Stimulated Growth and Innate Immunity in Brook Charr (Salvelinus fontinalis) Treated with a General Probiotic (Bactocell®) and Two Endogenous Probiotics That Inhibit Aeromonas salmonicida In Vitro

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Antagonism Assays
2.1.1. Bacterial Strains and Culture Conditions
2.1.2. Radial Diffusion Assays on Agar
2.2. Comparative Genomic Risk Assessment
2.2.1. DNA Extraction, Sequencing and Annotation
2.2.2. Taxonomic Identification of Probiotic Candidates
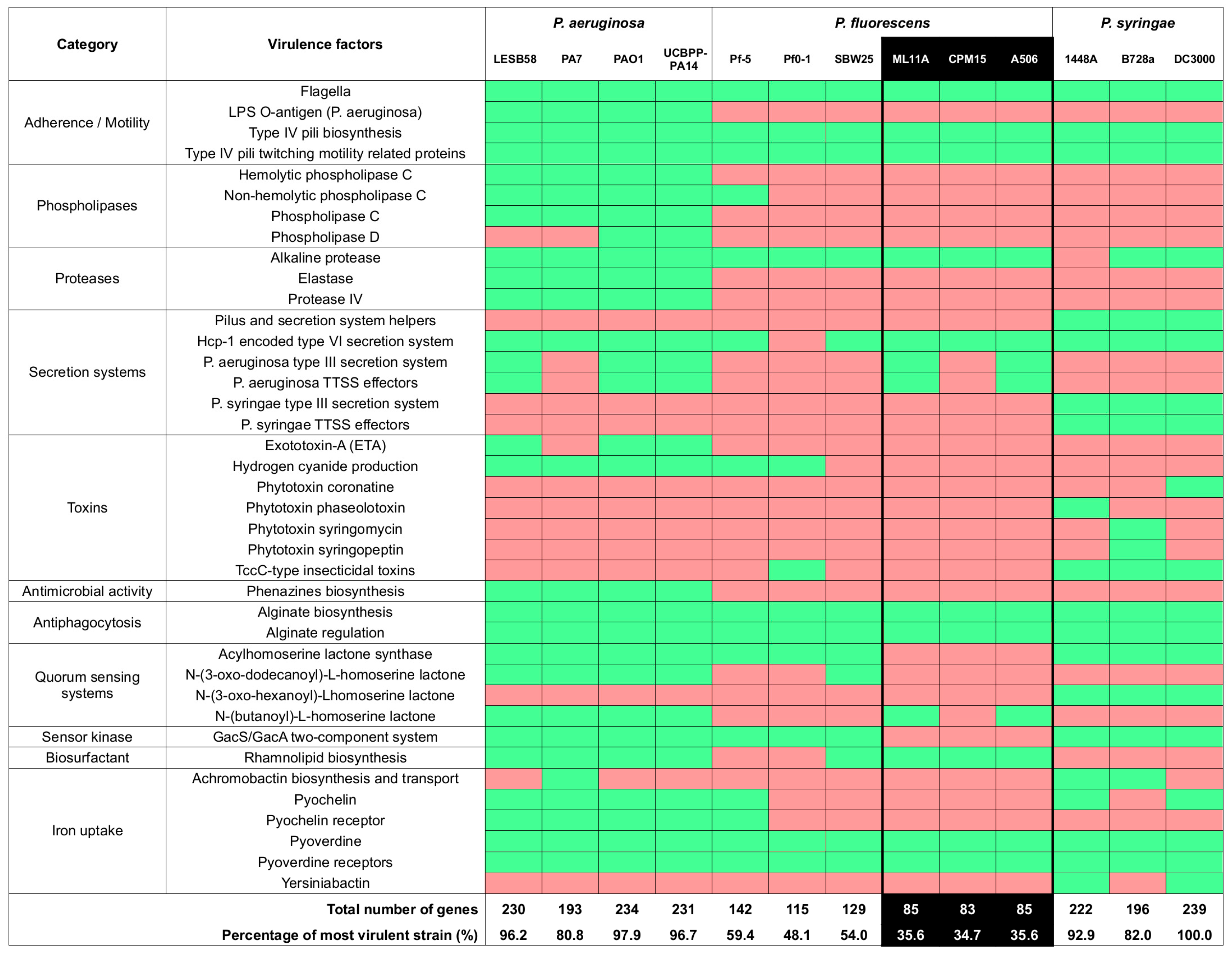
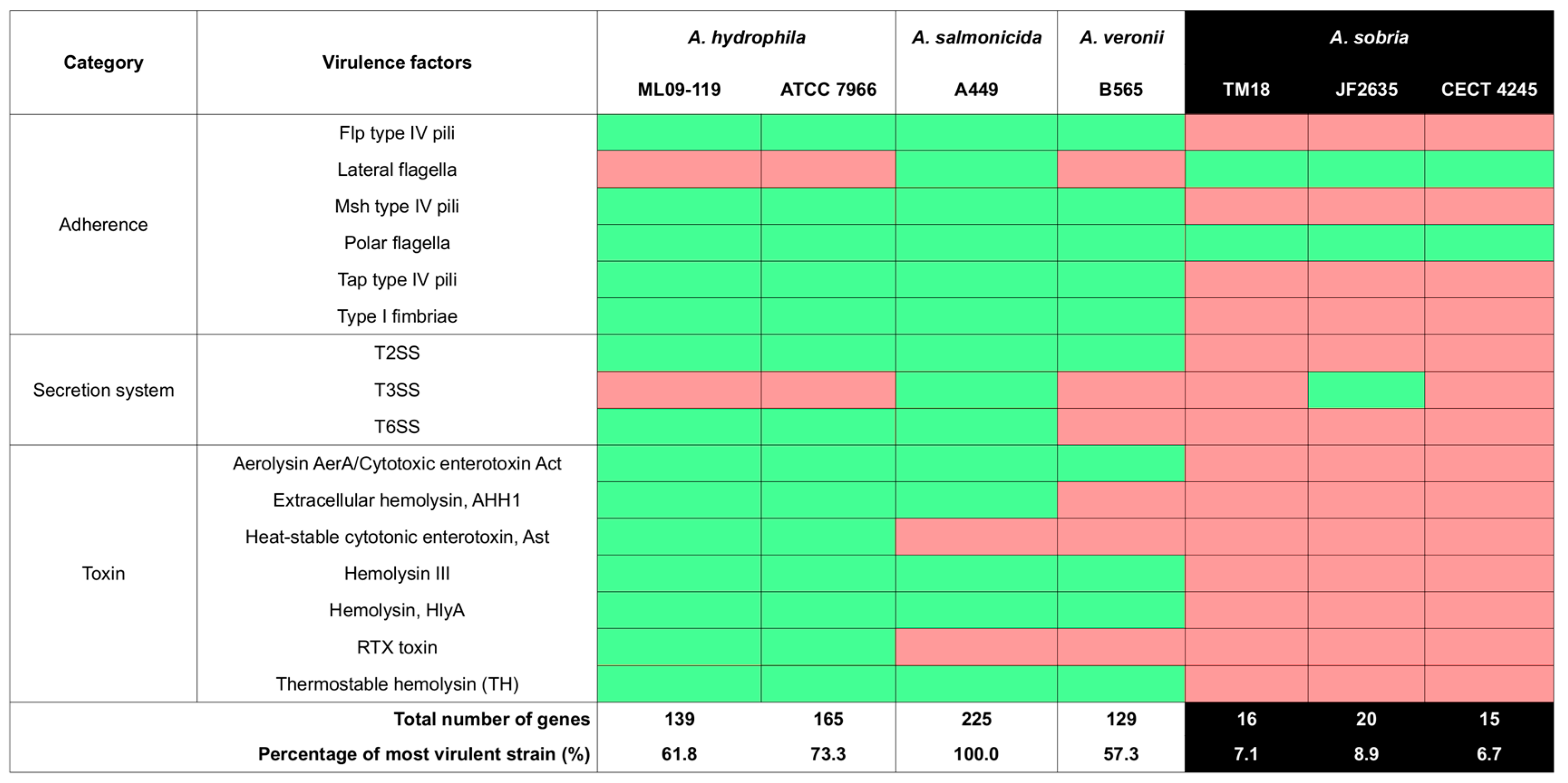
2.2.3. Virulence Factor Gene Contents
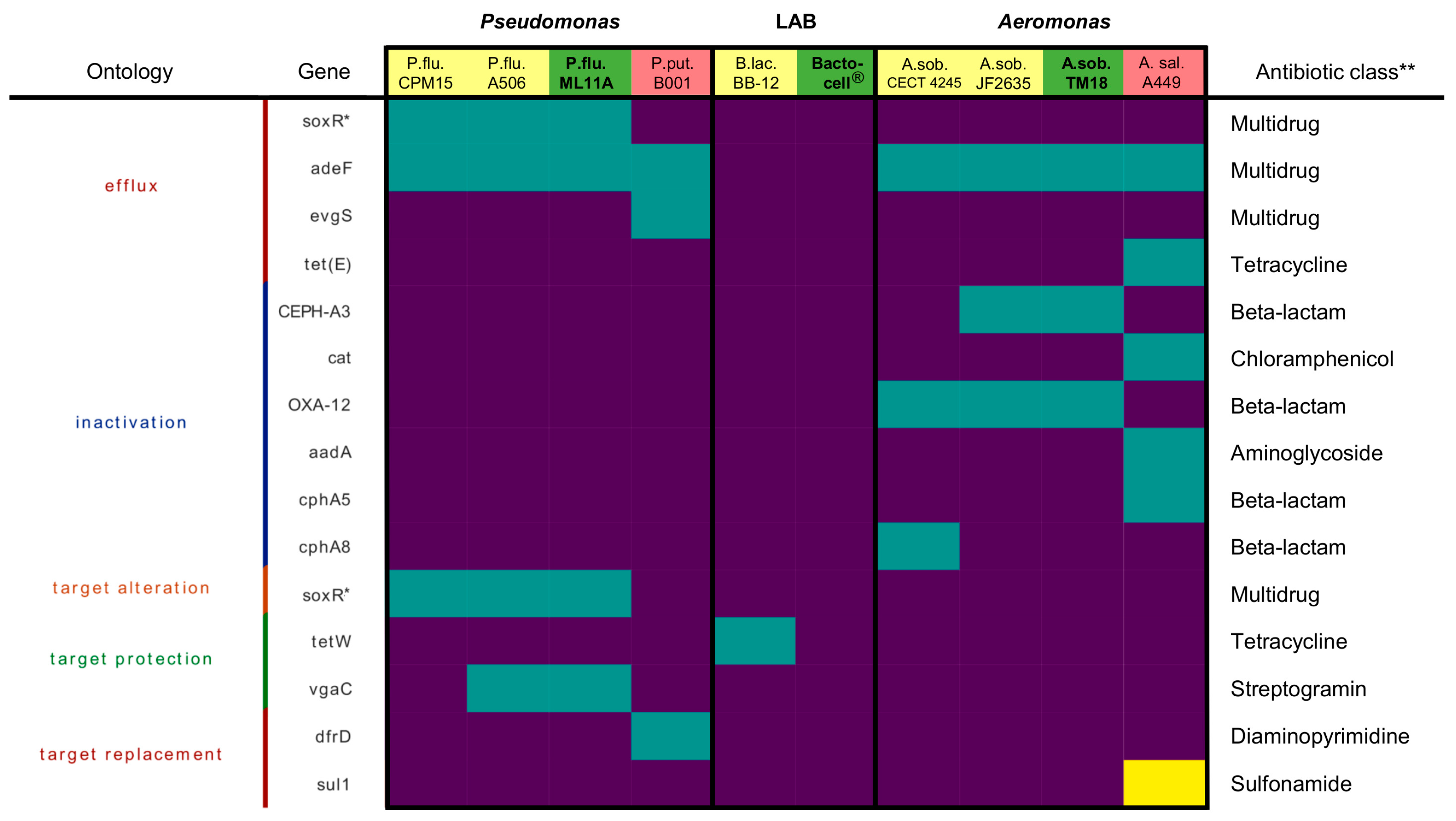
2.2.4. Antibiotic Resistance Genes
2.3. In Vivo Probiotic Innocuity Experiment
2.3.1. Animal Ethics Statement
2.3.2. Brook Charr Genetic Families and Rearing Conditions
2.3.3. Treatment Preparation and Dosage
2.3.4. Experimental Design and Sampling Procedures
2.4. Measurements and Statistical Analysis
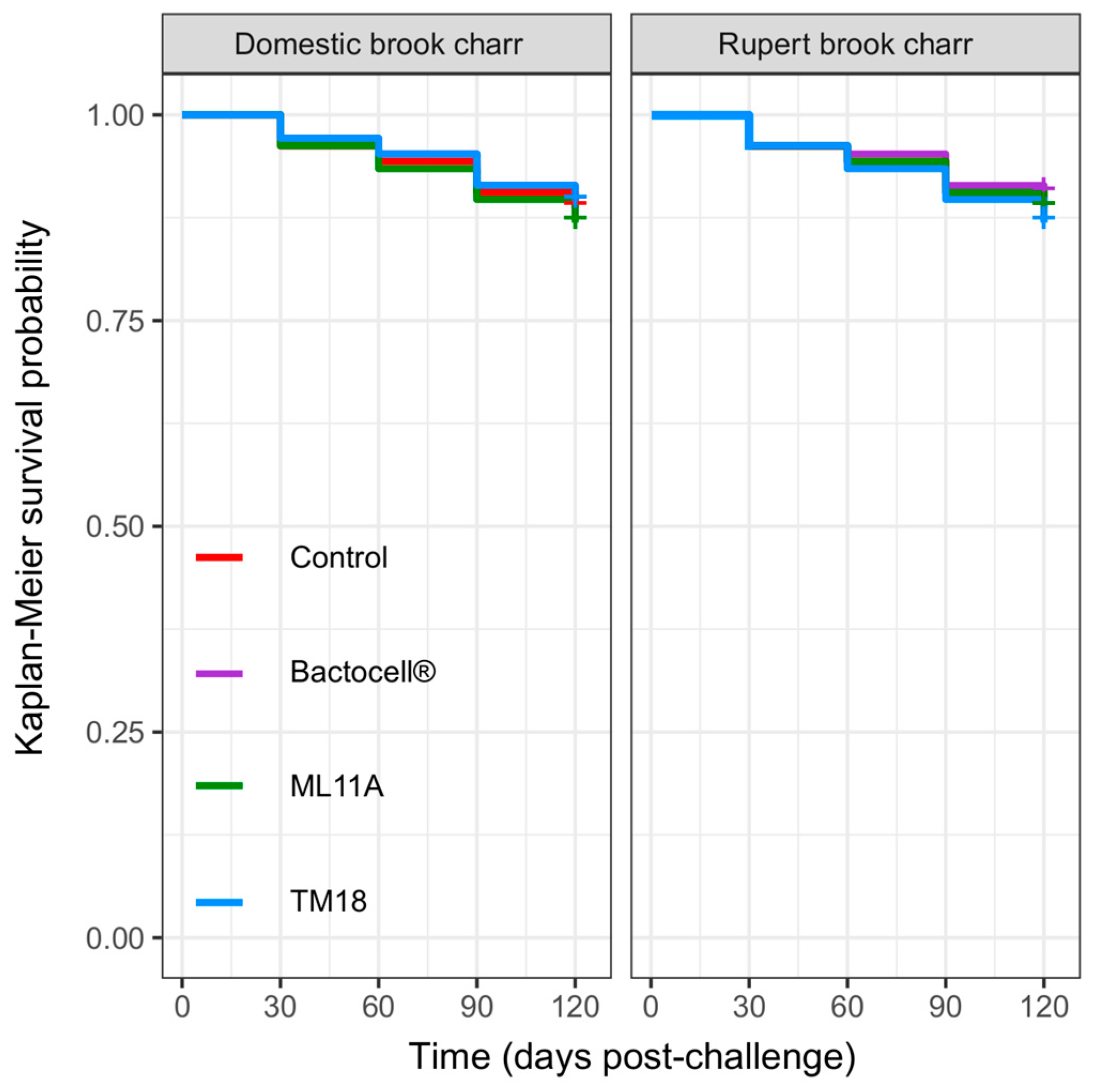
2.4.1. Survival Analysis
2.4.2. Physiological Condition Markers
2.4.3. Statistical Modeling
3. Results
3.1. P. fluorescens ML11A and A. sobria TM18 Inhibit In Vitro Growth of A. salmonicida subsp. salmonicida
3.2. Antimicrobial Resistance Profiles of ML11A, TM18 and Bactocell®
3.3. Innocuity of ML11A, TM18 and Bactocell® toward Brook Charr
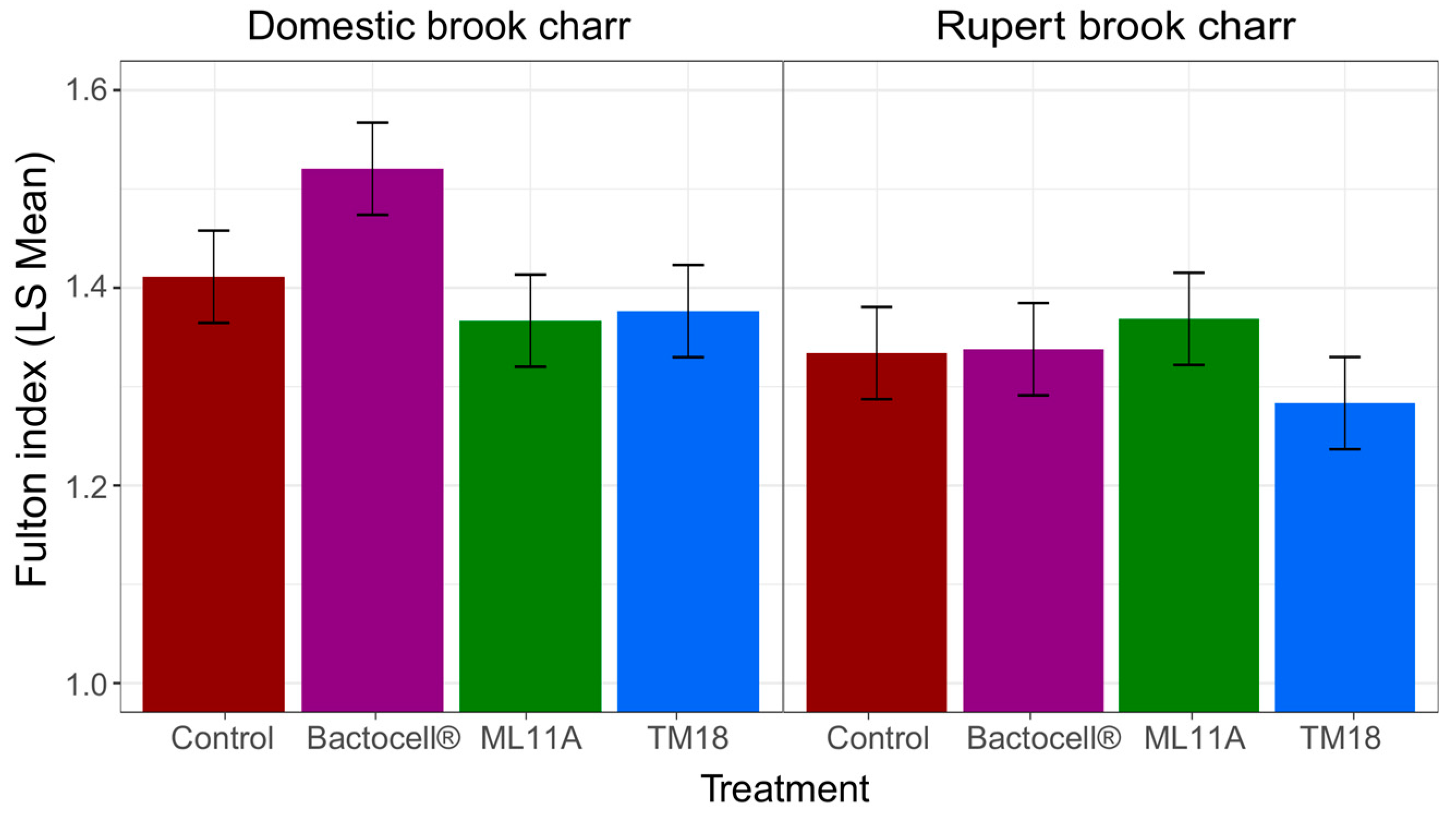
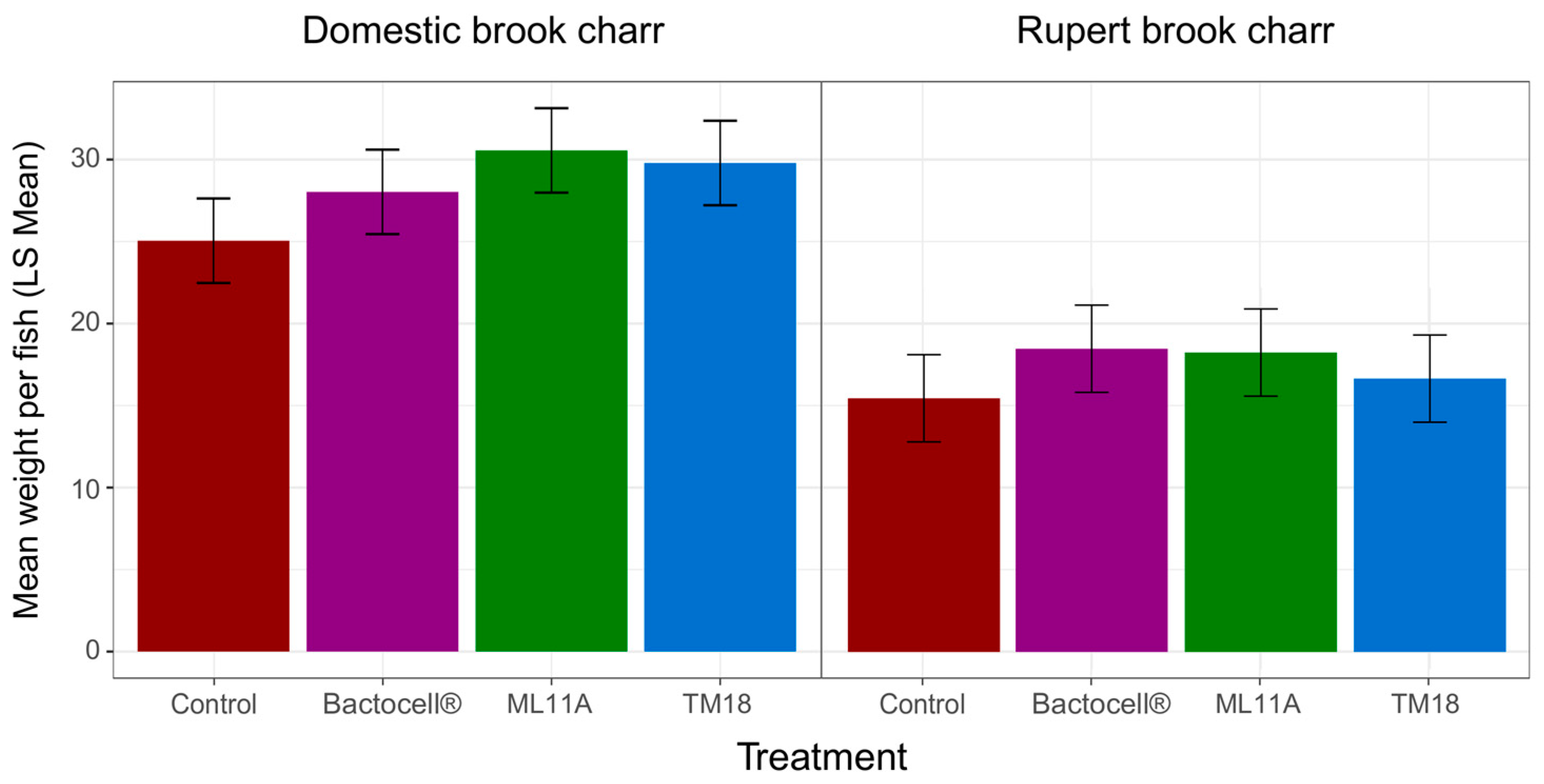
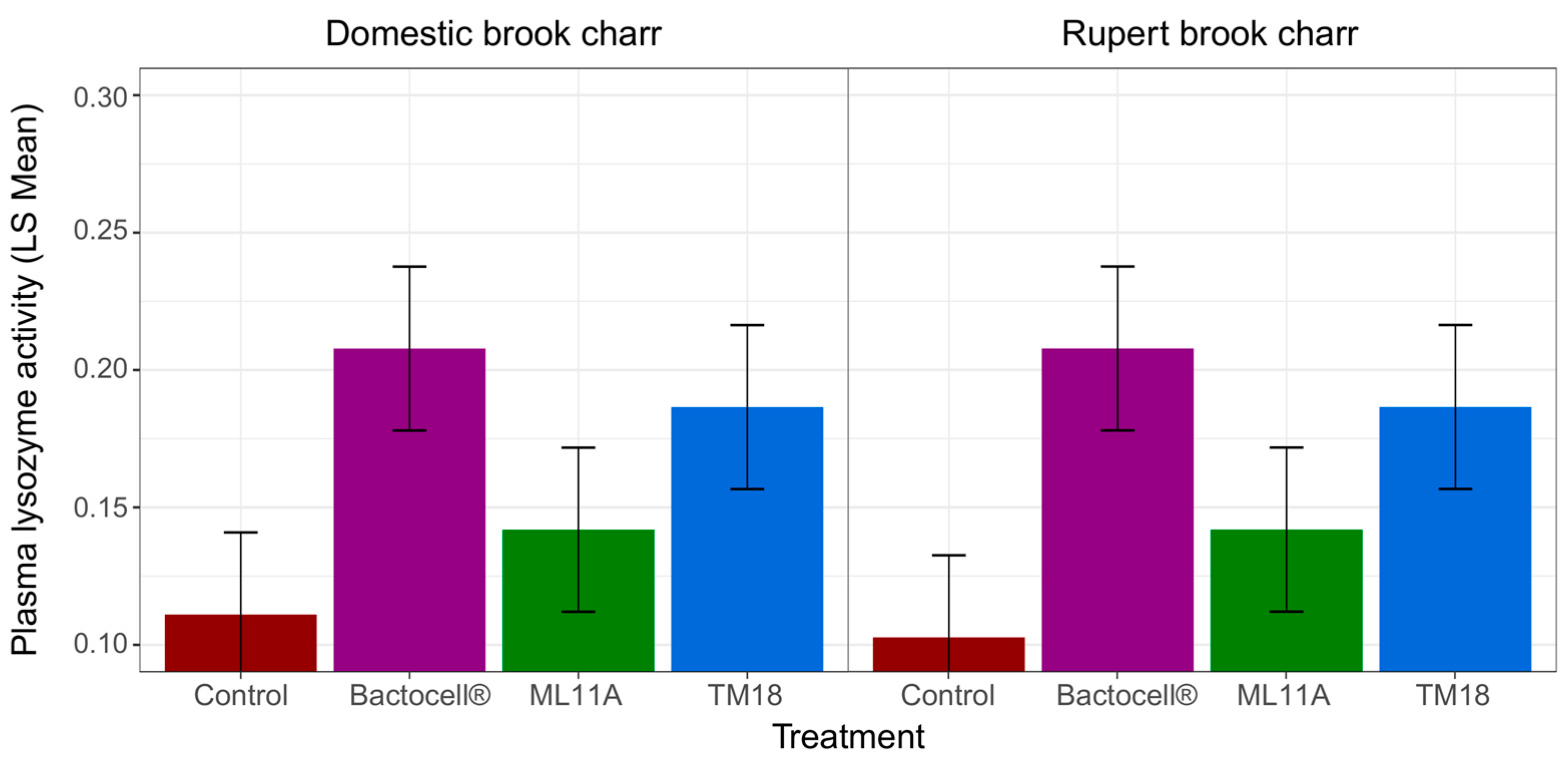
3.4. Effect of ML11A, TM18 and Bactocell® on Brook Charr Physiological Condition, Weight Gain and Plasma Lysozyme Activity
4. Discussion
4.1. ML11A and TM18 Inhibit A. salmonicida on Agar via Diffusible Compounds
4.2. ML11A and TM18 Lack Key Virulence Factors Found in Related Pathogenic Strains
- In P. aeruginosa, an opportunistic human pathogen [50]: the LPS O-antigen, phospholipases C and D, elastase, protease IV, exotoxin A, hydrogen cyanide production, as well as several quorum sensing systems.
- In P. syringae, a well-known plant pathogen [51]: the P. syringae variant of the type three secretion system (TTSS) and its effectors, phytotoxins coronatine, phaseolotoxin, syringomycin and syringopeptin, the acylhomoserine and N-(3-oxo-hexanoyl)-L-homoserine lactone quorum sensing systems, and achromobactin biosynthesis and transport.
4.3. ML11A, TM18 and Bactocell® Lack Genes Conferring Resistance to Key Antimicrobials in Salmonid Aquaculture
4.4. Growth Promotion and Immune Modulation by Probionts Bactocell, ML11A and TM18
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Austin, B.; Austin, D.A. Aeromonadaceae Representative (Aeromonas salmonicida). In Bacterial Fish Pathogens; Springer: Berlin/Heidelberg, Germany, 2012; pp. 147–228. [Google Scholar]
- Cipriano, R.C.; Bullock, G.L. Furunculosis and Other Diseases Caused by Aeromonas salmonicida; U.S. Fish and Wildlife Service: Washington, DC, USA, 2001; pp. 1–33.
- Pickering, A.D. Husbandry and Stress. In Furunculosis; Bernoth, E.-M., Ellis, A.E., Midtlyng, P.J., Olivier, G., Smith, P., Eds.; Academic Press: Cambridge, MA, USA, 1997; pp. 178–202. [Google Scholar]
- Scott, M. The Pathogenicity of Aeromonas salmonicida (Griffin) in Sea and Brackish Waters. J. Gen. Microbiol. 1968, 50, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Samuelsen, O.B.; Hjeltnes, B.; Glette, J. Efficacy of Orally Administered Florfenicol in the Treatment of Furunculosis in Atlantic Salmon. J. Aquatic Anim. Health 1998, 10, 56–61. [Google Scholar] [CrossRef]
- Amend, D.F. Comparison of certain sulfonamide drugs for treatment of bacterial infections of Pacific salmon. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1965. [Google Scholar]
- Morin, R. The use of antibiotics to fight furunculosis in brook charr generates resistance in Aeromonas salmonicida. Que. Fish Farmers Assoc. Bull. 2010, 15, 3–6. [Google Scholar]
- World Health Organization. Antimicrobial resistance: Global Report on Surveillance. Available online: https://apps.who.int/iris/bitstream/handle/10665/112642/9789241564748_eng.pdf (accessed on 7 July 2019).
- Inglis, V.; Yimer, E.; Bacon, E.J.; Ferguson, S. Plasmid-mediated antibiotic resistance in Aeromonas salmonicida isolated from Atlantic salmon, Salmo salar L., in Scotland. J. Fish Dis. 1993, 16, 593–599. [Google Scholar] [CrossRef]
- Reith, M.E.; Singh, R.K.; Curtis, B.; Boyd, J.M.; Bouevitch, A.; Kimball, J.; Munholland, J.; Murphy, C.; Sarty, D.; Williams, J.; et al. The genome of Aeromonas salmonicida subsp. salmonicida A449: Insights into the evolution of a fish pathogen. BMC Genom. 2008, 9, 427. [Google Scholar] [CrossRef]
- Dallaire-Dufresne, S.; Tanaka, K.H.; Trudel, M.V.; Lafaille, A.; Charette, S.J. Virulence, genomic features, and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Vet. Microbiol. 2014, 169, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, D.; Cunningham, M.; Ji, B.; Fekete, F.A.; Parry, E.M.; Clark, S.E.; Zalinger, Z.B.; Gilg, I.C.; Danner, G.R.; Johnson, K.A.; et al. Transferable, multiple antibiotic and mercury resistance in Atlantic Canadian isolates of Aeromonas salmonicida subsp. salmonicida is associated with carriage of an IncA/C plasmid similar to the Salmonella enterica plasmid pSN254. J. Antimicrob. Chemother. 2008, 61, 1221–1228. [Google Scholar] [CrossRef]
- Vincent, A.T.; Trudel, M.V.; Paquet, V.E.; Boyle, B.; Tanaka, K.H.; Dallaire-Dufresne, S.; Daher, R.K.; Frenette, M.; Derome, N.; Charette, S.J. Detection of Variants of the pRAS3, pAB5S9, and pSN254 Plasmids in Aeromonas salmonicida subsp. salmonicida: Multidrug Resistance, Interspecies Exchanges, and Plasmid Reshaping. Antimicrob. Agents Chemother. 2014, 58, 7367–7374. [Google Scholar]
- Vincent, A.T.; Emond-Rheault, J.-G.; Barbeau, X.; Attéré, S.A.; Frenette, M.; Lagüe, P.; Charette, S.J. Antibiotic resistance due to an unusual ColE1-type replicon plasmid in Aeromonas salmonicida. Microbiology 2016, 162, 942–953. [Google Scholar] [CrossRef]
- Trudel, M.V.; Vincent, A.T.; Attéré, S.A.; Labbé, M.; Derome, N.; Culley, A.I.; Charette, S.J. Diversity of antibiotic-resistance genes in Canadian isolates of Aeromonas salmonicida subsp. salmonicida: Dominance of pSN254b and discovery of pAsa8. Sci. Rep. 2016, 6, 35617. [Google Scholar]
- Barton, B.A.; Iwama, G.K. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Annu. Rev. Fish Dis. 1991, 1, 3–26. [Google Scholar] [CrossRef]
- Boutin, S.; Bernatchez, L.; Audet, C.; Derôme, N. Network analysis highlights complex interactions between pathogen, host and commensal microbiota. PLoS ONE 2013, 8, e84772. [Google Scholar] [CrossRef] [PubMed]
- Stecher, B.; Maier, L.; Hardt, W.-D. “Blooming” in the gut: How dysbiosis might contribute to pathogen evolution. Nat. Rev. Microbiol. 2013, 11, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Gauthier, J.; Charette, S.J.; Derome, N. Chapter 1: The rise and fall of antibiotics in aquaculture. In Microbial Communities in Aquaculture Ecosystems: Improving Productivity and Sustainability; Derome, N., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 1–20. [Google Scholar]
- Boutin, S.; Bernatchez, L.; Audet, C.; Derôme, N. Antagonistic effect of indigenous skin bacteria of brook charr (Salvelinus fontinalis) against Flavobacterium columnare and F. psychrophilum. Vet. Microbiol. 2012, 155, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Boutin, S.; Audet, C.; Derôme, N. Probiotic treatment by indigenous bacteria decreases mortality without disturbing the natural microbiota of Salvelinus fontinalis. Can. J. Microbiol. 2013, 59, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Goulden, E.F.; Hall, M.R.; Pereg, L.L.; Høj, L. Identification of an Antagonistic Probiotic Combination Protecting Ornate Spiny Lobster (Panulirus ornatus) Larvae against Vibrio owensii Infection. PLoS ONE 2012, 7, e39667. [Google Scholar] [CrossRef] [PubMed]
- Schubiger, C.B.; Orfe, L.H.; Sudheesh, P.S.; Cain, K.D.; Shah, D.H.; Call, D.R. Entericidin Is Required for a Probiotic Treatment (Enterobacter sp. Strain C6-6) To Protect Trout from Cold-Water Disease Challenge. Appl. Environ. Microbiol. 2015, 81, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Núñez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion on the efficacy of Bactocell (Pediococcus acidilactici) when used as a feed additive for fish. EFSA J. 2012, 10, 2886. [Google Scholar] [CrossRef]
- Gauthier, J.; Charette, S.J.; Derome, N. Draft Genome Sequence of Pseudomonas fluorescens ML11A, an Endogenous Strain from Brook Charr with Antagonistic Properties against Aeromonas salmonicida subsp. salmonicida. Genome Announc. 2017, 5. [Google Scholar] [CrossRef]
- Gauthier, J.; Vincent, A.T.; Charette, S.J.; Derome, N. Strong Genomic and Phenotypic Heterogeneity in the Aeromonas sobria Species Complex. Front. Microbiol. 2017, 8, 2434. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, J. Développement D’outils Thérapeutiques à Base de Probiotiques Endogènes Contre la Furonculose (Aeromonas salmonicida) chez L’Omble de Fontaine D’aquaculture (Phase 1- In Vitro). Available online: https://corpus.ulaval.ca/jspui/handle/20.500.11794/27594 (accessed on 7 July 2019).
- Wahli, T.; Burr, S.E.; Pugovkin, D.; Mueller, O.; Frey, J. Aeromonas sobria, a causative agent of disease in farmed perch, Perca fluviatilis L. J. Fish Dis. 2005, 28, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.; Véron, M. A Taxonomic Study of the Aeromonas hydrophila-Aeromonas punctata Group. Microbiology 1976, 94, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Barreau, G.; Tompkins, T.A.; Carvalho, V.G. de Draft Genome Sequence of Probiotic Strain Pediococcus acidilactici MA18/5M. J. Bacteriol. 2012, 194, 901. [Google Scholar] [CrossRef] [PubMed]
- Michel, C. Furunculosis of salmonids: Vaccination attempts in rainbow trout (Salmo gairdneri) by formalin-killed germs. Ann. Rech. Vet. 1979, 10, 33–40. [Google Scholar] [PubMed]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Informations sur les Poissons—Centre de Transfert et de Sélection des Salmonidés (CTSS) inc. Available online: http://www.ctss-genetique.com/Informations-sur-les-poissons (accessed on 15 May 2019).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: ftp://ftp.uvigo.es/CRAN/web/packages/dplR/vignettes/intro-dplR.pdf (accessed on 7 July 2019).
- RStudio Team. RStudio: Integrated Development Environment for R. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.651.1157&rep=rep1&type=pdf#page=14 (accessed on 7 July 2019).
- Therneau, T.M. A Package for Survival Analysis in S. Available online: https://cran.r-project.org/web/packages/survival/survival.pdf (accessed on 5 July 2019).
- Kassambara, A.; Kosinski, M. Survminer: Drawing Survival Curves Using “ggplot2”. Available online: https://cran.r-project.org/web/packages/survminer/survminer.pdf (accessed on 5 July 2019).
- Fulton, T.W. Rate of Growth of Sea Fishes; Neill & Co.: Edinburgh, UK, 1902; pp. 326–439. [Google Scholar]
- Massart, S.; Redivo, B.; Flamion, E.; Mandiki, S.N.M.; Falisse, E.; Milla, S.; Kestemont, P. The trenbolone acetate affects the immune system in rainbow trout, Oncorhynchus mykiss. Aquat. Toxicol. 2015, 163, 109–120. [Google Scholar] [CrossRef]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–213. [Google Scholar]
- Charette, S.J.; Brochu, F.; Boyle, B.; Filion, G.; Tanaka, K.H.; Derome, N. Draft Genome Sequence of the Virulent Strain 01-B526 of the Fish Pathogen Aeromonas salmonicida. J. Bacteriol. 2012, 194, 722–723. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bergh, P.; Heller, M.; Braga-Lagache, S.; Frey, J. The Aeromonas salmonicida subsp. salmonicida exoproteome: Global analysis, moonlighting proteins and putative antigens for vaccination against furunculosis. Proteome Sci. 2013, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- O’Regan, E.; Quinn, T.; Pagès, J.-M.; McCusker, M.; Piddock, L.; Fanning, S. Multiple regulatory pathways associated with high-level ciprofloxacin and multidrug resistance in Salmonella enterica serovar Enteritidis: Involvement of RamA and other global regulators. Antimicrob. Agents Chemother. 2009, 53, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Coyne, S.; Rosenfeld, N.; Lambert, T.; Courvalin, P.; Périchon, B. Overexpression of resistance-nodulation-cell division pump AdeFGH confers multidrug resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4389–4393. [Google Scholar] [CrossRef] [PubMed]
- Novotna, G.; Janata, J. A new evolutionary variant of the streptogramin A resistance protein, Vga(A)LC, from Staphylococcus haemolyticus with shifted substrate specificity towards lincosamides. Antimicrob. Agents Chemother. 2006, 50, 4070–4076. [Google Scholar] [CrossRef] [PubMed]
- El Zowalaty, M.E.; Al Thani, A.A.; Webster, T.J.; El Zowalaty, A.E.; Schweizer, H.P.; Nasrallah, G.K.; Marei, H.E.; Ashour, H.M. Pseudomonas aeruginosa: Arsenal of resistance mechanisms, decades of changing resistance profiles, and future antimicrobial therapies. Future Microbiol. 2015, 10, 1683–1706. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- BlightBan® A506. Available online: https://www2.nufarm.com/uscrop/product/blightban-a506/ (accessed on 19 May 2019).
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J.E. Control of fire blight by Pseudomonas fluorescens A506 and Pantoea vagans C9-1 applied as single strains and mixed inocula. Phytopathology 2010, 100, 1330–1339. [Google Scholar] [CrossRef]
- Ma, Q.; Zhai, Y.; Schneider, J.C.; Ramseier, T.M.; Saier, M.H. Protein secretion systems of Pseudomonas aeruginosa and P. fluorescens. Biochim. Biophys. Acta 2003, 1611, 223–233. [Google Scholar] [CrossRef]
- Loper, J.E.; Kobayashi, D.Y.; Paulsen, I.T. The Genomic Sequence of Pseudomonas fluorescens Pf-5: Insights Into Biological Control. Phytopathology 2007, 97, 233–238. [Google Scholar] [CrossRef]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef]
- Health Canada List of Veterinary Drugs That Are Authorized for Sale by Health Canada for Use in Food-Producing Aquatic Animals—Health Canada. Available online: https://www.canada.ca/en/health-canada/services/drugs-health-products/veterinary-drugs/legislation-guidelines/policies/list-veterinary-drugs-that-authorized-sale-health-canada-use-food-producing-aquatic-animals.html (accessed on 5 July 2019).
- Weir, M.; Rajić, A.; Dutil, L.; Cernicchiaro, N.; Uhland, F.C.; Mercier, B.; Tuševljak, N. Zoonotic bacteria, antimicrobial use and antimicrobial resistance in ornamental fish: A systematic review of the existing research and survey of aquaculture-allied professionals. Epidemiol. Infect. 2012, 140, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Tello, A.; Austin, B.; Telfer, T.C. Selective Pressure of Antibiotic Pollution on Bacteria of Importance to Public Health. Environ. Health Perspect 2012, 120, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Han, S.H.; Lee, J.H.; Han, Y.S.; Lee, Y.S.; Rong, X.; McSpadden Gardener, B.B.; Park, H.-S.; Kim, Y.C. Draft genome sequence of the biocontrol bacterium Pseudomonas putida B001, an oligotrophic bacterium that induces systemic resistance to plant diseases. J. Bacteriol. 2011, 193, 6795–6796. [Google Scholar] [CrossRef]
- Jungersen, M.; Wind, A.; Johansen, E.; Christensen, J.E.; Stuer-Lauridsen, B.; Eskesen, D. The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms 2014, 2, 92–110. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Melville, C.M.; Barbosa, T.M.; Flint, H.J. Occurrence of the new tetracycline resistance gene tet(W) in bacteria from the human gut. Antimicrob. Agents Chemother. 2000, 44, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Oliver, W.T.; Wells, J.E. Lysozyme as an alternative to growth promoting antibiotics in swine production. J. Anim. Sci. Biotechnol. 2015, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Host of Origin | Source | Known Biological Roles | Reference |
|---|---|---|---|---|---|
| Pseudomonas fluorescens | ML11A | Brook charr (Salvelinus fontinalis) | Skin mucus | Candidate probiotic | [27] |
| CPM15 (control) | Brook charr (Salvelinus fontinalis) | Skin mucus | Candidate probiotic | [29] | |
| Aeromonas sobria | TM18 | Brook charr (Salvelinus fontinalis) | Gut contents | Candidate probiotic | [28] |
| JF2635 (control) | European perch (Perca fluviatilis) | Skin | Clinical sample | [30] | |
| CECT 4245 (control) | Fish (unknown) | Unknown | Unknown | [31] | |
| Pediococcus acidilactici | MA18/5M (Bactocell) | Natural pasture Gramineae | Unknown | Probiotic, silage inoculant | [32] |
| A. salmonicida subsp. salmonicida | Inhibition Surface (mm2) | |||||||
|---|---|---|---|---|---|---|---|---|
| P. fluorescens | A. sobria | P. acidilactici | ||||||
| Country of Origin | Strain | Source and/or Reference * | ML11A | CPM15 | TM18 | JF2635 | CECT 4245T | MA18/5M (Bactocell®) ** |
| Quebec, Canada | 01-B522 | FMVUM | ++ | - | +++ | - | - | - |
| 01-B526 | FMVUM [45] | ++ | - | ++ | - | - | - | |
| 09-0167 | FMVUM | +++ | - | ++++ | - | - | - | |
| M15879-11 | FMVUM | +++ | - | + | - | - | - | |
| m23067-09 | FMVUM | +++ | - | ++++ | - | - | - | |
| New Brunswick, Canada | 04-05MF26 | FOC | +++ | - | ++++ | - | - | - |
| 09-144K3 | FOC | +++ | - | +++ | - | - | - | |
| Norway | HER1085 | FHRC | +++ | - | ++++ | - | - | - |
| Switzerland | JF2267 | [46] | +++ | - | + | - | - | - |
| France | A449T | [33] | ++ | - | +++ | - | - | - |
| Strain | Inhibition Diameter (mm) * | ||||
|---|---|---|---|---|---|
| TET | CHL | NAL | SXT | FFC | |
| P. fluorescens Iso11A | +++ | + | +++ | ++ | R |
| A. sobria TM18 | +++ | +++ | +++ | +++ | +++ |
| P. acidilactici MA18/5M (Bactocell®) | +++ | +++ | R | R | +++ |
| A. salmonicida 01-B526 (sensitive control) | +++ | +++ | +++ | +++ | +++ |
| Factor | d.f. | F-Ratio | p-Value |
|---|---|---|---|
| Treatment (ML11A, TM18, Bactocell® or Control) | 3 | 6.0276 | 0.0007 |
| Lineage (Domestic or Rupert) | 1 | 27.6307 | 5 × 10−7 |
| Treatment-Species interaction | 3 | 5.1425 | 0.002 |
| Residuals | 136 | - | - |
| Factor | d.f. | F-Ratio | p-Value |
|---|---|---|---|
| Treatment (ML11A, TM18, Bactocell® or Control) | 3 | 3.6277 | 0.01 |
| Lineage (Domestic or Rupert) | 1 | 160.8322 | Below 10−16 |
| Treatment-Species interaction | 3 | 1.0138 | 0.4 |
| Residuals | 136 | - | - |
| Factor | d.f. | Kruskal-Wallis X2 | p-Value |
|---|---|---|---|
| Treatment (ML11A, TM18, Bactocell® or Control) | 3 | 33.801 | 2.183 × 10-7 |
| Lineage (Domestic or Rupert) | 1 | 0.0067 | 0.9347 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gauthier, J.; Rouleau-Breton, S.; J. Charette, S.; Derome, N. Stimulated Growth and Innate Immunity in Brook Charr (Salvelinus fontinalis) Treated with a General Probiotic (Bactocell®) and Two Endogenous Probiotics That Inhibit Aeromonas salmonicida In Vitro. Microorganisms 2019, 7, 193. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070193
Gauthier J, Rouleau-Breton S, J. Charette S, Derome N. Stimulated Growth and Innate Immunity in Brook Charr (Salvelinus fontinalis) Treated with a General Probiotic (Bactocell®) and Two Endogenous Probiotics That Inhibit Aeromonas salmonicida In Vitro. Microorganisms. 2019; 7(7):193. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070193
Chicago/Turabian StyleGauthier, Jeff, Stéphanie Rouleau-Breton, Steve J. Charette, and Nicolas Derome. 2019. "Stimulated Growth and Innate Immunity in Brook Charr (Salvelinus fontinalis) Treated with a General Probiotic (Bactocell®) and Two Endogenous Probiotics That Inhibit Aeromonas salmonicida In Vitro" Microorganisms 7, no. 7: 193. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070193