The Effect of Maternal Postpartum Practices on Infant Gut Microbiota: A Chinese Cohort Study



1
Affiliation Wuhan University School of Health Sciences, Wuhan University, 169 Donghu Road, Wuhan 430071, China
2
Affiliation Emory University Nell Hodgson Woodruff School of Nursing, 1520 Clifton Road, Atlanta, GA 30322, USA
*
Author to whom correspondence should be addressed.
Microorganisms 2019, 7(11), 511; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110511
Submission received: 15 October 2019
/
Revised: 25 October 2019
/
Accepted: 28 October 2019
/
Published: 30 October 2019
(This article belongs to the Special Issue Feature Papers in Gut Microbiota)
Abstract
:(1) Background: The human gut microbiota at early life is shaped by numerous factors, especially factors from mothers, which have huge influence on infants’ gut microbiotas. The aim of this study was to investigate the effect of maternal adherence to Chinese traditional postpartum practices of “doing the month” on the development of infant gut microbiota at 6-month postpartum. (2) Methods: A cohort of 62 Chinese women at late pregnancy was recruited from a tertiary general hospital in a central region of China. The participants and their babies were followed up to 6 months postpartum. Finally, 50 mother-infant dyads were enrolled in the study. Women’s adherence to the traditional postpartum practices was measured by adherence to doing the month practices (ADP). Infant fecal samples were collected at six months of age and were analyzed using 16S rRNA V3 and V4 gene region sequences. (3) Results: Ruminococcus gnavus was significantly less abundant in infants whose mothers had a better adherence to the traditional postpartum practices of “doing the month.” Infants receiving Clostridium-butyricum during the first month after delivery had a significant dominance of Escherichia/Shigella. (4) Conclusions: Adherence to the traditional postpartum practices of “doing the month” can impact an infant’s gut microbiota at 6 months of age. Infants receiving probiotics during the first month after delivery had a significant dominance of opportunistic pathogens.
1. Introduction
The gut microbiota is the second most important genome of the human body. It represents a large microbial ecosystem which hosts trillions of microbial cells [1]. The gut microbiota provides essential functionality for the human body, including biosynthesis, metabolism, maturation and homeostasis of the human immune system, host cell proliferation, and vascularization and neurologic signaling [2]. A dysbiotic gut microbiota is associated with a variety of diseases and chronic conditions, such as diabetes [3], obesity [4], and allergenic diseases [5].
Recent research has uncovered the importance of the gut microbiota in human health and has attempted to understand the gut microbiota from the beginning of life. The ecological theory revealed that the priority effects (e.g., the order and timing of gut microbiota arrival) and microbial transmission (e.g., in utero transmission, infant delivery, and feeding modalities) determine subsequent development and health in later life [6,7]. Studies have reported the existence of a mother-to-infant gut microbiota transformation during the neonatal period [8,9,10]. A fetus acquires its microbiota in utero from its contact with maternal amniotic fluid [11], placenta [12,13], and umbilical cord blood [14]. After birth, the mother is still the main source of the infant’s gut microbiota, which includes maternal breast milk, skin, and the oral cavity [15]. Thus, various maternal factors can impact the development of the infant’s gut microbiota. Increasing evidence has demonstrated that maternal pre-pregnancy body mass index (BMI) and gestational weight gain (GWG) have direct impacts on infants. Excessive GWG, being overweight, or obesity before pregnancy regulated by maternal gut microbiota increase the risk of fetal gut microbiota imbalance, perinatal mortality, obesity, and metabolic syndromes [16,17]. Maternal stress during pregnancy affects the mother’s, as well as the infant’s, gut microbiota and increases the risk of anxiety in offspring [18,19]. Mueller et al. [20] also reported that the infant’s gut microbiota was affected through maternal antibiotic exposure during pregnancy, leading to obesity later in the infant’s life. Furthermore, a dysbiotic colonization of infant gut microbiota can result in detrimental healthcare outcomes in infants, such as higher rates of asthma, allergy, and gastrointestinal issues in later life [21].
Most recently, researchers have started to investigate the effect of human lifestyles and behaviors on gut microbiota in preclinical and clinical studies, including the study of diet patterns, physical activity, drinking, and smoking. Multiple studies suggest that diet and other lifestyle behaviors, such as physical activity, are primary factors determining the development and composition of the human gut microbiota [22,23,24]. In China, during the first month after delivery, postpartum women routinely adhere to the traditional maternal postpartum practices called “doing the month,” which are believed to restore body harmony and health, and prevent future illness [25]. “Doing the month” includes a series of practical behaviors related to the maternal role, such as activity limitations, maintaining warmth, and prescriptions about the types of food to consume or avoid [26]. As a unique set of culture-based postpartum practical behaviors, “doing the month” has been confirmed to lead to a limited maternal diet, poor hygiene, postpartum depression, and other physical discomfort in modern life [26]. Studies have shown that maternal nutritional composition influences the nutritional properties of breast milk [27], which serves as the main resource of microbiota that seeds the infant gut, and this could alter the infant’s gut microbiota to a large extent [28]. In addition, by directly contacting mothers’ skin, poor maternal hygiene may influence the infant gut microbiota [28]. “Doing the month” also leads to changes in maternal psychological health (e.g., depression and anxiety) [29], and this could impact the infant’s gut microbiota’s composition [30] Nevertheless, the effect of adherence to the traditionally practical postpartum behaviors—“doing the month,” on the development of the infant’s gut microbiota is unknown. We hypothesize that a mother’s adherence to the traditionally practical postpartum behaviors of “doing the month” is a major factor that affects the development of the infant’s gut microbiota at 6 months after delivery.
The goals of this study were to: (1) investigate the effect of maternal adherence to traditional Chinese “doing the month” practical behaviors on an infant’s gut microbiota at 6 months; and (2) examine the influence of early life events on the infant’s gut microbiota, including antibiotic exposure, consumption of probiotics, delivery mode, and feeding type. Understanding associations between traditionally maternal postpartum behaviors and the gut microbiota in infants would provide a new angle to promote healthy growth and development in infants.
2. Materials and Methods
2.1. Study design, Setting, and Participants
This study used a prospective maternal-child cohort design. Sixty-two women during their late pregnancy were recruited from an obstetric outpatient clinic in a tertiary hospital of Wuhan University in Wuhan, Hubei Province, China. The study was conducted between 4 March 2017 and 12 April 2018. This study was approved by the Research Ethics Boards of Medical School at Wuhan University (3 March 2017, JKHL2017-03-03). Informed consent statements were signed by all participants.
All women were selected independent of genetic background. The inclusion criteria were (1) pregnant women who planned to deliver a baby in a tertiary hospital of Wuhan University; and (2) a Wuhan resident. Pregnant women with pregnancy complications, women receiving antibiotic treatment during pregnancy, or those with cognitive impairment were excluded. Finally, this study included 50 mother-infant dyads (12 families could not be reached by telephone or email at 6 months postpartum) and 50 fecal samples were collected at home based on the Human Microbiome Project (HMP) protocol [31]. After collection, the fecal samples were transferred in a cooler (+4 °C, 1.5 h on average) and taken to the laboratory where they were frozen at −80 °C before DNA extraction. A total of 50 fecal samples were provided for high quality Illumina sequencing.
2.2. Variables and Measures
The Chinese adherence to doing the month practices (ADP) was used to assess mothers’ ADP behaviors during the one-month postpartum period [25,26]. The score used for assessment ranges from 0 to 108, and a higher score indicates a higher level of adherence. The content validity index was 0.95 and Cronbach’s α was 0.86 [32]. This questionnaire was sent to participants by WeChat software at one month postpartum and women completed the online questionnaire via WeChat software.
Maternal and infant demographic data, GWG, and pre-pregnancy BMI were collected in the hospital based on maternal self-report 24 hours after childbirth. Infant antibiotic exposure, probiotic consumption, mode of delivery, and feeding type were ascertained when collecting samples at participants’ homes, based on maternal self-report. Pre-pregnancy weight was categorized as underweight, normal weight, overweight, or obese using the Asian-Pacific Recommendations [33]. According to the recommended range of the Institute of Medicine (IOM) [34], GWG was categorized as adequate, excessive, or low based on the pre-pregnancy weight.
The newborn consumption of probiotics containing Clostridium butyricum (C. butyricum, at least 75X10000000CFU) was meant to facilitate defecation and prevent jaundice in early neonates (0–1 month after birth). In this study, 35 parents prophylactically gave probiotic products to their healthy newborns despite the absence of physical signs of jaundice; among these infants, none of them developed jaundice during the course of the study. The use of probiotics was collected from the infant’s medical card, with the participants’ permission.
Infant antibiotic exposure was based on maternal self-report at the time of infant fecal sample collection. The antibiotics included cephalosporin and penicillin, which were given with intravenous fluids or orally. There were 16 infants that had antibiotic exposure during the 6 months postpartum.
Infant feeding type and the ADP questionnaire were collected one month after birth, based on mothers’ self-reports. Feeding type was also collected at six months after birth, coinciding with fecal sample collection, based on mothers’ self-reports. We categorized breast milk feeding groups as breast-fed or mixed feeding; infants who were mainly fed formula mixed with little breast milk were categorized as the mixed feeding group. None of the infants were fed solid food when fecal samples were collected.
2.3. DNA Extraction and PCR Amplification
Microbial DNA was extracted from fecal samples using the E.Z.N.A.® soil DNA Kit (Omega Bio-tek, Norcross, GA, USA) according to manufacturer’s protocols. The final DNA concentration and purification were determined by NanoDrop 2000 UV-vis spectrophotometer (Thermo Scientific, Wilmington, DE, USA), and DNA quality was checked by 1% agarose gel electrophoresis. The V3-V4 hypervariable regions of the bacterial 16S rRNA gene were amplified with primers 338F (5′- ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) by thermocycler PCR system (GeneAmp 9700, ABI, Walthma, MA, USA). The PCR reactions were conducted using the following program: 3 min of denaturation at 95 °C, 27 cycles of 30 s at 95 °C, 30 s for annealing at 55 °C, and 45 s for elongation at 72 °C, and a final extension at 72 °C for 10 min. PCR reactions were performed in triplicate using a 20 μL mixture containing 4 μL of 5 × FastPfu Buffer, 2 μL of 2.5 mM dNTPs, 0.8 μL of each primer (5 μM), 0.4 μL of FastPfu Polymerase and 10 ng of template DNA. The resulting PCR products were extracted from a 2% agarose gel and further purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA) and quantified using QuantiFluor™-ST (Promega, Madison, WI, USA) according to the manufacturer’s protocol.
2.4. Illumina MiSeq Sequencing
Purified amplicons were pooled in equimolar quantities and paired-end sequenced (2 × 300) on an Illumina MiSeq platform (Illumina, San Diego, CA, USA) according to the standard protocols by Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China).
2.5. Processing of Sequencing Data
Raw 16S rRNA sequencing data were demultiplexed, quality-filtered by Trimmomatic, and merged by FLASH with the following criteria: (1) the reads were truncated at any site receiving an average quality score <20 over a 50 bp sliding window. (2) Primers were exactly matched, allowing two nucleotide mismatches, and reads containing ambiguous bases were removed. (3) Sequences whose overlaps were longer than 10 bp were merged according to their overlap sequence.
Operational taxonomic units (OTUs) were clustered with a 97% similarity cutoff using UPARSE (version 7.1 http://drive5.com/uparse/), and chimeric sequences were identified and removed using UCHIME. The taxonomy of each 16S rRNA gene sequence was analyzed by RDP Classifier algorithm (http://rdp.cme.msu.edu/) against the Silva (SSU123) 16S rRNA database using a confidence threshold of 70%.
2.6. Statistical Methods
In the analysis, Alpha-diversity metrics were used, including diversity (Shannon, Simpson), richness (Chao, ace, sobs), and evenness (heip). In this study, Shannon indexes on OTU level were analyzed using Mothur (Version 1.30.2; www.mothur.org/). A larger Shannon value indicates the higher community diversity.
Principal coordinate analysis (PCoA) was conducted according to the Bray–Curtis distance matrix calculated using OTU information from each sample. Permutational multivariate analysis of variance (PERMANOVA) was conducted to analyze the explanative degree of different grouping factors on the sample difference at the OTU level using the Bray–Curtis distance matrix. In PERMANOVA, we controlled confounders, including mode of delivery, feeding type, probiotic consumption, antibiotic exposure, and adherence to doing the month behaviors. Beta diversity metrics were analyzed based on mode of delivery, feeding type, probiotics consumption, and antibiotic exposure.
To investigate associations between maternal and infant characteristics and infant gut microbiota compositions, the linear discriminant analysis (LDA) effect size (LEfSe) method was implemented to identify the distinguishing taxa from phylum to genus levels within groups (http://huttenhower.sph.harvard.edu/galaxy/). A taxonomic bar chart and cladograms were used to visualize the results.
Multivariate analysis of general liner model (GLM) was used to adjust for possible confounders and to analyze the impact of significant factors on the diversity and abundance of the infant’s gut microbiota. This model used the significant genera within groups from LEfSe to adjust confounders. Additionally, to evaluate the effect of maternal “doing the month” on infant gut microbiota, all genera were included if they represented at least 20% of the infants and they had a minimum relative abundance of 0.25%. Finally, 21 genera meeting the prevalence cutoffs were included in the bioinformatic analysis. The following maternal characteristics were also included in the analysis: adherence behaviors of “doing the month,” delivery mode (categorical data: natural delivery versus C-section), antibiotic exposure (categorical data: yes versus no), feeding type (categorical data: breast feeding versus mixed feeding), and probiotic consumption (categorical data: yes versus no).
Data regarding the relative abundances of genera were centered log ratio (CLR) transformed before the analysis. The multiple hypothesis tests were adjusted using Benjamini and Hochberg false discovery rate (FDR), and an association below an FDR threshold of 0.05 was considered significant.
3. Results
3.1. Characteristics of Sequencing Results
A total of 2,551,852 usable sequences and 664 OTUs were obtained from 50 infants’ samples using Illumina MiSeq sequencing (Table 1). From these, 1,075,250 high-quality sequences were selected, with an average of 21,505 sequences per sample. In total, 532 OTUs were delineated at a 97% similarity level in infants after data trimming and quality filtering to an equal sequencing depth (43,953 reads per sample). The pan analysis indicated that only a small number of new, shared phylotypes would be expected with additional sequencing. The values of Good’s coverage for observed OTUs was 99.6 ± 0.03% (mean ± SD).
3.2. Microbiota Taxonomy
The bacterial distribution was characterized in terms of the relative taxonomic abundances. A total of 21 phyla, 130 families, and 258 genera were found from 50 infants’ samples at 6 months postpartum. There were 54 genera present in at least 20% of infants’ gut microbiotas, and using 0.25% relative abundance as a cutoff, 21 genera met these conditions (Table S1). Figure 1 shows taxonomic compositions of the dominant bacteria on the genus level (relative abundance >1%). For the relative abundance, there were 14 genera above 1%, and the 10 most abundant genera were Bifidobacterium (38.18%), Escherichia-Shigella (10.63%), Veillonella (8.37%), Clostridium sensu stricto 1(7.64%), Klebsiella (5.37%), Bacteroides (5.3%), Ruminococcus gnavus group (3.29%), Megasphaera (3%), Enterococcus (2.88%), and Lactobacillus (1.72%).
3.3. Microbiotal Diversity
Based on the multivariate analysis of GLM, no significant factors were associated with Shannon’s index (mean ± SD: 1.81 ± 0.45) of infants’ gut microbiotas, controlling for maternal adherence to “doing the month,” feeding type, delivery mode, antibiotic exposure, and probiotic consumption.
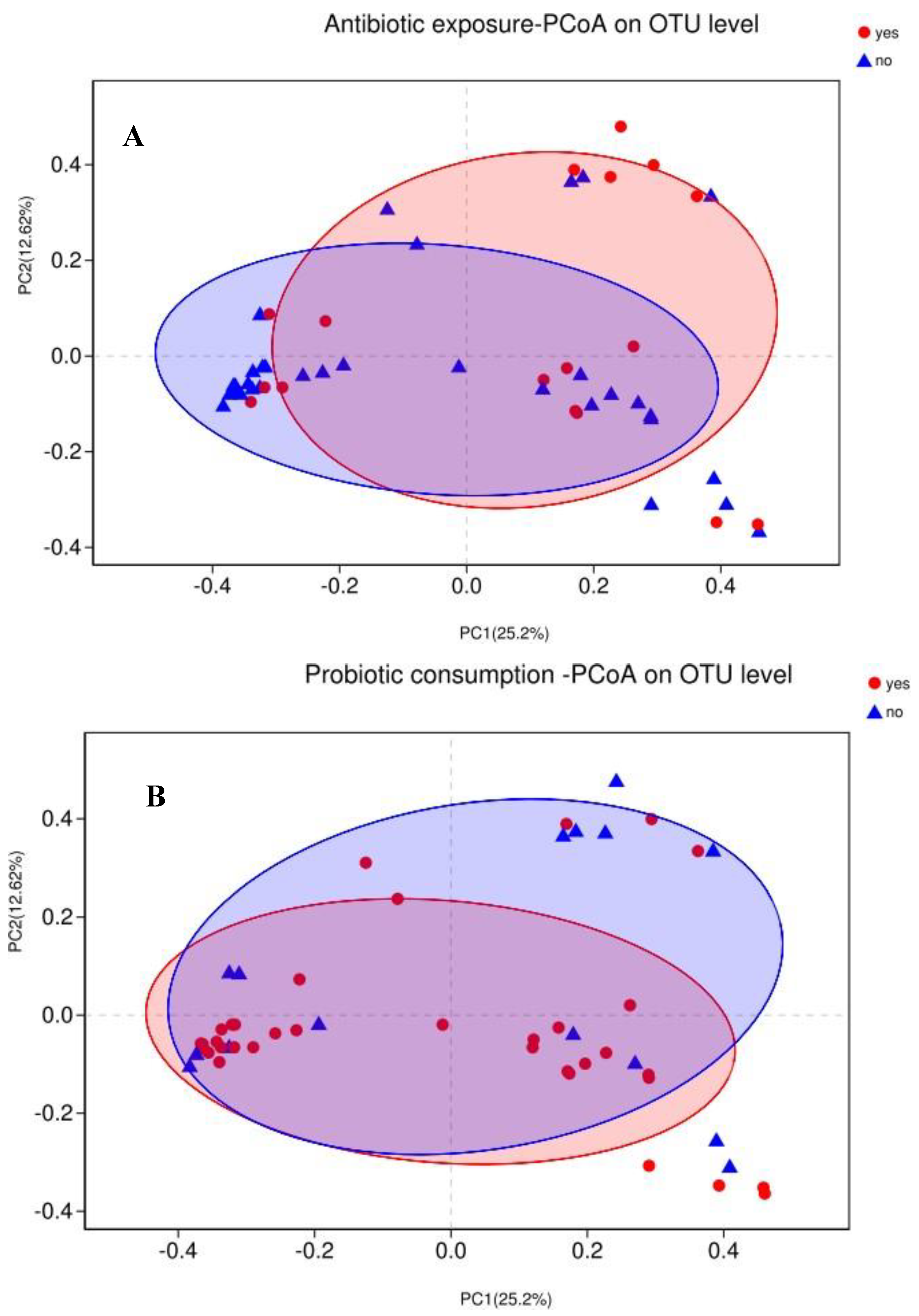
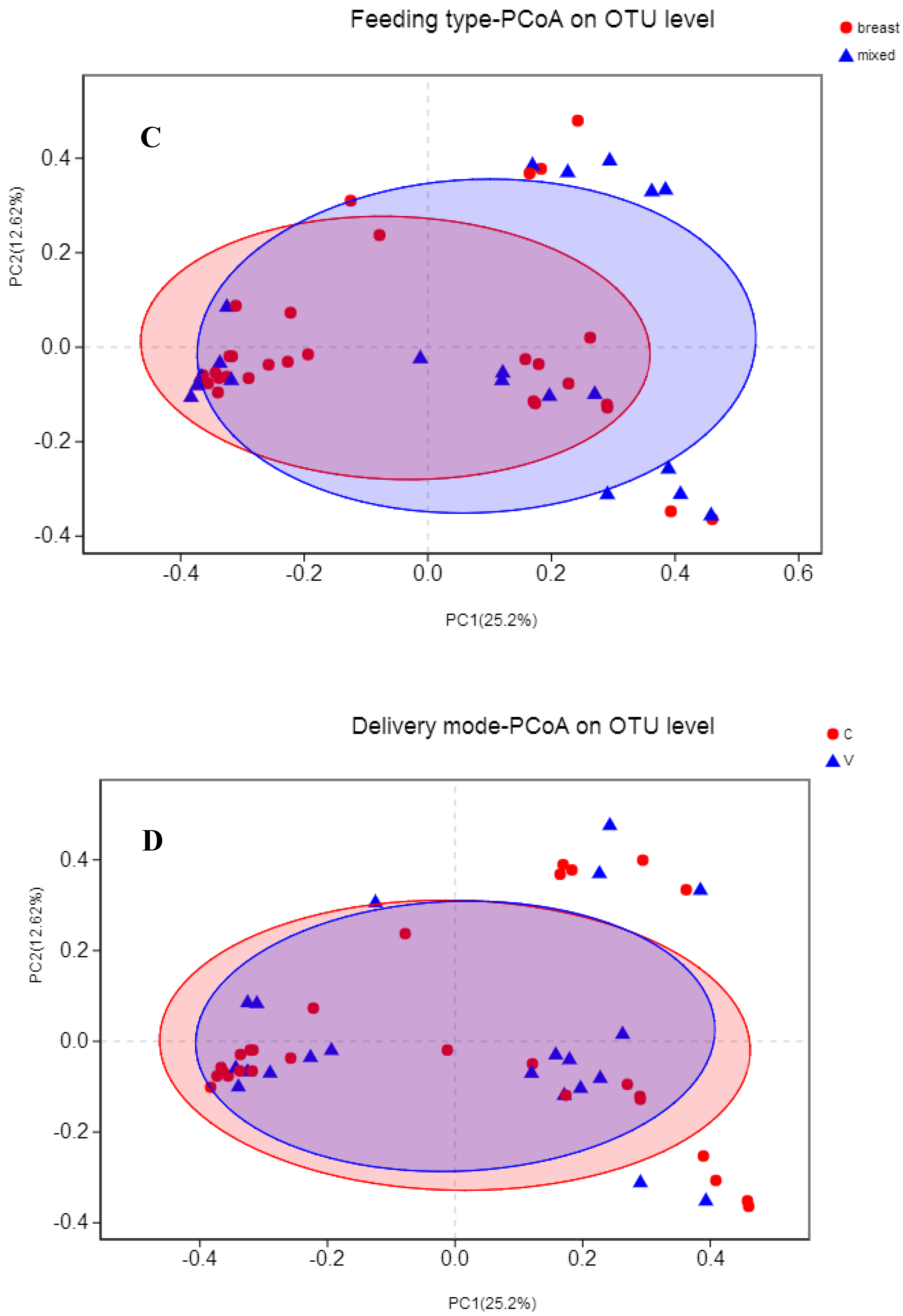
Variations in infants’ gut microbiota compositions among different groups (beta-diversity) were visualized by PCoA based on Bray–Curtis dissimilarity matrices on OTU level (Figure 2). Differences were further tested with PERMANOVA. Not surprisingly, the gut microbiota was not significantly separated based on antibiotic exposure (R2 = 0.03, adjust p = 0.24), probiotic consumption (R2 = 0.03, adjust p = 0.24), feeding type (R2 = 0.02, adjust p = 0.36), or delivery mode (R2 = 0.02, adjust p = 0.56).
3.4. Association between Maternal Adherence to “Doing the Month” and an Infant’s Gut Microbiota at 6 Months Postpartum
In the PERMANOVA analysis, there were no significant differences in maternal adherence to “doing the month” (R2 = 0.03, adjust p = 0.24). Controlling for confounders, we further explored the GLM by analyzing the impact of maternal adherence to “doing the month” on infants’ dominant genus and alpha diversity. According to the presence rate of no less than 20% and at least 0.25% relative abundance cutoffs, there were 21 genera included in the model. Results showed that Ruminococcus gnavus was statistically less abundant in infants whose mothers had higher adherence to the Chinese traditional practices of “doing the month” (β = −43.52, CI = (−72.9, −14.14), p = 0.005, adjust p = 0.012), and Ruminococcus gnavus was the seventh most abundant genus among infants. Intestinibacter (β = 0.02, CI = (0.001, 0.039), p = 0.035, adjust p = 0.376) and Tyzzerella (β = 0.019, CI = (0.0001, 0.037), p = 0.047, adjust p = 0.376) were positively correlated with maternal adherence to “doing the month.” However, after FDR adjusting, there were no significant relationships between maternal adherence to “doing the month” and an infant’s gut microbiota at 6 months postpartum.
3.5. Association between Events in Early Life and an Infant’s Gut Microbiota at 6 Months Postpartum
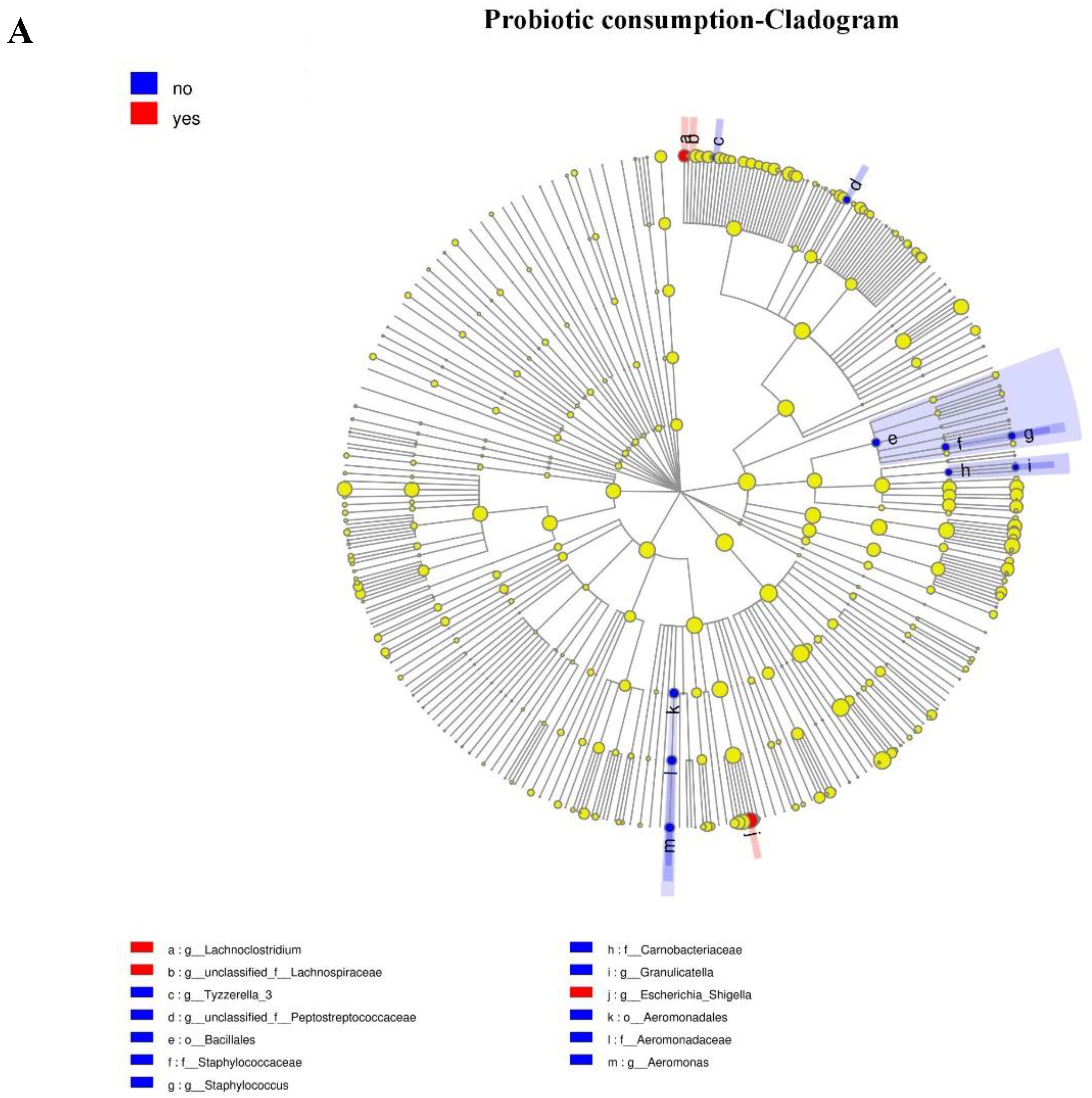
Because of the absence of any clear clusters in PCoA for gut microbiota within groups, the LEfSe method was implemented to identify the distinguishing taxa from the phylum to genus level within groups including probiotic consumption, delivery mode, feeding type, and antibiotic exposure (Figure 3). Furthermore, the significantly different genera were incorporated into the GLM to control confounders. Results showed that infants who consumed probiotics had a dominance of Escherichia-Shigella (β = 287.44, CI = (172.79, 558.09), p = 0.038, adjust p = 0.042), as the second most abundant genus. There were no differences in genera abundance within delivery mode (natural delivery versus C-section), antibiotic exposure (yes versus no), or feeding type (breast versus mixed) groups.
4. Discussion
Recent evidence has shown that the acquisition and development of an infant’s microbiota are key to establishing a healthy host-microbiotal symbiosis. The maternal microbial reservoir plays a crucial role in this process [15,35,36]. In this study, we found that maternal adherence to traditionally practical behaviors of “doing the month” in the postpartum period was associated with a lower abundance of Ruminococcus gnavus genus in the infant’s gut microbiota. Ruminococcus gnavus, which has well defined roles in intestinal mucus degradation and immune function, increases the probability of infectious diarrhea [37]. Specifically, the Lachnospiraceae family, which includes the genus Ruminococcus spp., has been proven to be associated with protection against C. difficile infections [38] and obesity [39].
We hypothesized that maternal adherence to “doing the month” might influence the infant gut microbiota by three pathways. Firstly, women with higher maternal adherence to traditional practices of “doing the month” generally had poor personal hygiene and did not bathe or wash their hair during the one-month postpartum period [29]. A recent study suggested that 10.3% of infants’ gut microbiotas were received from mothers’ areolar skin during the first 30 days of life [28]. By directly contacting with mothers’ skin and surrounding environments, higher maternal adherence to traditional practices of “doing the month” might decrease Ruminococcus spp. in infants due to poor maternal hygiene. Secondly, infant gut microbiota could be affected through breast-feeding. During the first month postpartum, higher adherence to traditional practices of “doing the month” are always accompanied with higher probability of a single diet [26], sleeping problems [32], and decreased immunity [40], all of which could modulate maternal breast milk directly or indirectly by the hypothalamic–pituitary–adrenal (HPA) axis. Breast milk is the main resource seeding infants’ gut microbiotas [28], which could alter abundance of Ruminococcus spp. in infants’ gut microbiotas. In addition, during the first month after birth, postpartum women spend a large amount of daily time in bed. The restriction of physical activity is known to cause musculoskeletal and cardiovascular deconditioning [32,41]. For maternal psychological health, studies suggested that longer adherence to traditional practices of “doing the month” increased stress and postpartum depression [42,43]. Higher adherence to “doing the month” makes postpartum women prone to stress and depression [29], and maternal psychological health may affect an infant’s gut microbiota’s composition [30]. In a longitudinal study, Zijlmans et al. prospectively investigated the relationship between exposure to prenatal stress and infants’ gut microbiotas at 3 months of age; in their study, 58 women in their third trimester were assessed for prenatal stress, and their infants’ gut microbiotas were assessed at 3 months of age. They found that increasing abundance of Proteobacteria, as well as decreased lactic acid bacteria and Bifidobacteria, were associated with infants being born to mothers with high levels of reported stress. The study was the first to prove that maternal stress could modulate an infant’s gut microbiota [30].
In this study, results showed that infant gut microbiota did not show any difference in breast milk infants and mixed feeding infants. Evidence has shown that feeding type is a major factor determining early microbial colonization, influencing the neonatal gut microbiota composition and gastrointestinal function [34]. Studies have reported that the gut microbiotas of breast-fed infants contain higher levels of Lactobacilli and Bifidobacteria and lower levels of potential pathogens compared with those with formula feeding [44,45,46], which was inconsistent with our findings. A likely explanation was that the majority of infants in this study were breast-fed during the first month; however, most were formula-fed for the remainder of our study. Another study also confirmed that infants fed breast milk exclusively showed the same composition of gut microbiota compared with those receiving formula and breast milk [47].
We explored the effect of Clostridium butyricum consumption at early life on infant gut microbiota. Of the 35 healthy infants who were prophylactically given Clostridium butyricum to prevent jaundice, none developed jaundice over the course of this study; we found that these infants had a dominance of Escherichia/Shigella compared with those not receiving Clostridium butyricum. Clostridium butyricum is known as a butyrate producer and a regulator of gut health [48]. As an anaerobic, Gram-positive, butyric acid-producing bacillus, Clostridium butyricum resides in the gastrointestinal tract and has a protective role against pathogenic bacteria and intestinal injury by modulating gut microbial metabolites, such as short-chain fatty acids (SCFAs) [49]. Several studies have indicated that, via modulation of the gut microbiota, Clostridium butyricum effectively improved major depressive disorder [50], irritable bowel syndrome (IBS) [51], allergic rhinitis [52], and cerebral ischemia/reperfusion (I/R) injury [48]. They also suggested that Clostridium butyricum influenced microbial metabolism by optimizing the structure of microbiota and enhancing acetate production and utilization for butyrate production [53]. In our study, the oral administration of Clostridium butyricum during early life had an influence on infants’ gut microbiota’s compositions at 6 months of age. It significantly increased the Gram-negative, conditionally pathogenic Escherichia/Shigella at 6 months of age. Further clinical studies considering these factors should explore possible roles of Clostridium butyricum’s consumption in gut microbiol development. We should note that the overuse of probiotics in healthy infants is a serious problem that may pose a threat to human health. Studies in healthy adults have revealed that probiotic consumption cannot improve blood lipid profiles, and its benefits (e.g., stool consistency and bowel movement) are temporary [54]. Despite the current lack of warnings, there is insufficient evidence to conclude that probiotics have no deleterious side effects. The use of probiotics in healthy infants in order to provide sustained health benefits requires more in-depth study to confirm.
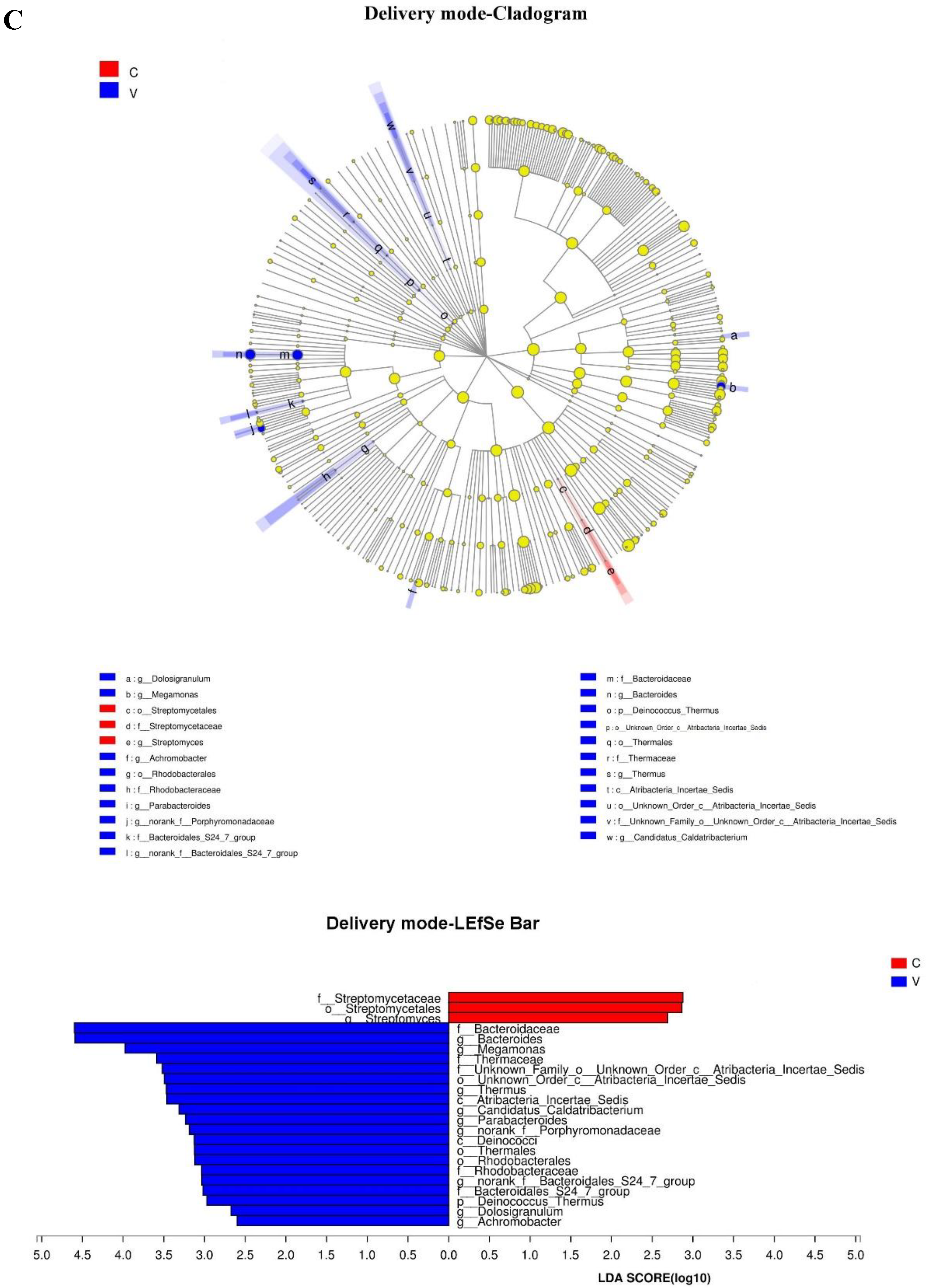
The delivery mode can affect infants’ gut microbiota [55,56,57]. Our study showed that differences caused by delivery mode were absent in the infants of six months. Chu et al. [58] proved that the gut microbiotas of neonates were influenced by mode of delivery at birth, but the influences disappeared at 6 weeks after birth. Compared with vaginally born infants, Penders et al. [59] found that those Cesarean-section born had lower abundance of Bifidobacteria and Bacteroides, whereas they had Clostridium difficile predominance. Another study showed that differences in infants’ early gut microbial colonization caused by delivery mode persisted 4 months after birth [55,60]. At 3 months after birth, the abundances of Actinomycobacteria and Bacteroides were relatively lower, and infants delivered by Cesarean section had a predominant Firmicutes phylum. For vaginally delivered infants, the dominant bacteria were Bifidobacteria and Bacteroides. Clostridium and Lactobacillus were the dominant bacteria for infants delivered by Cesarean section. However, these differences disappeared 6–12 months after delivery [61]. Given the complexities of the research results, our studies made an assumption: that diet consumption was an important factor to renovate gut microbiota in early human life [62]. The structures and compositions of infants’ gut microbiotas born with Cesarean section were similar to those of infants delivered vaginally at 4 months [60]. With the transmission of solid food, it is meaningful to investigate how dietary patterns influence an infant’s gut microbiota.
Recently, the public has been paying more attention to the abuse and overuse of antibiotics. In a 3-year longitudinal study of 39 infants in England, Yassour et al. found that there was a lower diversity of gut microbiota in antibiotic treated children than untreated children [63]. After antibiotic exposure one month later, the gut microbiota appeared to return to normal, which was consistent with our study. However, Bokulich et al. analyzed 43 infants over the first 2 years of life in the United States [57]. Infants who had prenatal and postnatal antibiotic treatment during the first 24 months of life showed a delayed development of the gut microbiome and suppressed Clostridiales, including Lachnospiraceae. However, considering the different durations, doses, ages, and approaches to antibiotics including the deviations between the methods of administration, it is not surprising that the results were incomparable among themselves. Numerous studies have shown that antibiotic exposure contributed to poor infant health at a later stage of life, such as by increasing the risk for childhood obesity [20], asthma [64], food allergies [65], and inflammatory bowel disease [66]. Future studies will need to keep exploring the impact of antibiotic use on children’s health and development longitudinally.
5. Limitations
Although our study attempted to provide a comprehensive look into the relationships between maternal postpartum practices, including other relevant factors (e.g., antibiotic exposure, probiotics consumption, delivery mode, and feeding type), and infant gut microbiota at 6 months of age, there are several limitations to be addressed in future studies. First, it would have been more significant if we had explored the effect of antibiotics on infants’ gut microbiotas in a quantificational measurement. Second, the study design of 50 cross-sectional samples at 6 months of age, rather than at earlier age, may have missed capturing effects of early maternal practices and factors (i.e., “doing the month,” mode of delivery, feeding type, antibiotic exposure, and probiotic consumption) on infants’ gut microbiotas. Third, it is important to collect longitudinal samples to understand the longitudinal dynamics of the gut microbiotas in infants and their influencing factors. Finally, the data on relative abundance were not sensitive enough to distinguish difference among groups compared to absolute quantification.
6. Conclusions
The results suggested that maternal adherence to traditionally practical behaviors of “doing the month” can induce changes in the gut microbiotas of infants. It emphasized the importance of mothers’ behaviors in seeding infants’ gut microbiotas. This study showed that Clostridium butyricum use early in life could increase conditional pathogenic bacteria Escherichia/Shigella. Probiotic use has raised enormous safety concerns. A longitudinal study with a larger sample size should be conducted.
Supplementary Materials
Supplementary materials can be found at https://0-www-mdpi-com.brum.beds.ac.uk/2076-2607/7/11/511/s1.
Author Contributions
Project administration and supervision, X.C.; writing—reviewing and editing, J.B.; funding acquisition and writing—reviewing and editing, Y.L.; writing—original draft preparation, Y.W.
Funding
This research was funded by Natural Science Foundation of Hubei Province, grant number 2017CFB343, and The Key Independent Scientific Research Project of WuHan University School of Health Sciences, grant number JSZD2016001.
Acknowledgments
We thank Shanghai Meiji biomedical technology co. LTD for help in sample management and sequence production; Y.W. for conducting the research and analyzing data; Y.W., Y.L., and J.B. for writing and revising paper; and X.C. for development of overall research plan, and helpful discussions. Y.W. and other co-authors have no conflict of interest.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.J.N.B. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.H.; Huang, K.C.; Huang, T.J.; Hsu, R.W.W. Case Reports: Fatal Necrotizing Fasciitis Caused by Aeromonas sobria in Two Diabetic Patients. Clin. Orthop. Relat. Res. 2009, 467, 846–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.J.; Ma, L.J.; Ma, Y.B.; Zhang, F.M.; Zhao, C.H.; Nie, Y.Z. Insights into the role of gut microbiota in obesity: Pathogenesis, mechanisms, and therapeutic perspectives. Protein Cell 2018, 9, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Hand, T.W.; Vujkovic-Cvijin, I.; Ridaura, V.K.; Belkaid, Y. Linking the Microbiota, Chronic Disease, and the Immune System. Trends Endocrinol. Metab. 2016, 27, 831–843. [Google Scholar] [CrossRef]
- Sprockett, D.; Fukami, T.; Relman, D.A. Role of priority effects in the early-life assembly of the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 197–205. [Google Scholar] [CrossRef]
- Walker, R.W.; Clemente, J.C.; Peter, I.; Loos, R.J.F. The prenatal gut microbiome: Are we colonized with bacteria in utero? Pediatric Obes. 2017, 12, 3–17. [Google Scholar] [CrossRef]
- Chang, J.Y.; Shin, S.M.; Chun, J.; Lee, J.H.; Seo, J.K. Pyrosequencing-based Molecular Monitoring of the Intestinal Bacterial Colonization in Preterm Infants. J. Pediatric Gastroenterol. Nutr. 2011, 53, 512–519. [Google Scholar] [CrossRef]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef]
- Gosalbes, M.J.; Llop, S.; Valles, Y.; Moya, A.; Ballester, F.; Francino, M.P. Meconium microbiota types dominated by lactic acid or enteric bacteria are differentially associated with maternal eczema and respiratory problems in infants. Clin. Exp. Allergy 2013, 43, 198–211. [Google Scholar] [CrossRef]
- Rautava, S.; Collado, M.C.; Salminen, S.; Isolauri, E. Probiotics Modulate Host-Microbe Interaction in the Placenta and Fetal Gut: A Randomized, Double-Blind, Placebo-Controlled Trial. Neonatology 2012, 102, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.M. Author response to comment on “The placenta harbors a unique microbiome”. Sci. Transl. Med. 2014, 6, 245. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Xiao, X.H.; Zhang, Q.; Mao, L.L.; Yu, M.; Xu, J.P. The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates. Nutrients 2015, 7, 6924–6937. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; Fernandez, L.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodriguez, J.M. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef]
- Ferretti, P.; Pasolli, E.; Tett, A.; Asnicar, F.; Gorfer, V.; Fedi, S.; Armanini, F.; Truong, D.T.; Manara, S.; Zolfo, M.; et al. Mother-to-Infant Microbial Transmission from Different Body Sites Shapes the Developing Infant Gut Microbiome. Cell Host Microbe 2018, 24, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Gohir, W.; Ratcliffe, E.M.; Sloboda, D.M. Of the bugs that shape us: Maternal obesity, the gut microbiome, and long-term disease risk. Pediatric Res. 2015, 77, 196–204. [Google Scholar] [CrossRef]
- Stanislawski, M.A.; Dabelea, D.; Wagner, B.D.; Sontag, M.K.; Lozupone, C.A.; Eggesbo, M. Pre-pregnancy weight, gestational weight gain, and the gut microbiota of mothers and their infants. Microbiome 2017, 5, 113. [Google Scholar] [CrossRef]
- Jasarevic, E.; Howerton, C.L.; Howard, C.D.; Bale, T.L. Alterations in the Vaginal Microbiome by Maternal Stress Are Associated with Metabolic Reprogramming of the Offspring Gut and Brain. Endocrinology 2015, 156, 3265–3276. [Google Scholar] [CrossRef]
- Bailey, M.T.; Lubach, G.R.; Coe, C.L. Prenatal stress alters bacterial colonization of the gut in infant monkeys. J. Pediatric Gastroenterol. Nutr. 2004, 38, 414–421. [Google Scholar] [CrossRef]
- Mueller, N.T.; Whyatt, R.; Hoepner, L.; Oberfield, S.; Dominguez-Bello, M.G.; Widen, E.M.; Hassoun, A.; Perera, F.; Rundle, A. Prenatal exposure to antibiotics, cesarean section and risk of childhood obesity. Int. J. Obes. 2015, 39, 665–670. [Google Scholar] [CrossRef]
- von Mutius, E. The microbial environment and its influence on asthma prevention in early life. J. Allergy Clin. Immunol. 2016, 137, 680–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressa, C.; Bailen-Andrino, M.; Perez-Santiago, J.; Gonzalez-Soltero, R.; Perez, M.; Montalvo-Lominchar, M.G.; Mate-Munoz, J.L.; Dominguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, M.G.V.; Closs, V.E.; Junges, V.M.; Schwanke, C.H.A. Impact of human aging and modern lifestyle on gut microbiota. Crit. Rev. Food Sci. 2018, 58, 1557–1564. [Google Scholar] [CrossRef] [PubMed]
- Callister, L.C. Doing the month: Chinese postpartum practices. MCN Am. J. Matern. Child Nurs. 2006, 31, 390. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Petrini, M.; Maloni, J.A. “Doing the month”: Postpartum practices in Chinese women. Nurs. Health Sci. 2015, 17, 5–14. [Google Scholar] [CrossRef]
- Bravi, F.; Wiens, F.; Decarli, A.; Dal Pont, A.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.; Yang, S.X.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association Between Breast Milk Bacterial Communities and Establishment and Development of the Infant Gut Microbiome. JAMA Pediatric 2017, 171, 647–654. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Maloni, J.A.; Petrini, M.A. Effect of Postpartum Practices of Doing the Month on Chinese Women’s Physical and Psychological Health. Biol. Res. Nurs. 2014, 16, 55–63. [Google Scholar] [CrossRef]
- Zijlmans, M.A.C.; Korpela, K.; Riksen-Walraven, J.M.; de Vos, W.M.; de Weerth, C. Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrino 2015, 53, 233–245. [Google Scholar] [CrossRef]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.Y.; Qin, Y.W.; Tang, L.Q.; Zhao, H.; Stenvang, J.; Li, Y.L.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2017, 66, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Chien, L.Y.; Tai, C.J.; Ko, Y.L.; Huang, C.H.; Sheu, S.J. Adherence to “doing-the-month” practices is associated with fewer physical and depressive symptoms among postpartum women in Taiwan. Res. Nurs. Health 2006, 29, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Wen-Harn, P.; Wen-Ting, Y.J.A. How to define obesity? Evidence-based multiple action points for public awareness, screening, and treatment: An extension of Asian-Pacific recommendations. Asia Pac. J. Clin. Nutr. 2008, 17, 5. [Google Scholar]
- Rasmussen, K.M.; Yaktine, A.L. Weight Gain During Pregnancy: Reexamining the Guidelines; Rasmussen, K.M., Yaktine, A.L., Eds.; National Academies Press: Washington, DC, USA, 2009. [Google Scholar] [CrossRef]
- Yassour, M.; Jason, E.; Hogstrom, L.J.; Arthur, T.D.; Tripathi, S.; Siljander, H.; Selvenius, J.; Oikarinen, S.; Hyoty, H.; Virtanen, S.M.; et al. Strain-Level Analysis of Mother-to-Child Bacterial Transmission during the First Few Months of Life. Cell Host Microbe 2018, 24, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Sagheddu, V.; Patrone, V.; Miragoli, F.; Puglisi, E.; Morelli, L. Infant Early Gut Colonization by Lachnospiraceae: High Frequency of Ruminococcus gnavus. Front. Pediatrics 2016, 4, 57. [Google Scholar] [CrossRef] [Green Version]
- Petrof, E.O.; Gloor, G.B.; Vanner, S.J.; Weese, S.J.; Carter, D.; Daigneault, M.C.; Brown, E.M.; Schroeter, K.; Allen-Vercoe, E. Stool substitute transplant therapy for the eradication of Clostridium difficile infection: ‘RePOOPulating’ the gut. Microbiome 2013, 1, 3. [Google Scholar] [CrossRef]
- Cho, I.; Yamanishi, S.; Cox, L.; Methe, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef]
- Aiello, A.E.; Larson, E.L.; Sedlak, R. Hidden heroes of the health revolution Sanitation and personal hygiene. Am. J. Infect. Control 2008, 36, 128–151. [Google Scholar] [CrossRef]
- Maloni, J.A. Antepartum Bed Rest for Pregnancy Complications: Efficacy and Safety for Preventing Preterm Birth. Biol. Res. Nurs. 2010, 12, 106–124. [Google Scholar] [CrossRef]
- Shou-Hua, L.U.J.; Medicine, E. Investigation of postpartum depression and its related factors in community. J. Clin. Exp. Med. 2011, 10, 1099–1100. [Google Scholar]
- Gao, L.L.; Chan, S.W.C.; You, L.M.; Li, X.M. Experiences of postpartum depression among first-time mothers in mainland China. J. Adv. Nurs. 2010, 66, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Bezirtzoglou, E.; Tsiotsias, A.; Welling, G.W. Microbiota profile in feces of breast- and formula-fed newborns by using fluorescence in situ hybridization (FISH). Anaerobe 2011, 17, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Gritz, E.C.; Bhandari, V. The human neonatal gut microbiome: A brief review. Front. Pediatrics 2015, 3, 17. [Google Scholar] [CrossRef]
- Harmsen, H.J.M.; Wildeboerveloo, A.C.M.; Raangs, G.C.; Wagendorp, A.A.; Klijn, N.; Bindels, J.G.; Welling, G.W.J. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J. Pediatric Gastroenterol. Nutr. 2000, 30, 61–67. [Google Scholar] [CrossRef]
- Cong, X.; Judge, M.; Xu, W.; Diallo, A.; Janton, S.; Brownell, E.A.; Maas, K.; Graf, J.J.N.R. Influence of Feeding Type on Gut Microbiome Development in Hospitalized Preterm Infants. Nurs. Res. 2017, 66, 123. [Google Scholar] [CrossRef]
- Sun, J.; Wang, F.Y.; Ling, Z.X.; Yu, X.C.; Chen, W.Q.; Li, H.X.; Jin, J.T.; Pang, M.Q.; Zhang, H.Q.; Yu, J.J.; et al. Clostridium butyricum attenuates cerebral ischemia/reperfusion injury in diabetic mice via modulation of gut microbiota. Brain Res. 2016, 1642, 180–188. [Google Scholar] [CrossRef]
- Falony, G.; Vieira-Silva, S.; Raes, J. Microbiology Meets Big Data: The Case of Gut Microbiota-Derived Trimethylamine. Annu. Rev. Microbiol. 2015, 69, 305–321. [Google Scholar] [CrossRef]
- Miyaoka, T.; Kanayama, M.; Wake, R.; Hashioka, S.; Hayashida, M.; Nagahama, M.; Okazaki, S.; Yamashita, S.; Miura, S.; Miki, H.; et al. Clostridium butyricum MIYAIRI 588 as Adjunctive Therapy for Treatment-Resistant Major Depressive Disorder: A Prospective Open-Label Trial. Clin. Neuropharmacol. 2018, 41, 151–155. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Li, M.; Li, Y.Y.; Li, L.X.; Zhai, W.Z.; Wang, P.; Yang, X.X.; Gu, X.; Song, L.J.; Li, Z.; et al. The effect of Clostridium butyricum on symptoms and fecal microbiota in diarrhea-dominant irritable bowel syndrome: A randomized, double-blind, placebo-controlled trial. Sci. Rep. 2018, 8, 2964. [Google Scholar] [CrossRef]
- Xu, L.Z.; Yang, L.T.; Qiu, S.Q.; Yang, G.; Luo, X.Q.; Miao, B.P.; Geng, X.R.; Liu, Z.Q.; Liu, J.; Wen, Z.; et al. Combination of specific allergen and probiotics induces specific regulatory B cells and enhances specific immunotherapy effect on allergic rhinitis. Oncotarget 2016, 7, 54360–54369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, X.Y.; Liu, P.; Zhao, J.B.; Sun, J.; Guan, W.Y.; Johnston, L.J.; Leyesque, C.L.; Fan, P.X.; He, T.; et al. Dietary Clostridium butyricum Induces a Phased Shift in Fecal Microbiota Structure and Increases the Acetic Acid-Producing Bacteria in a Weaned Piglet Model. J. Agric. Food Chem. 2018, 66, 5157–5166. [Google Scholar] [CrossRef] [PubMed]
- Khalesi, S.; Bellissimo, N.; Vandelanotte, C.; Williams, S.; Stanley, D.; Irwin, C. A review of probiotic supplementation in healthy adults: Helpful or hype? Eur. J. Clin. Nutr. 2018, 73, 24. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [Green Version]
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hamalainen, A.M.; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 1551. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.L.; Lieber, A.D.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343. [Google Scholar] [CrossRef]
- Chu, D.M.; Ma, J.; Prince, A.L.; Antony, K.M.; Seferovic, M.D.; Aagaard, K.M. Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery. Nat. Med. 2017, 23, 314. [Google Scholar] [CrossRef]
- Penders, J.; Thijs, C.; Vink, C.; Stelma, F.F.; Snijders, B.; Kummeling, I.; Brandt, P.A.V.D.; Stobberingh, E.E.J.P. Factors Influencing the Composition of the Intestinal Microbiota in Early Infancy. Pediatrics 2006, 118, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.B.; Konya, T.; Maughan, H.; Guttman, D.S.; Field, C.J.; Chari, R.S.; Sears, M.R.; Becker, A.B.; Scott, J.A.; Kozyrskyj, A.L.; et al. Gut microbiota of healthy Canadian infants: Profiles by mode of delivery and infant diet at 4 months. Can. Med. Assoc. J. 2013, 185, 385–394. [Google Scholar] [CrossRef]
- Rutayisire, E.; Huang, K.; Liu, Y.H.; Tao, F.B. The mode of delivery affects the diversity and colonization pattern of the gut microbiota during the first year of infants’ life: A systematic review. BMC Gastroenterol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Baumann-Dudenhoeffer, A.M.; D’Souza, A.W.; Tarr, P.I.; Warner, B.B.; Dantas, G. Infant diet and maternal gestational weight gain predict early metabolic maturation of gut microbiomes. Nat. Med. 2018, 24, 1822. [Google Scholar] [CrossRef] [PubMed]
- Yassour, M.; Vatanen, T.; Siljander, H.; Hämäläinen, A.M.; Härkönen, T.; Ryhänen, S.J. Natural history of the infant gut microbiome and impact of antibiotic treatment on bacterial strain diversity and stability. Sci. Transl. Med. 2016, 8, 343–381. [Google Scholar] [CrossRef] [PubMed]
- Lapin, B.; Piorkowski, J.; Ownby, D.; Freels, S.; Chavez, N.; Hernandez, E.; Wagner-Cassanova, C.; Pelzel, D.; Vergara, C.; Persky, V. Relationship between prenatal antibiotic use and asthma in at-risk children. Ann. Allergy Asthma Immunol. 2015, 114, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, A.G.; Pollak, J.; Glass, T.A.; Poulsen, M.N.; Bailey-Davis, L.; Mowery, J.; Schwartz, B.S. Early-life antibiotic use and subsequent diagnosis of food allergy and allergic diseases. Clin. Exp. Allergy 2017, 47, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Hviid, A.; Svanström, H.; Frisch, M. Antibiotic use and inflammatory bowel diseases in childhood. Gut 2011, 60, 49–54. [Google Scholar] [CrossRef]
Figure 1.
Relative abundance (≥1%) of infant gut microbiota at the genus level in the differentiation of the microbiota of six-month old infants. ** Each bar represents one sample.
Figure 1.
Relative abundance (≥1%) of infant gut microbiota at the genus level in the differentiation of the microbiota of six-month old infants. ** Each bar represents one sample.

Figure 2.
Principal coordinate analysis (PCoA) of infant gut microbiota using Bray–Curtis distance matrix. Two-dimensional PCoA was used to describe the relative abundance of infant gut microbiota. Each point represents a single sample and is colored by different groups (A) antibiotic exposure; (B) probiotic consumption; (C) feeding type; (D) delivery mode.
Figure 2.
Principal coordinate analysis (PCoA) of infant gut microbiota using Bray–Curtis distance matrix. Two-dimensional PCoA was used to describe the relative abundance of infant gut microbiota. Each point represents a single sample and is colored by different groups (A) antibiotic exposure; (B) probiotic consumption; (C) feeding type; (D) delivery mode.


Figure 3.
Taxonomic representation of statistically and biologically consistent differences between different groups ((A) probiotic consumption; (B) antibiotic exposure; (C) delivery mode; (D) feeding type). Differences are represented by the color of the most abundant genus. The diameter of each circle is proportional to taxon abundance; from inside to outside circle indicate the phylum to genus level in turns (g: genus; f: family; o: order; c: class). Histogram of the linear discriminant analysis (LDA) scores for differentially abundant genera. The threshold on the logarithmic LDA score for discriminative features was set to 2.0; the larger the LDA score, the greater influence of species abundance on the difference between two groups. ((A) Probiotic consumption: More abundant in no consumption group: Aeromonadaceae, Aeromonas, Aeromonadales, Tyzzerella, Staphylococcaceae, Bacillales, Staphylococcus, Peptostreptococcaceae, Carnobacteriaceae, and Granulicatella. More abundance in consumption group: Escherichia_Shigella, Lachnoclostridium, and Lachnospiraceae. (B) Antibiotic exposure: More abundance in no antibiotic exposure group: Lachnoclostridium and Bacillales, Anaerotruncus. (C) Delivery mode: More abundance in Cesarean section group: Streptomycetaceae, Streptomycetales and Streptomyces. More abundance in vaginal delivery group: Bacteroidaceae, Bacteroides, Megamonas, Thermaceae, (f)Atribacteria Incertae Sedis, (o)Atribacteria Incertae Sedis Thermus, (c)Atribacteria Incertae Sedis, Candidatus Caldatribacterium, Parabacteroides, Porphyromonadaceae, Deinococci, Thermales, Rhodobacterales, Rhodobacteraceae, (g)Bacteroidales S247 group, (f)Bacteroidales S247 group, Deinococcus Thermus, Dolosigranulum, and Achromobacter. (D) Feeding type: More abundance in breast-fed group: Burkholderiales and Epulopiscium. More abundance in mixed-fed group: Dysgonomonas, Solobacterium, and Tissierella).
Figure 3.
Taxonomic representation of statistically and biologically consistent differences between different groups ((A) probiotic consumption; (B) antibiotic exposure; (C) delivery mode; (D) feeding type). Differences are represented by the color of the most abundant genus. The diameter of each circle is proportional to taxon abundance; from inside to outside circle indicate the phylum to genus level in turns (g: genus; f: family; o: order; c: class). Histogram of the linear discriminant analysis (LDA) scores for differentially abundant genera. The threshold on the logarithmic LDA score for discriminative features was set to 2.0; the larger the LDA score, the greater influence of species abundance on the difference between two groups. ((A) Probiotic consumption: More abundant in no consumption group: Aeromonadaceae, Aeromonas, Aeromonadales, Tyzzerella, Staphylococcaceae, Bacillales, Staphylococcus, Peptostreptococcaceae, Carnobacteriaceae, and Granulicatella. More abundance in consumption group: Escherichia_Shigella, Lachnoclostridium, and Lachnospiraceae. (B) Antibiotic exposure: More abundance in no antibiotic exposure group: Lachnoclostridium and Bacillales, Anaerotruncus. (C) Delivery mode: More abundance in Cesarean section group: Streptomycetaceae, Streptomycetales and Streptomyces. More abundance in vaginal delivery group: Bacteroidaceae, Bacteroides, Megamonas, Thermaceae, (f)Atribacteria Incertae Sedis, (o)Atribacteria Incertae Sedis Thermus, (c)Atribacteria Incertae Sedis, Candidatus Caldatribacterium, Parabacteroides, Porphyromonadaceae, Deinococci, Thermales, Rhodobacterales, Rhodobacteraceae, (g)Bacteroidales S247 group, (f)Bacteroidales S247 group, Deinococcus Thermus, Dolosigranulum, and Achromobacter. (D) Feeding type: More abundance in breast-fed group: Burkholderiales and Epulopiscium. More abundance in mixed-fed group: Dysgonomonas, Solobacterium, and Tissierella).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Demographic and birth characteristics of infants (n = 50).
| Items | Total N (%)/Mean (SD) |
|---|---|
| Maternal age | 30.13 (27.01−33.25) |
| Gestation | |
| 1 | 31 (62%) |
| 2 | 12 (24%) |
| 3 | 6 (12%) |
| 4 | 1 (2%) |
| Parity | |
| 0 | 34 (68%) |
| 1 prior child | 16 (32%) |
| Maternal education | |
| <12 a | 2 (4%) |
| 12 a | 31 (62%) |
| >12 a | 17 (34%) |
| Pre-pregnancy BMI category | |
| Underweight | 9 (18%) |
| Normal | 35 (70%) |
| Overweight | 6 (12%) |
| GWG category | |
| less | 8 (16%) |
| adequate | 20 (40%) |
| excessive | 22 (44%) |
| Gestational week | 39.69 (38.73–40.65) |
| Mode of delivery | |
| Vaginal | 22 (44%) |
| Cesarean section | 28 (56%) |
| Adherence to “doing the month” | 81.94 (66.34–97.54) |
| Feeding type | |
| Breast | 30 (60%) |
| Mixed | 20 (40%) |
| Probiotic consumption | |
| Yes | 35 (70%) |
| No | 15 (30%) |
| Antibiotic exposure | |
| Yes | 17 (34%) |
| No | 33 (66%) |
| Birth weight | 3.45 (3.09–3.81) |
| Sex | |
| Male | 20 (40%) |
| Female | 30 (60%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Liu, Y.; Bai, J.; Chen, X. The Effect of Maternal Postpartum Practices on Infant Gut Microbiota: A Chinese Cohort Study. Microorganisms 2019, 7, 511. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110511
AMA Style
Wang Y, Liu Y, Bai J, Chen X. The Effect of Maternal Postpartum Practices on Infant Gut Microbiota: A Chinese Cohort Study. Microorganisms. 2019; 7(11):511. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110511
Chicago/Turabian StyleWang, Ying, Yanqun Liu, Jinbing Bai, and Xiaoli Chen. 2019. "The Effect of Maternal Postpartum Practices on Infant Gut Microbiota: A Chinese Cohort Study" Microorganisms 7, no. 11: 511. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110511
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.