Phylogenetic Analysis of Lednice Orthobunyavirus

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
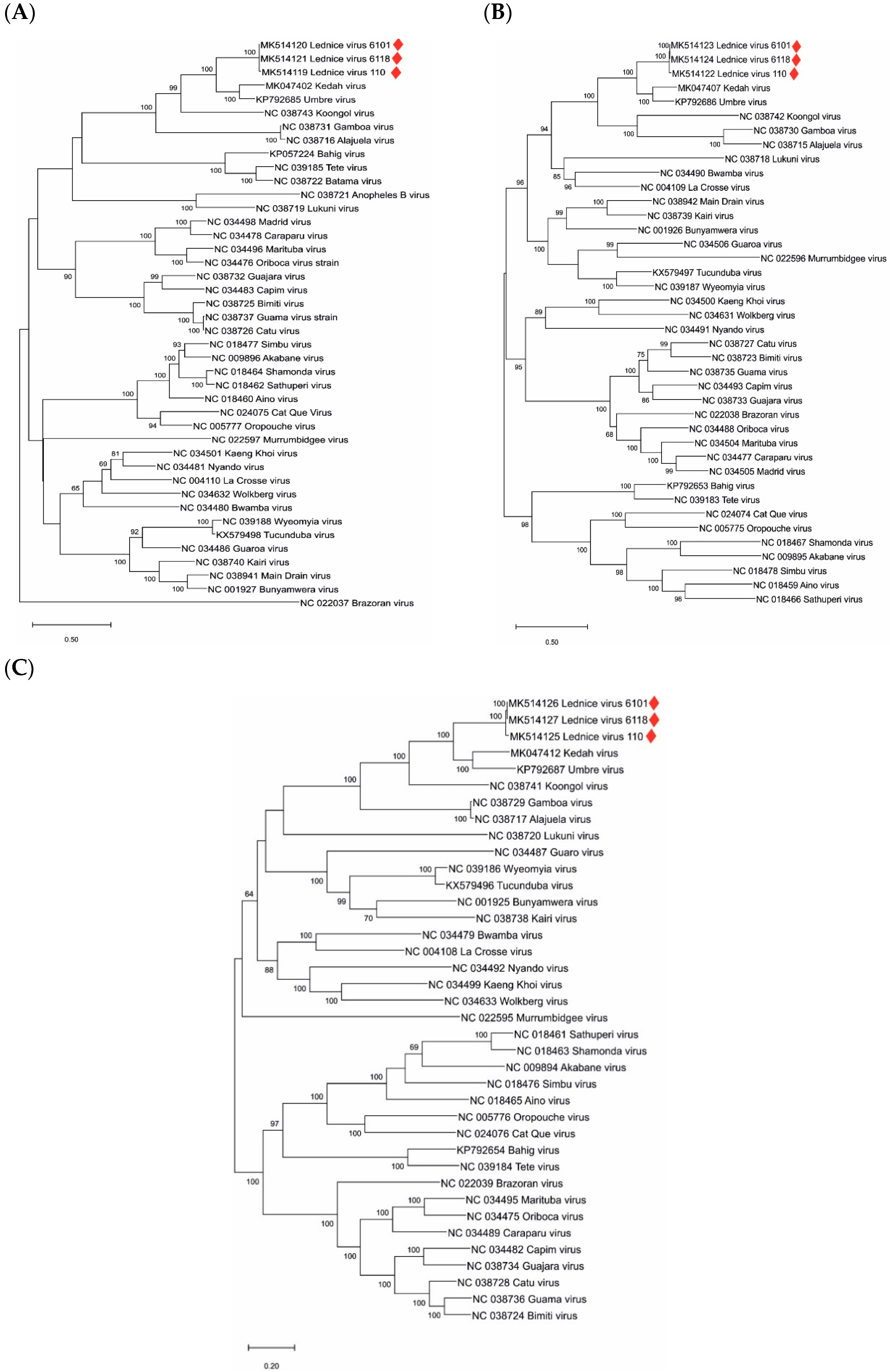
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Danielová, V.; Hájková, Z.; Kolman, J.M.; Málkova, D.; Minár, J.; Smetana, A. Results of the virological examination of mosquitoes in southern Moravia in 1962–1964. Cesk Epidemiol. Mikrobiol. Imunol. 1966, 15, 178–184. [Google Scholar] [PubMed]
- Málková, D.; Danielová, V.; Minář, J.; Rosicky, B.; Casals, J. Isolation of Yaba 1 arbovirus in Czechoslovakia. Acta Virol. 1972, 16, 93. [Google Scholar] [PubMed]
- Van Regenmortel, M.H.V.; Fauquet, C.M.; Bishop, D.H.L.; Carstens, E.B.; Estes, M.K.; Lemon, S.M.; Maniloff, J.; Mayo, M.A.; McGeoch, D.J.; Pringle, C.R.; et al. Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses; Academic Press: San Diego, CA, USA, 2000; p. 608. [Google Scholar]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2017, 46, D708–D717. [Google Scholar] [CrossRef] [PubMed]
- Kolman, J.M.; Minář, J.; Horak, I. Serologic examination of birds from the area of southern moravia for the presence of antibodies against arboviruses of the groups Alfa, Flavo, Bunyamwera Supergroup, and the virus Yaba 1-Lednice 110. 1. Domestic fowls. Zentralbl. Bakteriol. Orig. Reihe A: Med. Mikrobiol. Parasitol. 1975, 23, 279–287. [Google Scholar]
- Wojta, J.; Aspöck, H. Untersuchungen über die Möglichkeit der Einschleppung durch Stechmücken übertragener Arboviren durch Vögel nach Mitteleuropa. Mitt. Österr. Ges. Tropenmed. Parasitol. 1982, 4, 85–98. [Google Scholar]
- Malková, D.; Danielová, V.; Holubová, J.; Marhoul, Z. Less Known Arboviruses of Central Europe: A New Arbovirus Lednice; Academia Publishing House of the Czechoslovak Academy of Science: Prague, Czech Republic, 1986; p. 75. ISSN 0069-228X. [Google Scholar]
- Schwaiger, K.; Bauer, J. Epidemiology of emerging and resurging vector-borne diseases with special attention to climate change in Germany (Review). Berl. Münch. Tierärztl. Wochenschr. 2009, 122, 141–160. [Google Scholar]
- Málková, D.; Kolman, J.M. Yaba 1-Lednice 110 virus proliferation in chicken embryo. Cesk. Epidemiol. Mikrobiol. Imunol. 1975, 24, 225–230. [Google Scholar]
- Marhoul, Z.; Danielová, V.; Holubová-Krobová, J.; Málková, D. Cultivation of Lednice (Yaba 1) virus in goose, duck, and chick embryo cells. Acta Virol. 1976, 20, 499–505. [Google Scholar]
- Danielová, V.; Málková, D. Studies on viremia and antibody formation in ducklings and goslings after experimental infection wih Lednice (Yaba 1) virus. Folia Parasitol. (Praha) 1976, 23, 367–372. [Google Scholar]
- Málková, D.; Danielová, V.; Lím, D. Experimental infection of black-headed gulls (Larus ridibundu L.) and coots (Fulica atra L.) with Lednice (M’Poko) virus. Folia Parasitol. (Praha) 1979, 26, 85–88. [Google Scholar]
- Málková, D.; Danielová, V.; Viktora, L.; Holubová-Krobová, J. Experimental infection of Macaca mulatta monkeys with Lednice (Yaba 1) virus. Acta Virol. 1976, 20, 226–231. [Google Scholar] [PubMed]
- Danielová, V. Relationships of Mosquitoes to Ťahyňa Virus as Determinant Factors of Its Circulation in Nature; Academia Publishing House of the Czechoslovak Academy of Sciences: Prague, Czech Republic, 1992; p. 104. ISSN 0577-3652. [Google Scholar]
- Kramer, L.D.; Hardy, J.L.; Houk, E.J.; Presser, S.B. Characterization of the mesenteronal infection with Western equine encephalomyelitis virus in an incompetent strain of Culex tarsalis. Am. J. Trop. Med. Hyg. 1989, 41, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Hardy, J.L.; Reeves, W.C.; Presser, S.B.; Milby, M.M.; Meyer, R.P. Persistence of mosquito-borne viruses in Kern County, California, 1983–1988. Am. J. Trop. Med. Hyg. 1990, 43, 419–437. [Google Scholar] [CrossRef] [PubMed]
- Jakob, W.L.; Davis, T.; Francy, D.B. Occurrence of Culex erythrothorax in southeastern Colorado and report of virus isolations from this and other mosquito species. J. Am. Mosq. Control. Assoc. 1989, 5, 534–536. [Google Scholar] [PubMed]
- Yadav, P.D.; Chaubal, G.Y.; Shete, A.M.; Mourya, D.T. A mini-review of bunyaviruses recorded in India. Indian J. Med. Res. 2017, 145, 601–610. [Google Scholar]
- Sudeep, A.B.; Ghodke, Y.S.; George, R.P.; Ingale, V.S.; Dhaigude, S.D.; Gokhale, M.D. Vectorial capacity of Culex gelidus (Theobald) mosquitoes to certain viruses of public health importance in India. J. Vector Borne Dis. 2015, 52, 153–158. [Google Scholar]
- Yadav, P.D.; Mishra, A.C.; Mourya, D.T. Molecular characterization of Umbre virus (Bunyaviridae). Virol. J. 2008, 5, 115. [Google Scholar] [CrossRef]
- Boiro, I.; Lomonossov, N.N.; Fidarov, F.M.; Murzine, S.V.; Linev, N.B.; Camara, L.B.; Bah, A. Isolation of M’Poko virus (Turlock group, Bunyaviridae) from Culex cinereus mosquitoes in the Republic of Guinea. Bull. Soc. Pathol. Exot. Filiales. 1985, 78, 452–455. [Google Scholar]
- Málková, D.; Danielová, V.; Minář, J.; Ryba, J. Virological investigations of mosquitoes in some biotopes of southern Moravia in summer season 1972. Folia Parasitol. (Praha) 1974, 21, 363–372. [Google Scholar]
- Mihalov-Kovács, E.; Martella, V.; Lanave, G.; Bodnar, L.; Fehér, E.; Marton, S.; Kemenesi, G.; Jakab, F.; Bányai, K. Genome analysis of canine astroviruses reveals genetic heterogeneity and suggests possible inter-species transmission. Virus Res. 2017, 232, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Djikeng, A.; Halpin, R.; Kuzmickas, R.; Depasse, J.; Feldblyum, J.; Sengamalay, N.; Afonso, C.; Zhang, X.; Anderson, N.G.; Ghedin, E.; et al. Viral genome sequencing by random priming methods. BMC Genom. 2008, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Botting, C.H.; Li, P.; Niglas, M.; Brennan, B.; Shirran, S.L.; Szemiel, A.M.; Elliott, R.M. Bunyamwera orthobunyavirus glycoprotein precursor is processed by cellular signal peptidase and signal peptide peptidase. Proc. Natl. Acad. Sci. USA 2016, 113, 8825–8830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briese, T.; Williams, D.T.; Kapoor, V.; Diviney, S.M.; Certoma, A.; Wang, J.; Johansen, C.A.; Chowdhary, R.; Mackenzie, J.S.; Lipkin, W.I.; et al. Analysis of arbovirus isolates from Australia identifies novel Bunyaviruses including a Mapputta group virus from Western Australia that links Gan Gan and Maprik viruses. PLoS ONE 2016, 11, e0164868. [Google Scholar] [CrossRef]
- Müller, R.; Poch, O.; Delarue, M.; Bishop, D.H.; Bouloy, M. Rift valley fever virus L segment: correction of the sequence and possible functional role of newly identified regions conserved in RNA-dependent polymerases. J. Gen. Virol. 1994, 75, 1345–1352. [Google Scholar] [CrossRef]
- Reguera, J.; Weber, F.; Cusack, S. Bunyaviridae RNA polymerases (L-protein) have an N-terminal, influenza-like endonuclease domain, essential for viral cap-dependent transcription. PLoS Pathog. 2010, 6, e1001101. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Beaty, B.J.; Blair, C.D.; Brault, A.C.; Dobler, G.; Drebot, M.A.; Haddow, A.D.; Kramer, L.D.; LaBeaud, A.D.; Monath, T.P.; et al. Bunyavirus taxonomy: Limitations and misconceptions associated with the current ICTV criteria used for species demarcation. Am. J. Trop. Med. Hyg. 2018, 99, 11–16. [Google Scholar] [CrossRef]
- Lennette, E.H.; Ota, M.I.; Fujimoto, F.Y.; Wiener, A.; Loomis, E.C. Turlock virus: A presumably new arthropod-borne virus; isolation and identification. Am. J. Trop. Med. Hyg. 1957, 6, 1024–1035. [Google Scholar] [CrossRef]
- Palya, V.; Kovács, E.W.; Marton, S.; Tatár-Kis, T.; Felföldi, B.; Forró, B.; Domán, M.; Bányai, K. Novel orthobunyavirus causing severe kidney disease in broiler chickens, Malaysia, 2014–2017. Emerg. Infect. Dis. 2019, 25, 1110–1117. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berčič, R.L.; Bányai, K.; Růžek, D.; Fehér, E.; Domán, M.; Danielová, V.; Bakonyi, T.; Nowotny, N. Phylogenetic Analysis of Lednice Orthobunyavirus. Microorganisms 2019, 7, 447. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100447
Berčič RL, Bányai K, Růžek D, Fehér E, Domán M, Danielová V, Bakonyi T, Nowotny N. Phylogenetic Analysis of Lednice Orthobunyavirus. Microorganisms. 2019; 7(10):447. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100447
Chicago/Turabian StyleBerčič, Rebeka Lucijana, Krisztián Bányai, Daniel Růžek, Enikő Fehér, Marianna Domán, Vlasta Danielová, Tamás Bakonyi, and Norbert Nowotny. 2019. "Phylogenetic Analysis of Lednice Orthobunyavirus" Microorganisms 7, no. 10: 447. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100447