The Dark Side of Microalgae Biotechnology: A Heterotrophic Biorefinery Platform Directed to ω-3 Rich Lipid Production


Abstract
:1. Introduction
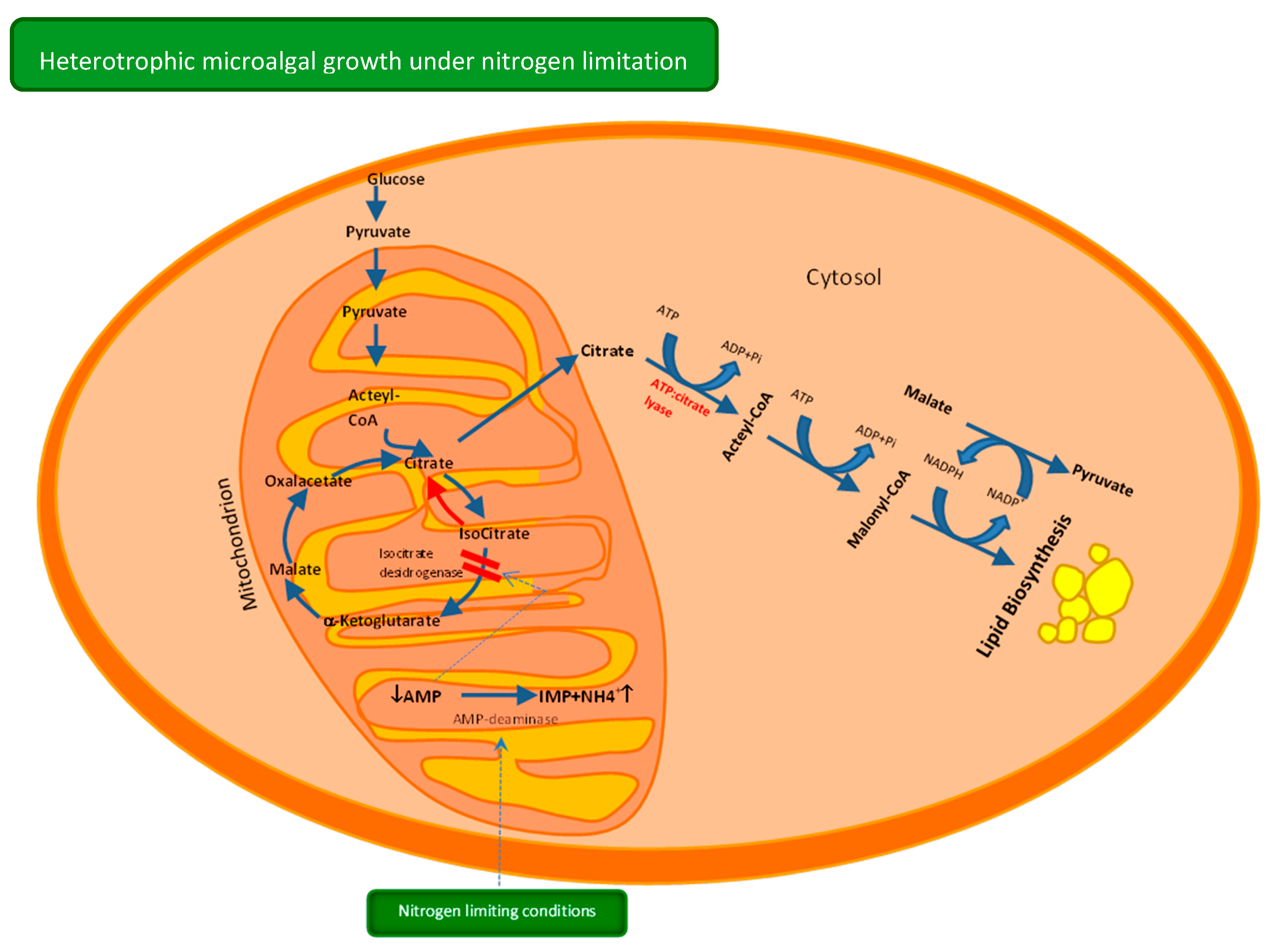
2. Heterotrophic Metabolism—Carbon Uptake and Lipid Synthesis
3. Oleaginous Heterotrophic Microalgae Strains for ω-3 Compounds
4. Effect of Operational Conditions on Microalgal Growth, Lipids and DHA Production
4.1. Medium Composition
4.2. Culture Mode
4.3. Dissolved Oxygen
4.4. Culture Medium pH
4.5. Temperature
4.6. Salinity
4.7. Microalgal Culture Monitoring by Flow Cytometry (FC)
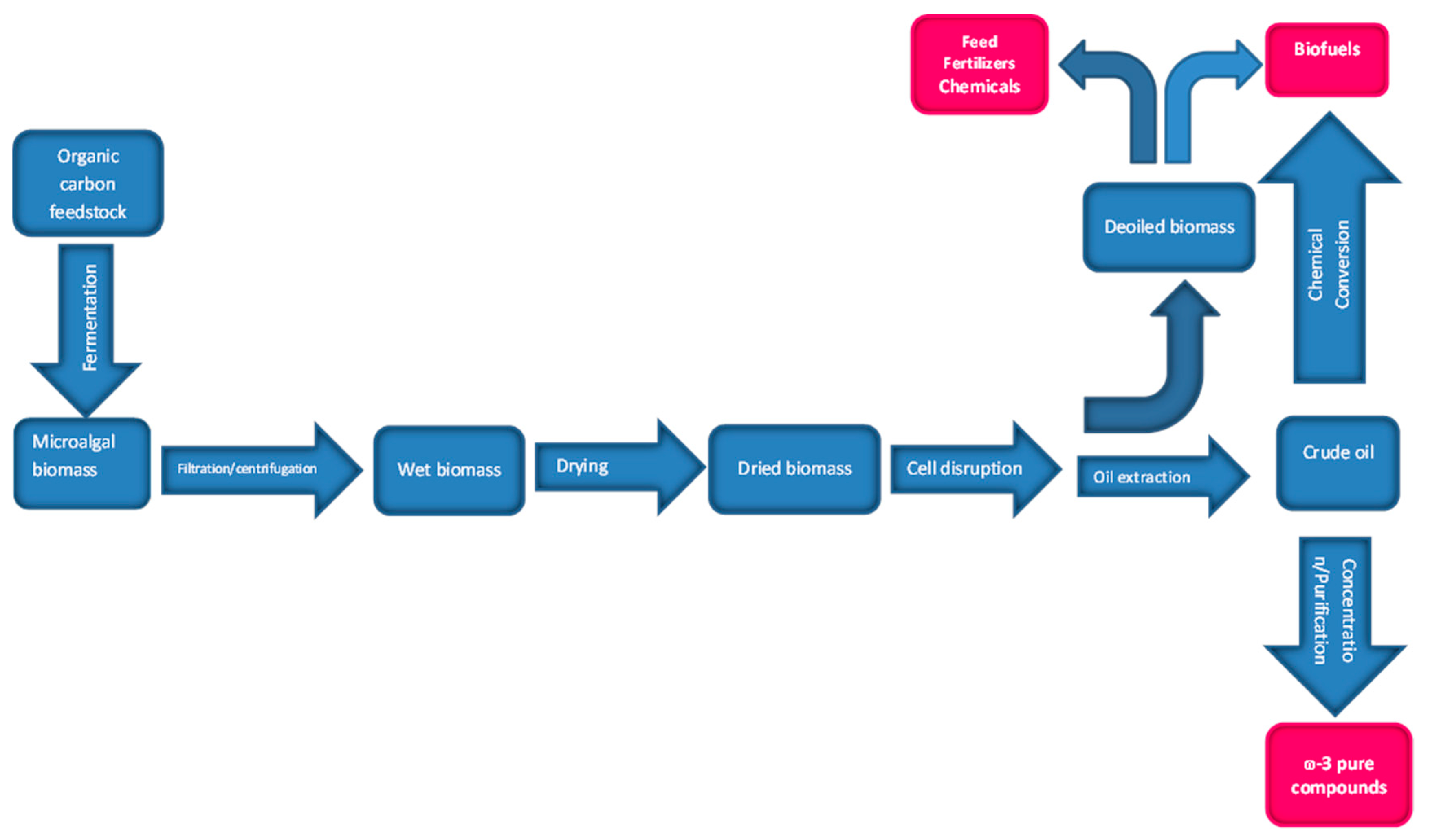
5. Downstream Processing
5.1. Oil Extraction
5.2. ω-3 Compounds Purification
6. EPA/DHA Industrial Production and Applications
7. EPA/DHA Industrial Producers
8. The Heterotrophic Biorefinery Platform
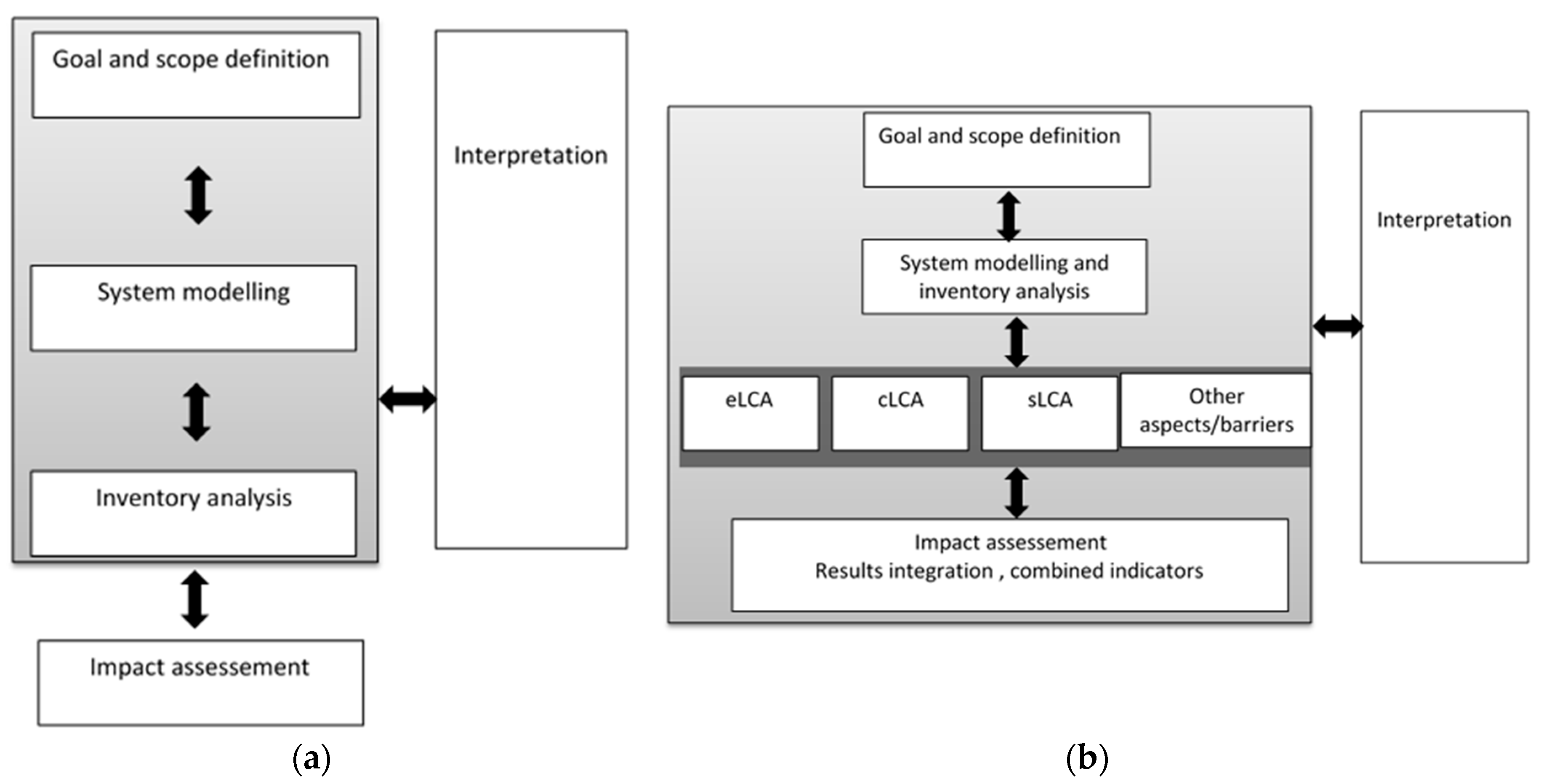
9. Sustainability Assessment of Benchmark—Omega 3 (ω-3) Biorefinery
10. Bottlenecks, Challenges and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Braunwald, T.; French, W.T.; Claupein, W.; Graeff-Honninger, S. Economic assessment of microbial biodiesel production using heterotrophic yeasts. Int. J. Green Energy 2016, 13, 274–282. [Google Scholar] [CrossRef]
- Da Silva, T.L.; Reis, A. Scale-up Problems for the Large Scale Production of Algae; Chapter 6; Capital Publishing Company: Sterling, VA, USA, 2015. [Google Scholar] [CrossRef]
- Patel, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. Heterotrophic cultivation of Auxenochlorella protothecoides using forest biomass as a feedstock for sustainable biodiesel production. Biotechnol. Biofuels 2018, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Finco, A.M.O.; Mamani, L.D.G.; Carvalho, J.C.; de Melo Pereira, G.V.; Thomaz-Soccol, V.; Soccol, C.R. Technological trends and market perspectives for production of microbial oils rich in omega-3. Crit. Rev. Biotechnol. 2017, 37, 656–671. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Eggersdorfer, M. Is the world supply of omega-3 fatty acids adequate for optimal human nutrition? Curr. Opin. Clin. Nutr. 2015, 18, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Voort, M.; Spruijtm, J.; Potters, J.; de Wolf, P.; Elissen, H. Socio-Economic Assessment of Algae-Based PUFA Production; The Value Chain from Microalgae to PUFA (‘PUFAChain’) Project no. 613303; PUFAChain: Göttingen, Germany, 2017. [Google Scholar]
- Medina, I.; Aubourg, S.P.; Perez Martin, R. Composition of phospholipids of white muscle of six tuna species. Lipids 1995, 30, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Slade, R.; Bauen, A. Micro-algae cultivation for biofuels: Cost, energy balance, environmental impacts and future prospects. Biomass Bioenergy 2013, 53, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef]
- Bruton, T.; Lyons, H.; Lerat, Y.; Stanley, M.; Rasmussen, M.B. A Review of the Potential of Marine Algae as a Source of Biofuel in Ireland; Sustainable Energy Ireland: Dublin, Ireland, 2009. [Google Scholar]
- Kim, S.; Park, J.E.; Cho, Y.B.; Hwang, S.J. Growth rate, organic carbon and nutrient removal rates of Chlorella sorokiniana in autotrophic, heterotrophic and mixotrophic conditions. Bioresour. Technol. 2013, 144, 8–13. [Google Scholar] [CrossRef]
- Morales-Sanchez, D.; Martinez-Rodriguez, O.A.; Martinez, A. Heterotrophic cultivation of microalgae: Production of metabolites of commercial interest. J. Chem. Technol. Biotechnol. 2017, 92, 925–936. [Google Scholar] [CrossRef]
- Khan, M.; Karmakar, R.; Das, B.; Diba, F.; Razu, M. Heterotrophic Growth of Micro Algae. In Recent Advances in Microalgal Biotechnology; Liu, J., Sun, Z., Eds.; OMICS: Hyderabad, India, 2016; pp. 1–19. [Google Scholar]
- Hu, J.J.; Nagarajan, D.; Zhang, Q.G.; Chang, J.S.; Lee, D.J. Heterotrophic cultivation of microalgae for pigment production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef]
- Ochsenreither, K.; Gluck, C.; Stressler, T.; Fischer, L.; Syldatk, C. Production Strategies and Applications of Microbial Single Cell Oils. Front. Microbiol. 2016, 7, 1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Swaaf, M.E.; Rijk, T.C.; Eggink, G.; Sijtsma, L. Optimization of docosahexaenoic acid production in batch cultivations by Crypthecodinium cohnii. J. Biotechnol. 1999, 70, 185–192. [Google Scholar] [CrossRef]
- De Swaaf, M.E.; Pronk, J.T.; Sijtsma, L. Fed-batch cultivation of the docosahexaenoic-acid-producing marine alga Crypthecodinium cohnii on ethanol. Appl. Microbiol. Biotechnol. 2003, 61, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Hosoglu, M.I. Aroma characterization of five microalgae species using solid-phase microextraction and gas chromatography-mass spectrometry/olfactometry. Food Chem. 2018, 240, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Pleissner, D.; Lam, W.C.; Sun, Z.; Lin, C.S.K. Food waste as nutrient source in heterotrophic microalgae cultivation. Bioresour. Technol. 2013, 137, 139–146. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Zhang, X.C.; Ji, L.; Song, X.J.; Kuang, C.H. Changes of lipid content and fatty acid composition of Schizochytrium limacinum in response to different temperatures and salinities. Process Biochem. 2007, 42, 210–214. [Google Scholar] [CrossRef]
- Mendes, A.; da Silva, T.L.; Reis, A. DHA concentration and purification from the marine heterotrophic microalga Crypthecodinium cohnii CCMP 316 by winterization and urea complexation. Food Technol. Biotechnol. 2007, 45, 38–44. [Google Scholar]
- Gong, Y.; Liu, J.; Jiang, M.; Liang, Z.; Jin, H.; Hu, X.; Wan, X.; Hu, C. Improvement of Omega-3 Docosahexaenoic Acid Production by Marine Dinoflagellate Crypthecodinium cohnii Using Rapeseed Meal Hydrolysate and Waste Molasses as Feedstock. PLoS ONE 2015, 10, 125368. [Google Scholar] [CrossRef] [Green Version]
- De Swaaf, M.E.; de Rijk, T.C.; van der Meer, P.; Eggink, G.; Sijtsma, L. Analysis of docosahexaenoic acid biosynthesis in Crypthecodinium cohnii by 13C labelling and desaturase inhibitor experiments. J. Biotechnol. 2003, 103, 21–29. [Google Scholar] [CrossRef]
- De Swaaf, M.E.; Sijtsma, L.; Pronk, J.T. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing marine alga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef]
- Wynn, J.; Behrens, P.; Sundararajan, A.; Hansen, J.; Apt, K. Production of Single Cell Oils by Dinoflagellates. In Single Cell Oils, 2nd ed.; Academic Press: Cambridge, MA, USA, 2010; pp. 115–129. [Google Scholar] [CrossRef]
- Duan, X.; Ren, G.Y.; Liu, L.; Zhu, W.X. Salt-induced osmotic stress for lipid overproduction in batch culture of Chlorella vulgaris. Afr. J. Biotechnol. 2012, 11, 7072–7078. [Google Scholar]
- Sun, X.M.; Ren, L.J.; Bi, Z.Q.; Ji, X.J.; Zhao, Q.Y.; Jiang, L.; Huang, H. Development of a cooperative two-factor adaptive-evolution method to enhance lipid production and prevent lipid peroxidation in Schizochytrium sp. Biotechnol. Biofuels. 2018, 11, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillig, F.; Pilarek, M.; Junne, S.; Neubauer, P. Cultivation of marine microorganisms in single-use systems. Adv. Biochem. Eng. Biotechnol. 2014, 138, 179–206. [Google Scholar] [PubMed]
- Wang, C.C.; Lan, C.Q. Effects of shear stress on microalgae—A review. Biotechnol. Adv. 2018, 36, 986–1002. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.K.; Lam, C.M.; Ma, Z.Y.; Wong, Y.H.; Wong, J.T. Involvement of calcium mobilization from caffeine-sensitive stores in mechanically induced cell cycle arrest in the dinoflagellate Crypthecodinium cohnii. Cell Calcium 2006, 39, 259–274. [Google Scholar] [CrossRef]
- Hu, W.; Gladue, R.; Hansen, J.; Wojnar, C.; Chalmers, J.J. The sensitivity of the dinoflagellate Crypthecodinium cohnii to transient hydrodynamic forces and cell-bubble interactions. Biotechnol. Prog. 2007, 23, 1355–1362. [Google Scholar] [CrossRef]
- Safdar, W.; Zan, X.; Shamoon, M.; Sharif, H.R.; Mukama, O.; Tang, X.; Song, Y. Effects of twenty standard amino acids on biochemical constituents, docosahexaenoic acid production and metabolic activity changes of Crypthecodinium cohnii. Bioresour. Technol. 2017, 238, 738–743. [Google Scholar] [CrossRef]
- Guo, X.-L.; Ji, X.-J.; Ren, L.-J.; Li, G.-L.; Huang, H. Improving Docosahexaenoic acid production by Schizchytrium sp. using a newly designed high-oxygen-supply bioreactor. AIChE 2015, 63, 4278–4286. [Google Scholar] [CrossRef]
- Safdar, W.; Shamoon, M.; Zan, X.; Haider, J.; Sharif, H.R.; Shoaib, M.; Song, Y. Growth kinetics, fatty acid composition and metabolic activity changes of Crypthecodinium cohnii under different nitrogen source and concentration. AMB Express 2017, 7, 85. [Google Scholar] [CrossRef]
- Wu, S.T.; Yu, S.T.; Lin, L.P. Effect of culture conditions on docosahexaenoic acid production by Schizochytrium sp S31. Process Biochem. 2005, 40, 3103–3108. [Google Scholar] [CrossRef]
- Ho, W.L.; Chou, C.C. Effects of carbon and nitrogen sources, sodium chloride and culture conditions on cytotoxin production by Salmonella choleraesuis. Int. J. Food Microbiol. 2001, 67, 81–88. [Google Scholar] [CrossRef]
- Behrens, P.W.; Kyle, D.J. Microalgae as a source of fatty acids. J. Food Lipids 1996, 3, 259–272. [Google Scholar] [CrossRef]
- Ward, O.P.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Process Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Bumbak, F.; Cook, S.; Zachleder, V.; Hauser, S.; Kovar, K. Best practices in heterotrophic high-cell-density microalgal processes: Achievements, potential and possible limitations. Appl. Microbiol. Biot. 2011, 91, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guneser, O.; Demirkol, A.; Yuceer, Y.K.; Togay, S.O.; Hosoglu, M.I.; Elibol, M. Production of flavor compounds from olive mill waste by Rhizopus oryzae and Candida tropicalis. Braz. J. Microbiol. 2017, 48, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarma, S.J.; Dhillon, G.S.; Brar, S.K.; Le Bihan, Y.; Buelna, G.; Verma, M. Investigation of the effect of different crude glycerol components on hydrogen production by Enterobacter aerogenes NRRL B-407. Renew. Energy 2013, 60, 566–571. [Google Scholar] [CrossRef]
- Da Silva, T.L.; Santos, C.A.; Reis, A. Multi-parameter flow cytometry as a tool to monitor heterotrophic microalgal batch fermentations for oil production towards biodiesel. Biotechnol. Bioprocess Eng. 2009, 14, 330–337. [Google Scholar] [CrossRef]
- De la Jara, A.; Mendoza, H.; Martel, A.; Molina, C.; Nordstron, L.; de la Rosa, V.; Diaz, R. Flow cytometric determination of lipid content in a marine dinoflagellate, Crypthecodinium cohnii. J. Appl. Phycol. 2003, 15, 433–438. [Google Scholar] [CrossRef]
- Da Silva, T.L.; Reis, A.; Medeiros, R.; Oliveira, A.C.; Gouveia, L. Oil Production Towards Biofuel from Autotrophic Microalgae Semicontinuous Cultivations Monitorized by Flow Cytometry. Appl. Biochem. Biotech. 2009, 159, 568–578. [Google Scholar] [CrossRef]
- Ganuza, E.T.; Izquierdo, M.L.; Ratledge, C. High-Cell-Density Cultivation of Schizochytrium Sp in Ammonia-Ph-Auxostat Fed-Batch System. Phycologia 2005, 44, 35–36. [Google Scholar]
- Marques, I.P.; Batista, A.P.; Coelho, A.; da Silva, T.L. Co-digestion of Rhodosporidium toruloides biorefinery wastes for biogas production. Process Biochem. 2018, 64, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.-Y.; Wu, Y.-H.; Wang, J.-H.; Wang, X.-X.; Victor, M.D.-E.; Dao, G.-H.; Tong, X.; Hu, H.-Y. Heterotrophic cultivation of microalgae in straw lignocellulose hydrolysate for production of high-value biomass rich in polyunsaturated fatty acids (PUFA). Chem. Eng. J. 2019, 367, 37–44. [Google Scholar] [CrossRef]
- Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Ornelas-Soto, N.; Romero-Ogawa, M.A.; Parra-Saldivar, R. Extraction and purification of high-value metabolites from microalgae: Essential lipids, astaxanthin and phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, S.P.J.N.; Shahidi, F. Concentration of docosahexaenoic acid (DHA) from algal oil via urea complexation. J. Food Lipids 2000, 7, 51–61. [Google Scholar] [CrossRef]
- Senanayake, S.P.J.N.; Shahidi, F. Enzymatic acidolysis of evening primrose oil with docosahexaenoic acid using response surface methodology. J. Food Lipids 2006, 13, 235–250. [Google Scholar] [CrossRef]
- Mansour, M.P. Reversed-phase high-performance liquid chromatography purification of methyl esters of C-16-C-28 polyunsaturated fatty acids in microalgae, including octacosaoctaenoic acid [28:8(n-3)]. J. Chromatogr. A 2005, 1097, 54–58. [Google Scholar] [CrossRef]
- Li, J.X.; Zhang, P.; Hong, L.; Liu, C.G. Purification and Characterization of a Novel Hydrolase That Can Specifically Degrade the Polysaccharide Isolated from Green Seaweed Ulva prolifera. J. Ocean Univ. China 2019, 18, 185–192. [Google Scholar] [CrossRef]
- Couto, R.M.; Simoes, P.C.; Reis, A.; Da Silva, T.L.; Martins, V.H.; Sanchez-Vicente, Y. Supercritical fluid extraction of lipids from the heterotrophic microalga Crypthecodinium cohnii. Eng. Life Sci. 2010, 10, 158–164. [Google Scholar]
- Adarme-Vega, T.C.; Lim, D.K.Y.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Fact. 2012, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.B.; Wen, Z.Y. Production of Biodiesel Fuel from the Microalga Schizochytrium limacinum by Direct Transesterification of Algal Biomass. Energy Fuel 2009, 23, 5179–5183. [Google Scholar] [CrossRef]
- Chang, K.J.L.; Dumsday, G.; Nichols, P.D.; Dunstan, G.A.; Blackburn, S.I.; Koutoulis, A. High cell density cultivation of a novel Aurantiochytrium sp strain TC 20 in a fed-batch system using glycerol to produce feedstock for biodiesel and omega-3 oils. Appl. Microbiol. Biotechnol. 2013, 97, 6907–6918. [Google Scholar] [CrossRef] [PubMed]
- Maurya, R.; Paliwal, C.; Ghosh, T.; Pancha, I.; Chokshi, K.; Mitra, M.; Ghosh, A.; Mishra, S. Applications of de-oiled microalgal biomass towards development of sustainable biorefinery. Bioresour. Technol. 2016, 214, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Guldhe, A.; Singh, P.; Kumari, S.; Rawat, I.; Permaul, K.; Bux, F. Biodiesel synthesis from microalgae using immobilized Aspergillus niger whole cell lipase biocatalyst. Renew. Energy 2016, 85, 1002–1010. [Google Scholar] [CrossRef]
- Fiori, L.; Volpe, M.; Lucian, M.; Anesi, A.; Manfrini, M.; Guella, G. From Fish Waste to Omega-3 Concentrates in a Biorefinery Concept. Waste Biomass Valori. 2017, 8, 2609–2620. [Google Scholar] [CrossRef]
- Pufachain european project and PUFAChain. GA No. 613303. Available online: https://www.pufachain (accessed on 11 October 2017).
- Keller, H.; Rettenmaier, N.; Reinhardt, G.A. Integrated life cycle sustainability assessment—A practical approach applied to biorefineries. Appl. Energy 2015, 154, 1072–1081. [Google Scholar] [CrossRef]
- Peiris, S.E.; De Silva, E.D.U.D.; Edussuriya, M.; Attanayake, A.M.U.R.K.; Peiris, B.C.N. CSUP technique: A low cost sterilization method using sodium hypochlorite to replace the use of expensive equipment in micropropagation. J. Natl. Sci. Found. Sri Lanka 2012, 40, 49–54. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Feedstock | Microorganism | Cultivation Mode/Time | Lipid Production | EPA/DHA Production | Reference |
|---|---|---|---|---|---|---|
| Pure sources | Glucose | C. cohnii ATCC 30772 | 2 L bioreactor, batch/91 h | 3.79 g·L−1 | 1.6 g·L−1 DHA | [16] |
| Ethanol | C. cohnii ATCC 30772 | 2 L bioreactor, fed-batch/220 h | 42.2% w·w−1, 35 g·L−1 | 11.7 g·L−1 DHA | [17] | |
| Acetate | C. cohnii ATCC 30772 | 2 L bioreactor, fed-batch/120 h | 61.0 g·L−1, 56.0% w·w−1 | 19 g·L−1 DHA | [17] | |
| Glycerol | C. cohnii CCMP 316 | 2 L stirred tank bioreactor/Batch mode/8 days | 2.34 g·L−1, 36.5% w·w−1 | DHA: 49 mg·g−1 | [18] | |
| Food industry effluents/wastes | Food waste hydrolysate | Schizochytrium mangrovei Chlorella pyrenoidosa | 2 L bioreactorBatch mode/7 days | 3.30 g·L−1; 16.49% w·w−1 1.05 g·L−1; 20.99% w·w−1 | 85.5 ± 11.2 mg·g−1 DHA | [19] |
| Sweet sorghum juice | Schizochytrium limacinum | 250 mL flasksBatch mode/5 days | 6.90 g·L−1; 73.4% w·w−1 | 273 mg·g−1 DHA1.1 mg·g−1 EPA | [20] | |
| Carob pulp syrup | C. cohnii CCMP 316 | 2 L bioreactor, fed-batch | 9.2% w·w−1 (as TFA) | 1.99 g·L−1 DHA45.2 mg·g−1 DHA | [21] | |
| Rapeseed meal hydrolysate + crude waste molasses | C. cohnii ATCC 30772 | 500 mL-Erlenmeyers, batch/7 days | 27.3% w·w−1, 26.9 mg·L−1 | 8.72 mg·L−1 DHA22–34 % w·w−1 DHA of TFA | [22] | |
| Cheese whey + Corn steep liquor | Crypthecodinium cohnii CCMP 316 | 250 mL-Erlenmeyers | 28.7% w·w−1 | 8.5–27% w·w−1 DHA of TFA | [18] |
| Biomass | Biorefinery | Processes | Products | Cost | Energy | CO2eq | Reference |
|---|---|---|---|---|---|---|---|
| Fish wastes 871 tonne year−1 | Modeled Aspen Plus™ | oil extraction from fish waste; fish oil trans-esterification with ethanol, and supercritical CO2 fractionation | Proteins for fishmeal (160 tonne year−1)’; Biofuel (160 tonne (year for CHP−1)); ω-3 concentrates for the nutraceutical sector (26.64 tonne year−1 or 30 kg PUFA (kg dry microalgae)−1; PUFA (58% mass fraction in EPA and DHA) | 3.34 M€-Equipment 178 k€ (year utility)−1 | Electricity needs 716 MWh year−1 (100% from biofuel CHP); Heat 1919 MWh year−1 (45% from CHP) | 695 tonne year−1; Or 26 tonne (tonne PUFA)−1 | [59] |
| Phototrophic algae (Prorocentrum cassubicum, Thalassiosira weissflogii and a combination of Chloridella simplex and Raphidonema nivale Lagerheim) 390–4900 ton dry weight year−1 | Modeled industrial scale 10–100 ha of land use | algae production; algae harvesting; cell disruption and spray drying and supercritical CO2-extraction and oil processing | 5–152 tonne PUFA year−1 or 5–9 g PUFA kg dry microalgae−1; extracted cake 21–3800 tonne year−1; oil wastes 8–200 tonne year−1 | Capital cost as CAPEX a 2.6–41.4 M€ year−1; Operational cost as OPEX b 2–31 M€ year−1; 400–1500 € (kg PUFA)−1 | Electricity 1000–95,000 MWh year−1; Heat 80,000–15,000,000 MJ year−1; or 22–4200 MWh year−1 | 1750 tonne (tonne PUFA)−1 or 350 tonne (tonne PUFA)−1 if more solar power is considered; Or 9–16 tonne (tonne autotrophic)−1; Microalgae−1 | [6] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes da Silva, T.; Moniz, P.; Silva, C.; Reis, A. The Dark Side of Microalgae Biotechnology: A Heterotrophic Biorefinery Platform Directed to ω-3 Rich Lipid Production. Microorganisms 2019, 7, 670. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120670
Lopes da Silva T, Moniz P, Silva C, Reis A. The Dark Side of Microalgae Biotechnology: A Heterotrophic Biorefinery Platform Directed to ω-3 Rich Lipid Production. Microorganisms. 2019; 7(12):670. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120670
Chicago/Turabian StyleLopes da Silva, Teresa, Patrícia Moniz, Carla Silva, and Alberto Reis. 2019. "The Dark Side of Microalgae Biotechnology: A Heterotrophic Biorefinery Platform Directed to ω-3 Rich Lipid Production" Microorganisms 7, no. 12: 670. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120670