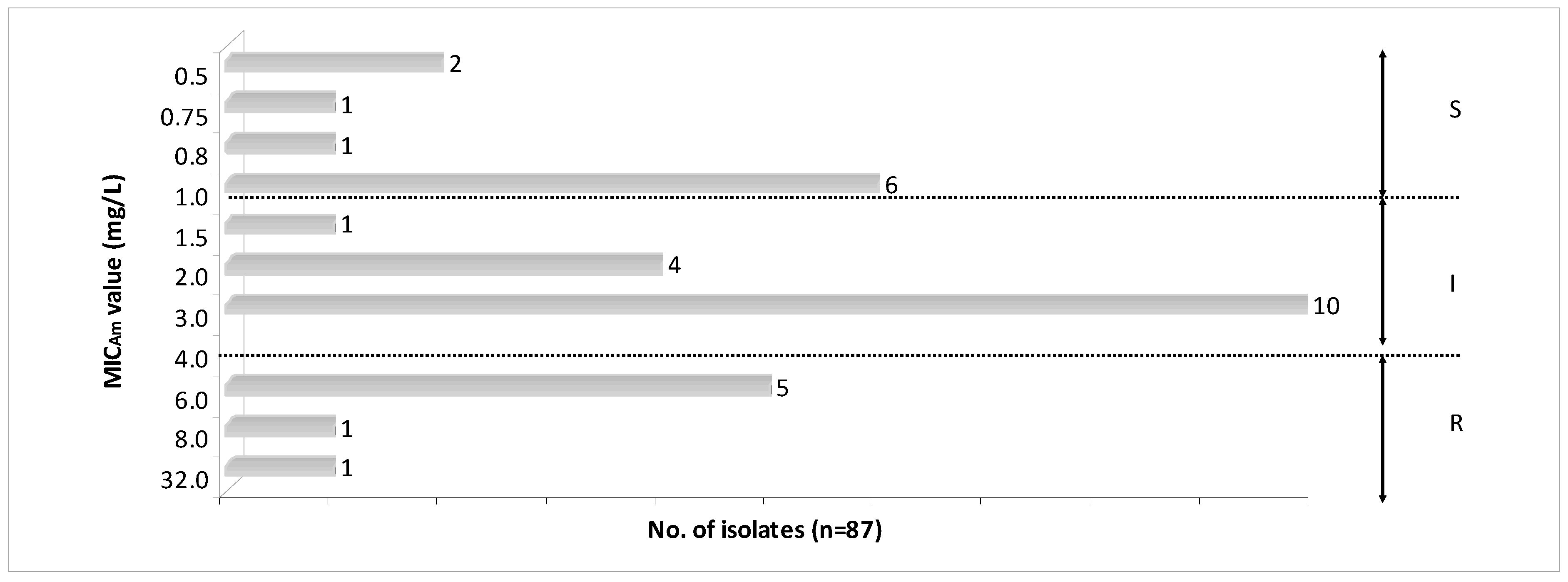
3.1. Antimicrobial Susceptibility Patterns Among H.parainfluenzae Isolates
3.2. The incidence of β-Lactamase-Positive Isolates According to Phenotypic Methods
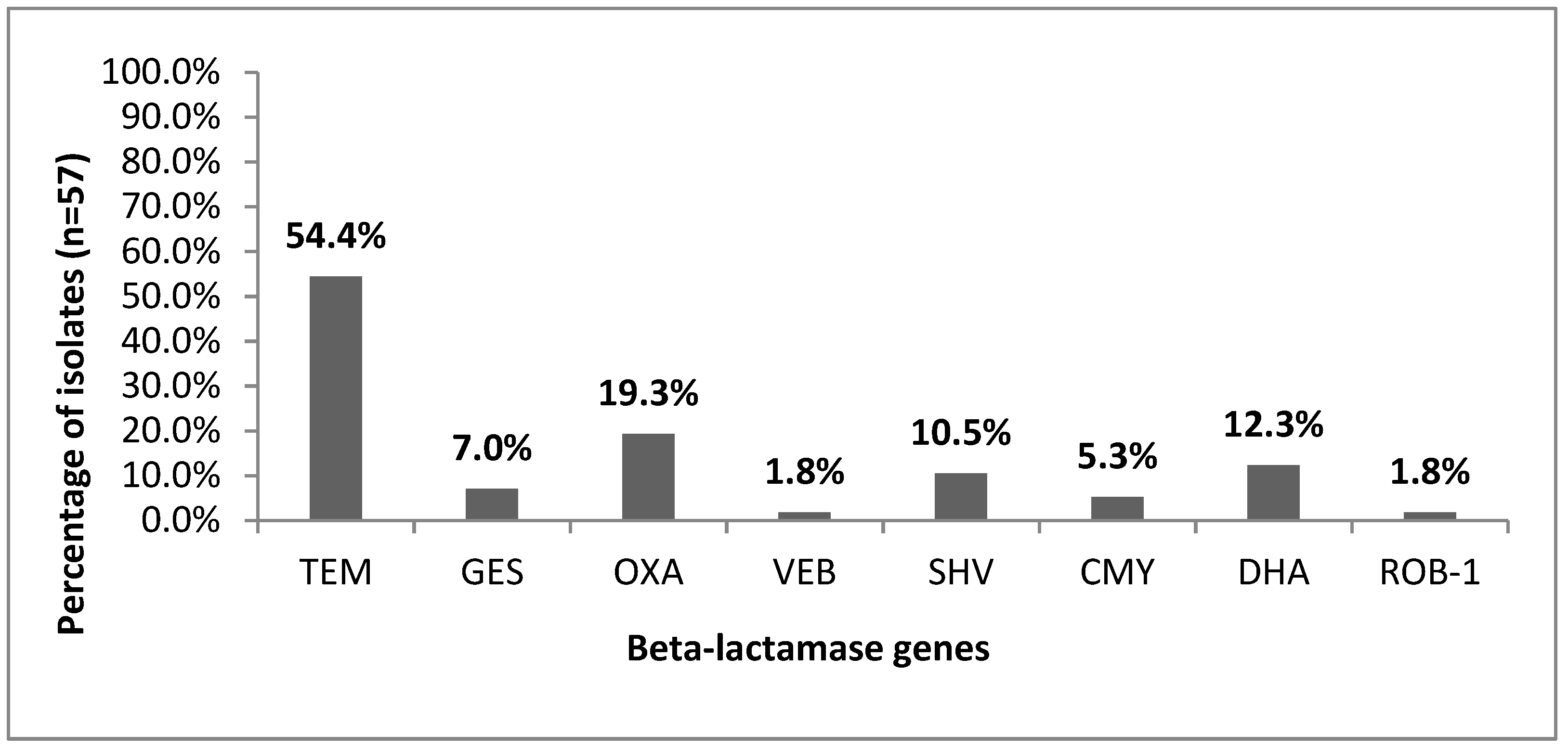
3.3. Prevalence of β-Lactamase Genes
3.4. Detection of ftsI Gene and Amino Acid Substitutions.
3.5. Relationship between Susceptibility to β-Lactam Antibiotics and Resistance Genes
4. Discussion
4.1. Resistance to Beta-Lactams among H. parainfluenzae
4.2. The Susceptibility to Beta-Lactam Antibiotics among H. parainfluenzae Isolates
4.3. Conventional Methods for Detection of Beta-Lactamase in Haemophili
4.4. PCR Amplification as Rapid Detection Method of Beta-Lactamase Production
4.5. Prevalence of Beta-Lactamase Type TEM-1 and ROB-1
4.6. Discrepancies in Phenotype and Genotype Classification for H. parainfluenzae
4.7. Polymorphism of the ftsI Resistance Gene
4.8. Beta-Lactamase-Negative ftsI-Positive Isolates
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Centre for Disease Prevention and Control Antimicrobial Consumption. Annual Epidemiological Report for 2017; ECDC: Stockholm, Sweden, 2018.
- Yeh, S. Epidemiology, Clinical Manifestations, and Treatment of Haemophilus influenzae. Available online: https://www.uptodate.com/contents/epidemiology-clinical-manifestations-and-treatment-of-haemophilus-influenzae (accessed on 28 May 2018).
- Mandell, L.A.; Wunderink, R.G.; Anzueto, A.; Bartlett, J.G.; Campbell, G.D.; Dean, N.C.; Dowell, S.F.; File, T.M., Jr.; Musher, D.M.; Niederman, M.S.; et al. Infectious Diseases Society of America/American Thoracic Society Consensus Guidelines on the Management of Community-Acquired Pneumonia in Adults. Clin. Infect. Dis. 2007, 44, S27–S72. [Google Scholar] [CrossRef] [PubMed]
- Hryniewicz, W.; Albrecht, P.; Radzikowski, A. Rekomendacje Postępowania w Pozaszpitalnych Zakażeniach Układu Oddechowego, 2nd ed.; The National Medicines Institute: Warszawa, Poland, 2016; ISBN 978-83-938000-5-6. [Google Scholar]
- Tristram, S.; Jacobs, M.R.; Appelbaum, P.C. Antimicrobial resistance in Haemophilus influenzae. Clin. Microbiol. Rev. 2007, 20, 368–389. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S. Mechanisms of antimicrobial resistance in Pasteurellaceae. In Pasteurellaceae. Biology and Molecular Aspects; Kuhnert, P., Christensen, H., Eds.; Caister Academic Press: Norfolk, UK, 2008; pp. 197–226. [Google Scholar]
- Tinguely, R.; Seiffert, S.N.; Furrer, H.; Perreten, V.; Droz, S.; Endimiani, A. Emergence of extensively drug-resistant Haemophilus parainfluenzae in Switzerland. Antimicrob. Agents Chemother. 2013, 57, 2867–2869. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Chiba, N.; Kobayashi, R.; Murayama, S.Y.; Iwata, S.; Sunakawa, K.; Ubukata, K. Rapidly increasing prevalence of beta-lactamase-nonproducing, ampicillin-resistant Haemophilus influenzae type b in patients with meningitis. Antimicrob. Agents Chemother. 2004, 48, 1509–1514. [Google Scholar] [CrossRef] [PubMed]
- Ubukata, K.; Shibasaki, Y.; Yamamoto, K.; Chiba, N.; Hasegawa, K.; Takeuchi, Y.; Sunakawa, K.; Inoue, M.; Konno, M. Association of amino acid substitutions in penicillin-binding protein 3 with beta-lactam resistance in beta-lactamase-negative ampicillin-resistant Haemophilus influenzae. Antimicrob. Agents Chemother. 2001, 45, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Yamamoto, K.; Chiba, N.; Kobayashi, R.; Nagai, K.; Jacobs, M.R.; Appelbaum, P.C.; Sunakawa, K.; Ubukata, K. Diversity of Ampicillin-Resistance Genes in Haemophilus influenzae in Japan and the United States. Microb. Drug Resist. 2003, 9, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Osaki, Y.; Sanbongi, Y.; Ishikawa, M.; Kataoka, H.; Suzuki, T.; Maeda, K.; Ida, T. Genetic approach to study the relationship between penicillin-binding protein 3 mutations and Haemophilus influenzae beta-lactam resistance by using site-directed mutagenesis and gene recombinants. Antimicrob. Agents Chemother. 2005, 49, 2834–2839. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-S.; Ki, C.-S.; Kim, S.; Oh, W.S.; Peck, K.R.; Song, J.-H.; Lee, K.; Lee, N.Y. Diversity of ampicillin resistance genes and antimicrobial susceptibility patterns in Haemophilus influenzae strains isolated in Korea. Antimicrob. Agents Chemother. 2007, 51, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Wienholtz, N.H.; Barut, A.; Norskov-Lauritsen, N. Substitutions in PBP3 confer resistance to both ampicillin and extended-spectrum cephalosporins in Haemophilus parainfluenzae as revealed by site-directed mutagenesis and gene recombinants. J. Antimicrob. Chemother. 2017, 72, 2544–2547. [Google Scholar] [CrossRef]
- Abotsi, R.E.; Govinden, U.; Moodley, K.; Essack, S. Fluoroquinolone, Macrolide, and Ketolide Resistance in Haemophilus parainfluenzae from South Africa. Microb. Drug Resist. 2017, 23, 667–673. [Google Scholar] [CrossRef]
- Kosikowska, U.; Biernasiuk, A.; Rybojad, P.; Łoś, R.; Malm, A. Haemophilus parainfluenzae as a marker of the upper respiratory tract microbiota changes under the influence of preoperative prophylaxis with or without postoperative treatment in patients with lung cancer. BMC Microbiol. 2016, 16, 62. [Google Scholar] [CrossRef] [PubMed]
- Sondergaard, A.; Norskov-Lauritsen, N. Contribution of PBP3 Substitutions and TEM-1, TEM-15, and ROB-1 Beta-Lactamases to Cefotaxime Resistance in Haemophilus influenzae and Haemophilus parainfluenzae. Microb. Drug Resist. 2016, 22, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Tristram, S.G.; Nichols, S. A multiplex PCR for beta-lactamase genes of Haemophilus influenzae and description of a new blaTEM promoter variant. J. Antimicrob. Chemother. 2006, 58, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Touati, A.; Achour, W.; Ben Hassen, A. Phenotypic and molecular characterization of beta-lactam resistance and capsular typing of colonizing Haemophilus influenzae strains isolated from neutropenic patients in Tunisia. Pathol. Biol. 2009, 57, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Yanagihara, K.; Morinaga, Y.; Izumikawa, K.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Kamihira, S.; Kohno, S. Multiplex real-time polymerase chain reaction for rapid detection of β-lactamase–negative, ampicillin-resistant Haemophilus influenzae. Diagn. Microbiol. Infect. Dis. 2009, 64, 64–69. [Google Scholar] [CrossRef]
- Harunur, R.; Mahbubur, R. Detection of β-lactamase in Haemophilus influenzae Isolates by Double Disk Synergy Test. J. Microb. Biochem. Technol. 2015, 7, 417–418. [Google Scholar]
- García-Cobos, S.; Arroyo, M.; Campos, J.; Pérez-Vázquez, M.; Aracil, B.; Cercenado, E.; Orden, B.; Lara, N.; Oteo, J. Novel mechanisms of resistance to β-lactam antibiotics in Haemophilus parainfluenzae: β-lactamase-negative ampicillin resistance and inhibitor-resistant TEM β-lactamases. J. Antimicrob. Chemother. 2013, 68, 1054–1059. [Google Scholar] [CrossRef]
- Farrell, D.J.; Morrissey, I.; Bakker, S.; Buckridge, S.; Felmingham, D. Global distribution of TEM-1 and ROB-1 β-lactamases in Haemophilus influenzae. J. Antimicrob. Chemother. 2005, 56, 773–776. [Google Scholar] [CrossRef]
- Tristram, S.G.; Pitout, M.J.; Forward, K.; Campbell, S.; Nichols, S.; Davidson, R.J. Characterization of extended-spectrum β-lactamase-producing isolates of Haemophilus parainfluenzae. J. Antimicrob. Chemother. 2008, 61, 509–514. [Google Scholar] [CrossRef]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.M.D.; Kamal, M.A. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi. J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef]
- Sondergaard, A.; San Millan, A.; Santos-Lopez, A.; Nielsen, S.M.; Gonzalez-Zorn, B.; Norskov-Lauritsen, N. Molecular organization of small plasmids bearing blaTEM-1 and conferring resistance to beta-lactams in Haemophilus influenzae. Antimicrob. Agents Chemother. 2012, 56, 4958–4960. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, A.; Stemplinger, I.; Jungwirth, R.; Giamarellou, H. Characterization of the plasmidic beta-lactamase CMY-2, which is responsible for cephamycin resistance. Antimicrob. Agents Chemother. 1996, 40, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraniak, A.; Fiett, J.; Hryniewicz, W.; Nordmann, P.; Gniadkowski, M. Ceftazidime-hydrolysing CTX-M-15 extended-spectrum β-lactamase (ESBL) in Poland. J. Antimicrob. Chemother. 2002, 50, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Barnaud, G.; Arlet, G.; Verdet, C.; Gaillot, O.; Lagrange, P.H.; Philippon, A. Salmonella enteritidis: AmpC plasmid-mediated inducible beta-lactamase (DHA-1) with an ampR gene from Morganella morganii. Antimicrob. Agents Chemother. 1998, 42, 2352–2358. [Google Scholar] [CrossRef] [PubMed]
- Ambler, R.P.; Coulson, A.F.; Frère, J.M.; Ghuysen, J.M.; Joris, B.; Forsman, M.; Levesque, R.C.; Tiraby, G.; Waley, S.G. A standard numbering scheme for the class A beta-lactamases. Biochem. J. 1991, 276, 269–270. [Google Scholar] [CrossRef] [PubMed]
- Kosikowska, U.; Stępień-Pyśniak, D.; Ożga, D.; Wernicki, A.; Malm, A. Identification of Bacillus spp. colonizing the nasal mucosa of healthy adults living in the suburban area using the matrix-assisted laser desorption-ionization time-of-flight mass spectrometry (MALDI-TOF MS) system. Curr. Issues Pharm. Med. Sci. 2014, 27, 137–141. [Google Scholar] [CrossRef]
- Kosikowska, U.; Stępień-Pyśniak, D.; Pietras-Ożga, D.; Andrzejczuk, S.; Juda, M.; Malm, A. Zastosowanie spektrometrii masowej MALDI-TOF MS w identyfikacji bakterii izolowanych z materiałów klinicznych od ludzi i zwierząt. Application of MALDI-TOF MS for identification of clinical isolates of bacteria from humans and animals. Diagn. Lab. J. Lab. Diagn. 2015, 1, 23–30. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 9.0. 2019. Available online: http://www.eucast.org/ (accessed on 15 April 2018).
- Janowicz, D.M.; Li, W.; Bauer, M.E. Host-pathogen Interplay of Haemophilus ducreyi. NIH-PA Author Manuscr. 2011, 23, 64–69. [Google Scholar] [CrossRef]
- De Medici, D.; Croci, L.; Delibato, E.; Di Pasquale, S.; Filetici, E.; Toti, L. Evaluation of DNA Extraction Methods for Use in Combination with SYBR Green I Real-Time PCR To Detect Salmonella enterica Serotype Enteritidis in Poultry. Appl. Environ. Microbiol. 2003, 69, 3456–3461. [Google Scholar] [CrossRef]
- Marques, A.S.; Moraes, E.P.; Júnior, M.A.A.; Moura, A.D.; Neto, V.F.A.; Neto, R.M.; Lima, K.M.G. Rapid discrimination of Klebsiella pneumoniae carbapenemase 2—Producing and non-producing Klebsiella pneumoniae strains using near-infrared spectroscopy (NIRS) and multivariate analysis. Talanta 2015, 134, 126–131. [Google Scholar] [CrossRef]
- Salgueiro, V.A.R. Characterização dos mecanismos de resistência aos antibióticos em estirpes de origem humana, ambientes associados aos cuidados de saúde e veterinária. Universidade de Lisboa, 2015. Available online: http://hdl.handle.net/10451/22527 (accessed on 30 May 2019).
- Steward, C.D.; Rasheed, J.K.; Hubert, S.K.; Biddle, J.W.; Raney, P.M.; Anderson, G.J.; Williams, P.P.; Brittain, K.L.; Oliver, A.; McGowan, J.E.J.; et al. Characterization of clinical isolates of Klebsiella pneumoniae from 19 laboratories using the National Committee for Clinical Laboratory Standards extended-spectrum beta-lactamase detection methods. J. Clin. Microbiol. 2001, 39, 2864–2872. [Google Scholar] [CrossRef] [PubMed]
- Girlich, D.; Naas, T.; Leelaporn, A.; Poirel, L.; Fennewald, M.; Nordmann, P. Nosocomial spread of the integron-located veb-1-like cassette encoding an extended-pectrum beta-lactamase in Pseudomonas aeruginosa in Thailand. Clin. Infect. Dis. 2002, 34, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, A.; Stemplinger, I.; Jungwirth, R.; Ernst, S.; Casellas, J.M. Sequences of beta-lactamase genes encoding CTX-M-1 (MEN-1) and CTX-M-2 and relationship of their amino acid sequences with those of other beta-lactamases. Antimicrob. Agents Chemother. 1996, 40, 509–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gniadkowski, M.; Palucha, A.; Grzesiowski, P.; Hryniewicz, W. Outbreak of ceftazidime-resistant Klebsiella pneumoniae in a pediatric hospital in Warsaw, Poland: Clonal spread of the TEM-47 extended-spectrum beta-lactamase (ESBL)-producing strain and transfer of a plasmid carrying the SHV-5-like ESBL-encoding gene. Antimicrob. Agents Chemother. 1998, 42, 3079–3085. [Google Scholar] [CrossRef]
- De Champs, C.; Poirel, L.; Bonnet, R.; Sirot, D.; Chanal, C.; Sirot, J.; Nordmann, P. Prospective survey of beta-lactamases produced by ceftazidime- resistant Pseudomonas aeruginosa isolated in a French hospital in 2000. Antimicrob. Agents Chemother. 2002, 46, 3031–3034. [Google Scholar] [CrossRef] [PubMed]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for Interpretation of MICs and Zone Diameters. Version 7.0. 2017. Available online: http://www.eucast.org (accessed on 15 May 2018).
- Skoczyńska, A.; Żabicka, D.; Hryniewicz, W. Rekomendacje Doboru Testów do Oznaczania Wrażliwości Bakterii na Antybiotyki i Chemioterapeutyki 2009. Oznaczanie Wrażliwości Neisseria Meningitidis i Haemophilus Influenzae; National Reference Center for Antimicrobial Susceptibility (KORLD): Warszawa, Poland, 2009. [Google Scholar]
- Hinic, V.; Reist, J.; Schibli, U.; Linnik, J.; Egli, A. Beta-lactam susceptibility in Haemophilus influenzae: The connection between genotype and phenotype. In Proceedings of the 27th ECCMID, Vienna, Austria, April 2017. [Google Scholar]
- Sevillano, D.; Giménez, M.-J.; Cercenado, E.; Cafini, F.; Gené, A.; Alou, L.; Marco, F.; Martínez-Martínez, L.; Coronel, P.; Aguilar, L. Genotypic versus phenotypic characterization, with respect to beta-lactam susceptibility, of Haemophilus influenzae isolates exhibiting decreased susceptibility to beta-lactam resistance markers. Antimicrob. Agents Chemother. 2009, 53, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Cardines, R.; Daprai, L.; Giufre, M.; Torresani, E.; Maria Laura Garlaschi, M.C. Genital carriage of the genus Haemophilus in pregnancy: Species distribution and antibiotic susceptibility. J. Med. Microbiol. 2015, 54, 724–730. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| bla Gene | Primer | Sequence (5′to 3′) | Product Size (bp) | Reference |
|---|---|---|---|---|
| blaTEM-1 | TEM-F | ATTCTTGAAGACGAAAGGGC | 1150 | [35] |
| TEM-R | ACGCTCAGTGGAACGAAAAC | |||
| blaGES | GESf | TTCCATCTCAAGGGATCACC | 890 | [36] |
| GESr | GCGTCAACTATTTGTCCGTG | |||
| blaOXA | OXA-F | AGTGTGTTTAGAATGGTGATC | 813 | [37] |
| OXA-R | GTTAGCGGTAATTTAACCAGATAG | |||
| blaVEB | VEB-F | GTTAGCGGTAATTTAACCAGATAG | 1070 | [38] |
| VEB-R | CGGTTTGGGCTATGGGCAG | |||
| blaCTX-M-1 | P1C | TTAATTCGTCTCTTCCAGA | 1000 | [27,39] |
| P2D | CAGCGCTTTTGCCGTCTAAG | |||
| blaSHV | SHV-A | ACTGAATGAGGCGCTTCC | 300 | [40] |
| SHV-B | ATCCCGCAGATAAATCACC | |||
| blaCMY | CMY-F | CAATGTGTGAGAAGCAGTC | 1432 | [26] |
| CMY-R | CGCATGGGATTTTCCTTGCTG | |||
| blaDHA | DHA-f | AACTTTCACAGGTGTGCTGGGT | 405 | [28] |
| DHA-r | CCGTACGCATACTGGCTTTGC | |||
| blaPER | PER-F | TGACGATCTGGAACCTTT | 900 | [41] |
| PER-R | AACTGCATAACCTACTCC | |||
| blaROB-1 | ROB-f | GGATCAGAGTAATAATTTCTG | 192 | [17] |
| ROB-r | GCCATTGAAAGCAAGTTTCAACGG | |||
| pbp3-BLN | BLN-F | GTCACACCACGGTTACTTGAA | 465 | [19] |
| BLN-R | CCCGCAGTAAATGCCACATATTTC | |||
| pbp3-INT | INT-F | GATACTACGTCCTTTAAATTAAGCG | 554 | [19] |
| INT-R | CCCGCAGTAAATGCCACATATTTC |
| Phenotype | Description |
| BLNAS | beta-lactamase-negative cefinase-negative ampicillin-susceptible isolate |
| BLNAI | beta-lactamase negative cefinase-negative isolate with reduced susceptibility to ampicillin |
| low-BLNAR | low-level BLNAR isolate; beta-lactamase-negative ampicillin-resistant isolate with ampicillin MICs in the range of 0.5–2.0 mg/L |
| BLNAR | beta-lactamase-negative ampicillin-resistant isolate with ampicillin MICs ≥ 2.0 mg/L |
| BLNBR | beta-lactamase negative cefinase-negative isolate resistant to one or more beta-lactams (benzylpenicillin, ampicillin, cephalosporins, or carbapenems) |
| BLPAS | beta-lactamase-positive cefinase-negative ampicillin-susceptible isolate |
| BLPAI | beta-lactamase-positive cefinase-negative isolate with reduced susceptibility to ampicillin |
| BLPAR | beta-lactamase-positive cefinase-positive ampicillin- and benzylpenicillin-resistant amoxicillin-clavulanic acid-susceptible isolate |
| BLPACR | beta-lactamase-positive cefinase-negative ampicillin-clavulanic acid-, ampicillin-, or benzylpenicillin-resistant isolate |
| Genotype | Description |
| gBLNAS | isolate negative for beta-lactamase genes ampicillin-susceptible without any amino acid substitutions in ftsI gene |
| gBLNAR | beta-lactamase-negative ampicillin-resistant isolate positive for β-lactamase genes with ftsI gene mutations: subgroup I—substitution of Arg-517→His-517 (Arg-517-His); II—substitution of Arg-526→Lys-526 (Arg-526-Lys); IIa—substitution at the position of 526 except Ala-502; IIb—substitution of Val-502→Ala-502 (Val-502-Ala); IIc—substitution of Thr-502→Ala-502 (Thr-502-Ala); IId—substitution of Val-449→Ile-449 (Val-449-Ile); III—substitutions of three amino acids Met-377→Ile-377, Ser-385→Thr-385 and Leu-389→Phe-389 with addition of Asn-526-Lys |
| low-gBLNAR | low-level gBLNAR beta-lactamase-negative isolate negative for β-lactamase genes isolate: subgroup I—without amino acid substitution; II—substitution at the Lys-526 position in ftsI gene |
| gBLPAR | isolate positive for beta-lactamase genes ampicillin-resistant without any amino acid substitutions in ftsI gene |
| gBLPACR | isolate positive for beta-lactamase genes ampicillin-resistant with ftsI gene mutations: subgroup I—substitutions of Arg-517-His and Arg-526-Lys; II—substitutions of Met-377-Ile, Ser-385-Thr, Leu-389-Phe, and Asn-526-Lys |
| gBLPBS | isolate positive for beta-lactamase genes with ftsI gene mutations: subgroup I—substitution at the Lys-526 position; II—substitutions at the Thr-385 and Lys-526 positions |
| Phenotypical Method | Sensitivity (%) | Specificity (%) | PPV 1 (%) | NPV 2 (%) |
|---|---|---|---|---|
| cefinase test | 52.25 | 100.0 | 100.0 | 34.57 |
| penicillinase test (API NH) | 54.72 | 96.43 | 90.91 | 36.84 |
| amoxicillin–clavulanate | 51.33 | 100.0 | 100.0 | 33.73 |
| benzylpenicillin screen | 90.63 | 100.0 | 100.0 | 82.35 |
| No. | Isolate Name | cef 1 | Resistance Pattern | MICAm 9 (mg/L) | Beta-Lactamase Gene | ftsI10 | ftsI Substitution | Phenotype | Genotype | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TEM-1 | GES | OXA | VEB | SHV | CMY | DHA | Thr-385 11/Lys-526 12 | Lys-526 | ||||||||
| beta-lactamase-positive isolates | ||||||||||||||||
| 1. | 2AU | - | Cxm 3 Ctx 4 | - | x | x | BLPAS | gBLPAS | ||||||||
| 2. | 2BU | - | Am 2 Cxm Ctx | 3.0 | x | + | + | BLPACR | gBLPACR I | |||||||
| 3. | 2CU | - | Am Sam 8 Cxm | 2.0 | x | + | + | BLPACR | gBLPACR I | |||||||
| 4. | 5BU | - | Am Sam | 3.0 | x | BLPACR | gBLPAR | |||||||||
| 5. | 11BU | - | Cxm Ctx | - | x | + | + | BLPAS | gBLPBS I | |||||||
| 6. | 23BU | - | Am Cxm Ctx | 3.0 | x | + | + | BLPACR | gBLPACR I | |||||||
| 7. | 24AU | - | Am Cxm Ctx | 2.0 | x | x | + | + | BLPACR | gBLPACR I | ||||||
| 8. | 27CU | - | Ctx | - | x | + | + | BLPAS | gBLPBS I | |||||||
| 9. | 28BU | - | Ctx | - | x | + | + | + | BLPAS | gBLPBS II | ||||||
| 10. | 28CU | - | Ipm 5 | - | x | + | + | BLPAS | gBLPBS I | |||||||
| 11. | 50AU | - | Cxm | - | x | + | + | BLPAS | gBLPBS I | |||||||
| 12. | 50CU | - | Am | 1.0 | x | BLPAR | gBLPAR | |||||||||
| 13. | W1HB | - | Am | 1.0 | x | x | x | x | BLPAR | gBLPAR | ||||||
| 14. | W1HE | - | Am | 1.5 | x | BLPAR | gBLPAR | |||||||||
| 15. | W4HB | - | Cxm Ctx Ipm Mem 6 | - | x | + | + | BLPAS | gBLPBS I | |||||||
| 16. | W4HC | + | Am AmC 7 Cxm Ctx Ipm Mem | 32.0 | x | + | + | BLPBR | gBLPACR I | |||||||
| 17. | W5HD | + | Am Cxm | 6.0 | x | + | + | BLPAR | gBLPACR I | |||||||
| 18. | W5HP | + | Am AmC Cxm Ctx | 1.0 | x | x | + | + | BLPBR | gBLPACR I | ||||||
| 19. | W6HB | - | Am Cxm | 6.0 | x | BLPAR | gBLPAR | |||||||||
| 20. | W7HC | - | Am Mem | 1.0 | x | + | + | BLPACR | gBLPACR I | |||||||
| 21. | W12HB | - | Am Cxm Ctx | 1.0 | x | BLPAR | gBLPAR | |||||||||
| 22. | IM 1GB | - | Am Cxm Ctx Ipm Mem | 0.5 | x | x | x | x | x | + | + | BLPACR | gBLPACR I | |||
| 23. | IM 2GB | - | Cxm | - | x | BLPAS | gBLPAS | |||||||||
| 24. | IM 4GB | + | Am | 0.75 | x | x | x | BLPAR | gBLPAR | |||||||
| 25. | IM 5GB | - | Cxm | - | x | BLPAS | gBLPAS | |||||||||
| 26. | IM 5GC | - | Am Cxm Ctx | 0.5 | x | + | + | BLPACR | gBLPACR II | |||||||
| 27. | IM 6GB | - | Cxm Ctx Mem | - | x | BLPAS | gBLPAS | |||||||||
| 28. | IM 6NLB | - | Cxm Ipm | - | x | x | BLPAS | gBLPAS | ||||||||
| 29. | IM 9GB | - | Am | 6.0 | x | BLPAR | gBLPAR | |||||||||
| 30. | IM 9GE | - | Cxm | - | x | x | x | BLPAS | gBLPAS | |||||||
| 31. | IM 10GB | - | Am | 6.0 | x | x | x | BLPAR | gBLPAR | |||||||
| 32. | IM 12NC | - | Cxm | - | x | x | BLPAS | gBLPAS | ||||||||
| 33. | IM 12GB | - | Cxm Ctx | - | x | BLPAS | gBLPAS | |||||||||
| 34. | IM 14GC | - | Ctx | - | x | x | x | BLPAS | gBLPAS | |||||||
| 35. | IM 18GA | - | Am | 3.0 | x | BLPAR | gBLPAR | |||||||||
| 36. | IM 20GB | - | Am | 1.0 | x | BLPAR | gBLPAR | |||||||||
| beta-lactamase-negative ftsI-positive isolates | ||||||||||||||||
| 1. | 10BU | - | Am Sam Cxm Ctx | 2.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 2. | 11AU | - | Am Cxm | 3.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 3. | 23CU | - | Am Cxm Ctx | 3.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 4. | 24GU | - | Am Cxm Ctx | 6.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 5. | 25BU | - | Am Cxm Ctx | 8.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 6. | 39CU | + | Am Cxm Ctx | 3.0 | + | + | BLPAR | gBLNAR II | ||||||||
| 7. | W1HC | - | Am | 0.8 | + | + | low-BLNAR | low-gBLNAR II | ||||||||
| 8. | W2HA | - | Am Cxm Ctx | 3.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 9. | W3HA | - | Am Cxm Ctx | 3.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 10. | W3HB | - | Am Cxm Ctx | 3.0 | + | + | BLNAR | gBLNAR II | ||||||||
| 11. | IM 18GB | - | Am | 2.0 | low-BLNAR | low-gBLNAR I | ||||||||||
| beta-lactamase-negative ftsI-negative isolates | ||||||||||||||||
| 1. | 4AU | - | Ipm | - | BLNAS | gBLNAS | ||||||||||
| 2. | 6BU | - | Cxm Ctx | - | BLNAS | gBLNAS | ||||||||||
| 3. | 7AU | - | Ctx | - | BLNAS | gBLNAS | ||||||||||
| 4. | 10AU | - | Ipm | - | BLNAS | gBLNAS | ||||||||||
| 5. | 22AU | - | Ctx Mem | - | BLNAS | gBLNAS | ||||||||||
| 6. | 25CU | - | Cxm Mem | - | BLNAS | gBLNAS | ||||||||||
| 7. | 26CU | - | Ctx | - | BLNAS | gBLNAS | ||||||||||
| 8. | 27BU | - | Ctx | - | BLNAS | gBLNAS | ||||||||||
| 9. | 43AU | - | Ctx | - | BLNAS | gBLNAS | ||||||||||
| 10. | 47BU | - | Ctx | - | BLNAS | gBLNAS | ||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrzejczuk, S.; Kosikowska, U.; Chwiejczak, E.; Stępień-Pyśniak, D.; Malm, A. Prevalence of Resistance to β-Lactam Antibiotics and bla Genes Among Commensal Haemophilus parainfluenzae Isolates from Respiratory Microbiota in Poland. Microorganisms 2019, 7, 427. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100427
Andrzejczuk S, Kosikowska U, Chwiejczak E, Stępień-Pyśniak D, Malm A. Prevalence of Resistance to β-Lactam Antibiotics and bla Genes Among Commensal Haemophilus parainfluenzae Isolates from Respiratory Microbiota in Poland. Microorganisms. 2019; 7(10):427. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100427
Chicago/Turabian StyleAndrzejczuk, Sylwia, Urszula Kosikowska, Edyta Chwiejczak, Dagmara Stępień-Pyśniak, and Anna Malm. 2019. "Prevalence of Resistance to β-Lactam Antibiotics and bla Genes Among Commensal Haemophilus parainfluenzae Isolates from Respiratory Microbiota in Poland" Microorganisms 7, no. 10: 427. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100427