Metabolic Profiling and Cold-Starvation Stress Response of Oxygen-Tolerant Lactobacillus gasseri Strains Cultured in Batch Bioreactor


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Batch bioreactor cultivation
2.3. Bacterial Growth Monitoring
2.4. HPLC Analysis
2.5. Tolerance to Cold-Starvation Stress
2.6. Comparative in Silico Analysis
2.7. Data Analysis
3. Results
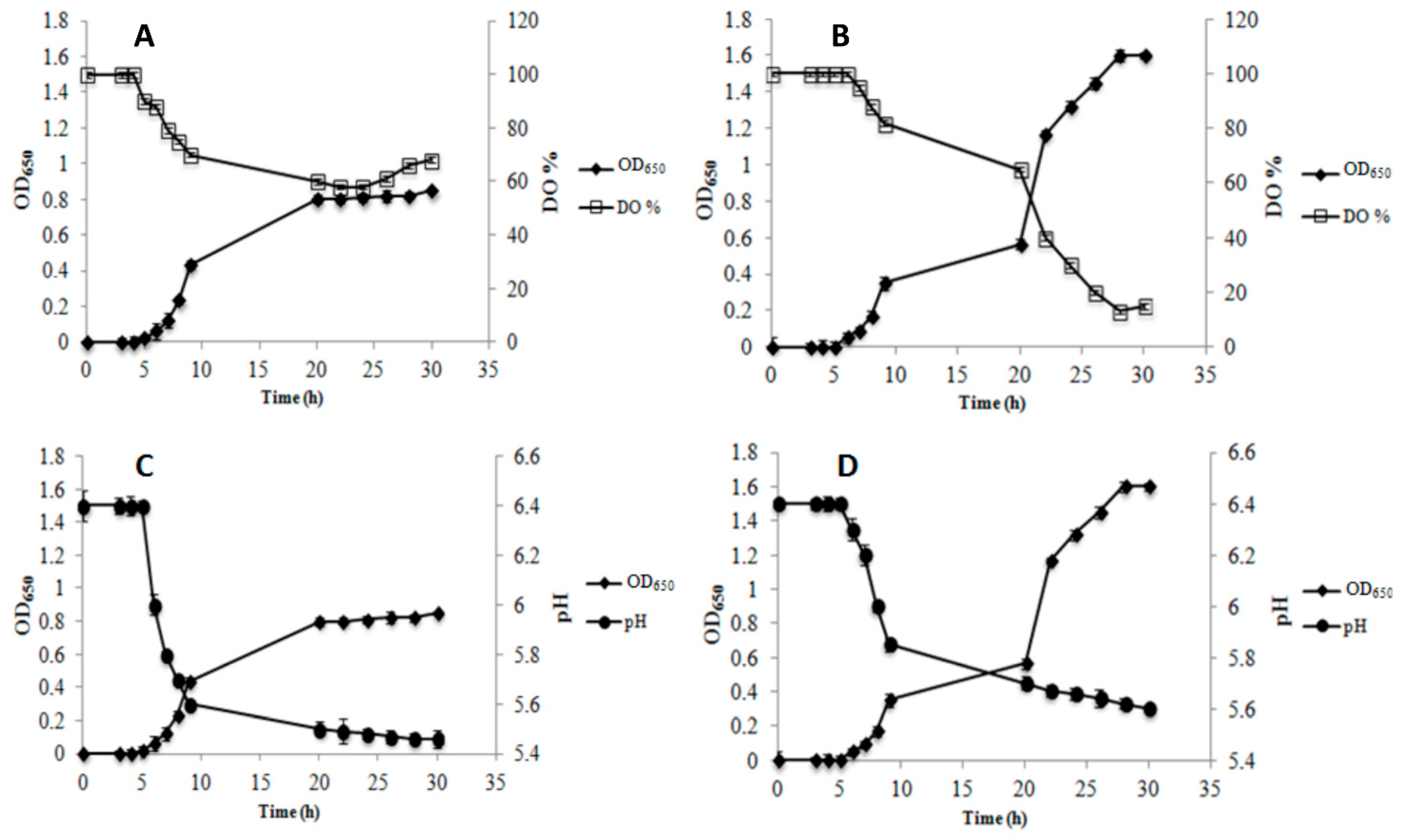
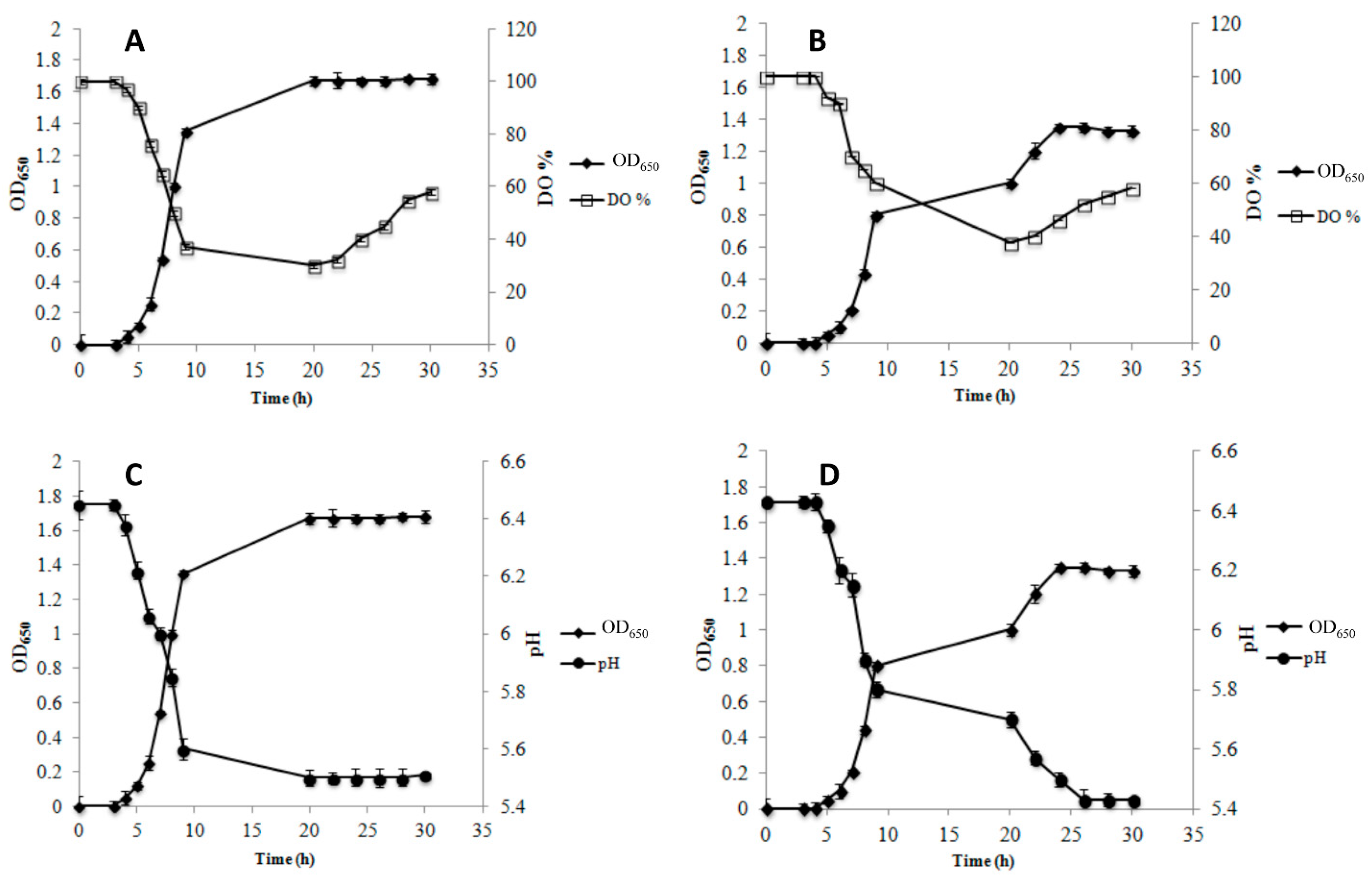
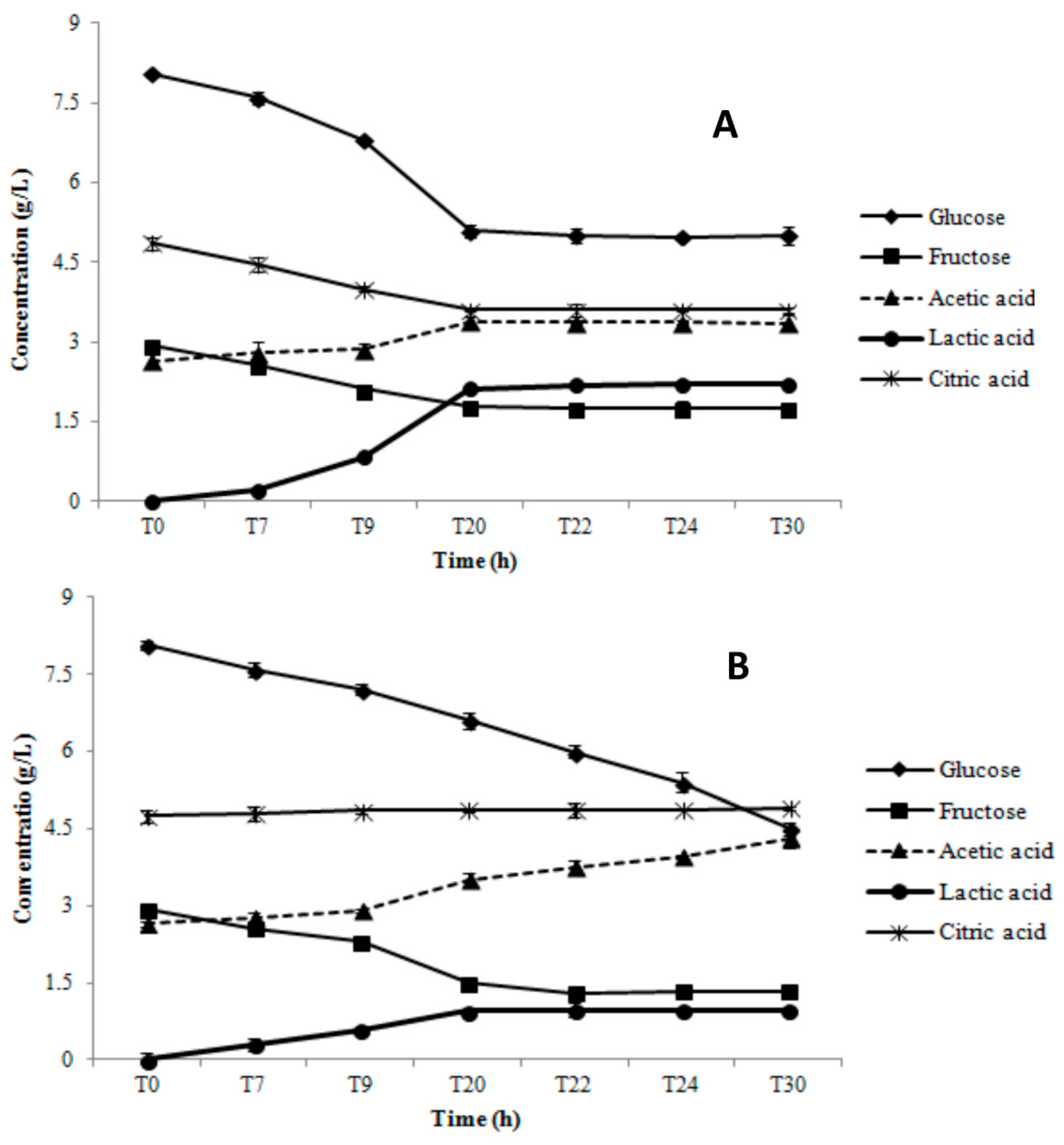
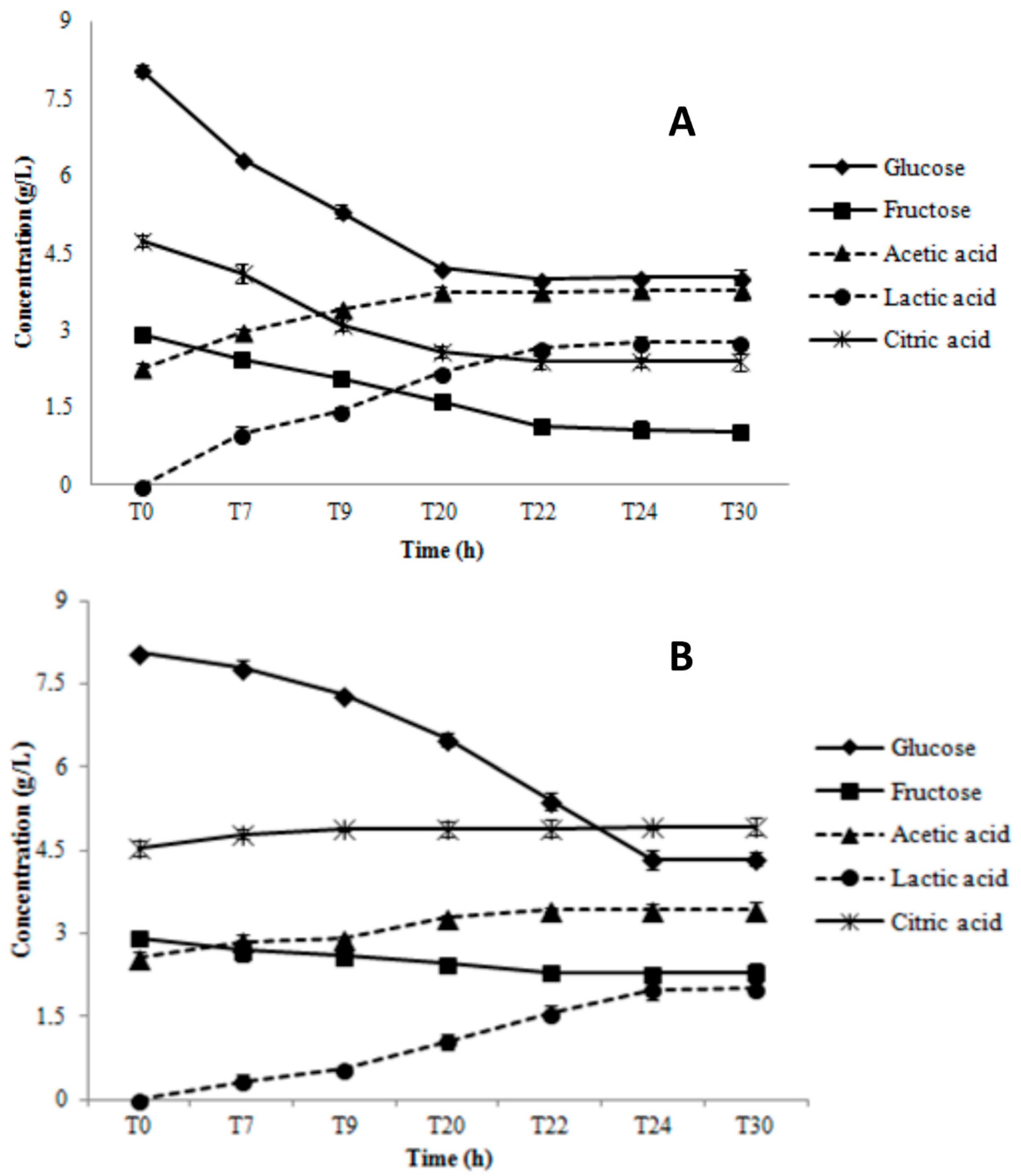
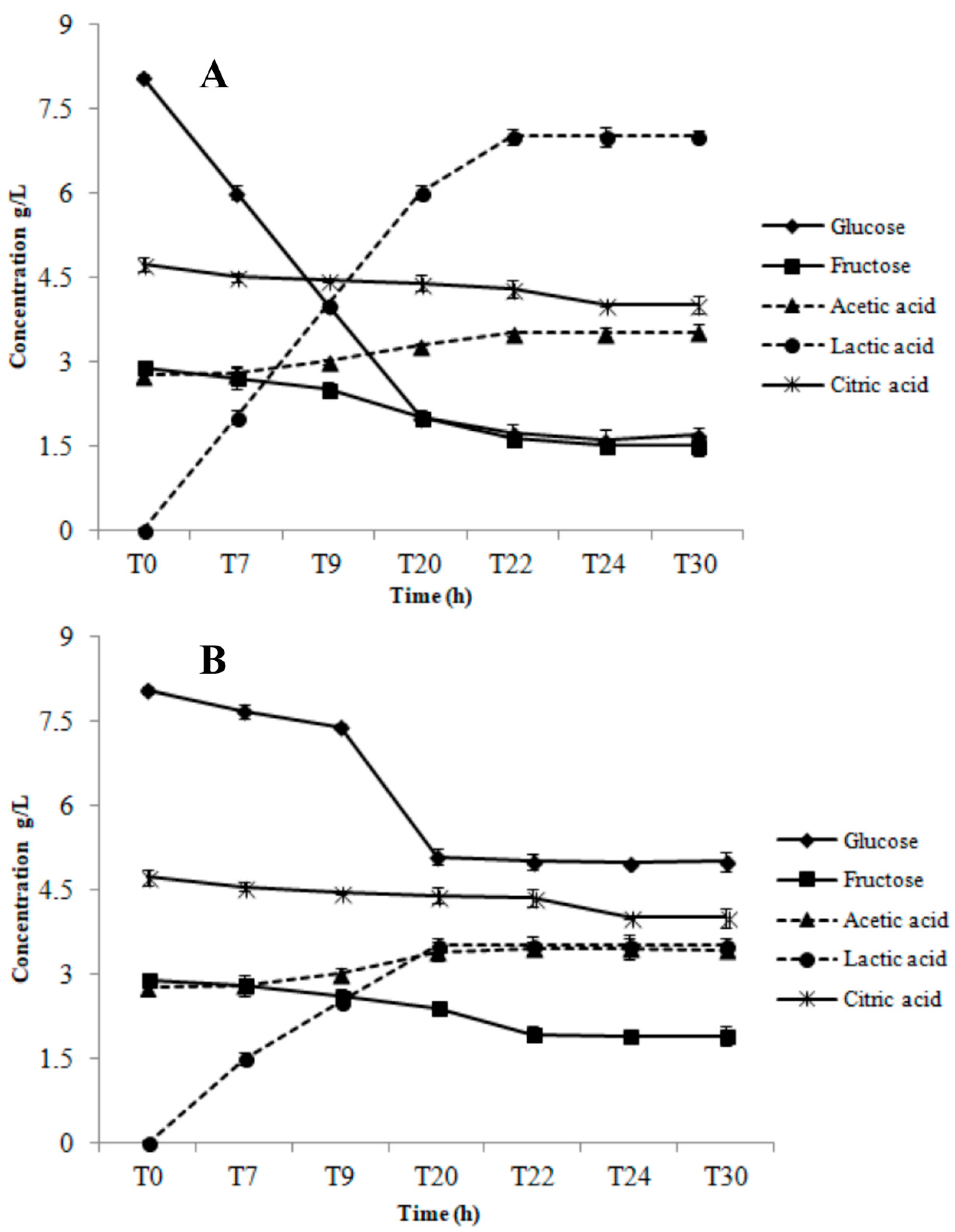
3.1. Growth Parameters and Metabolite Production
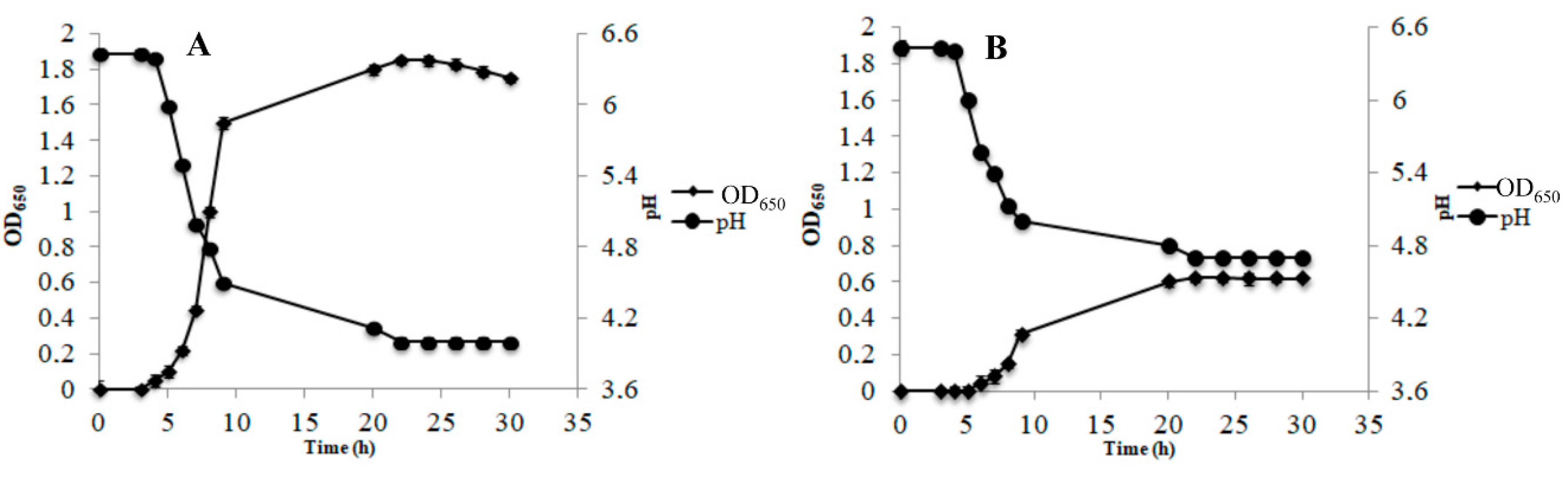
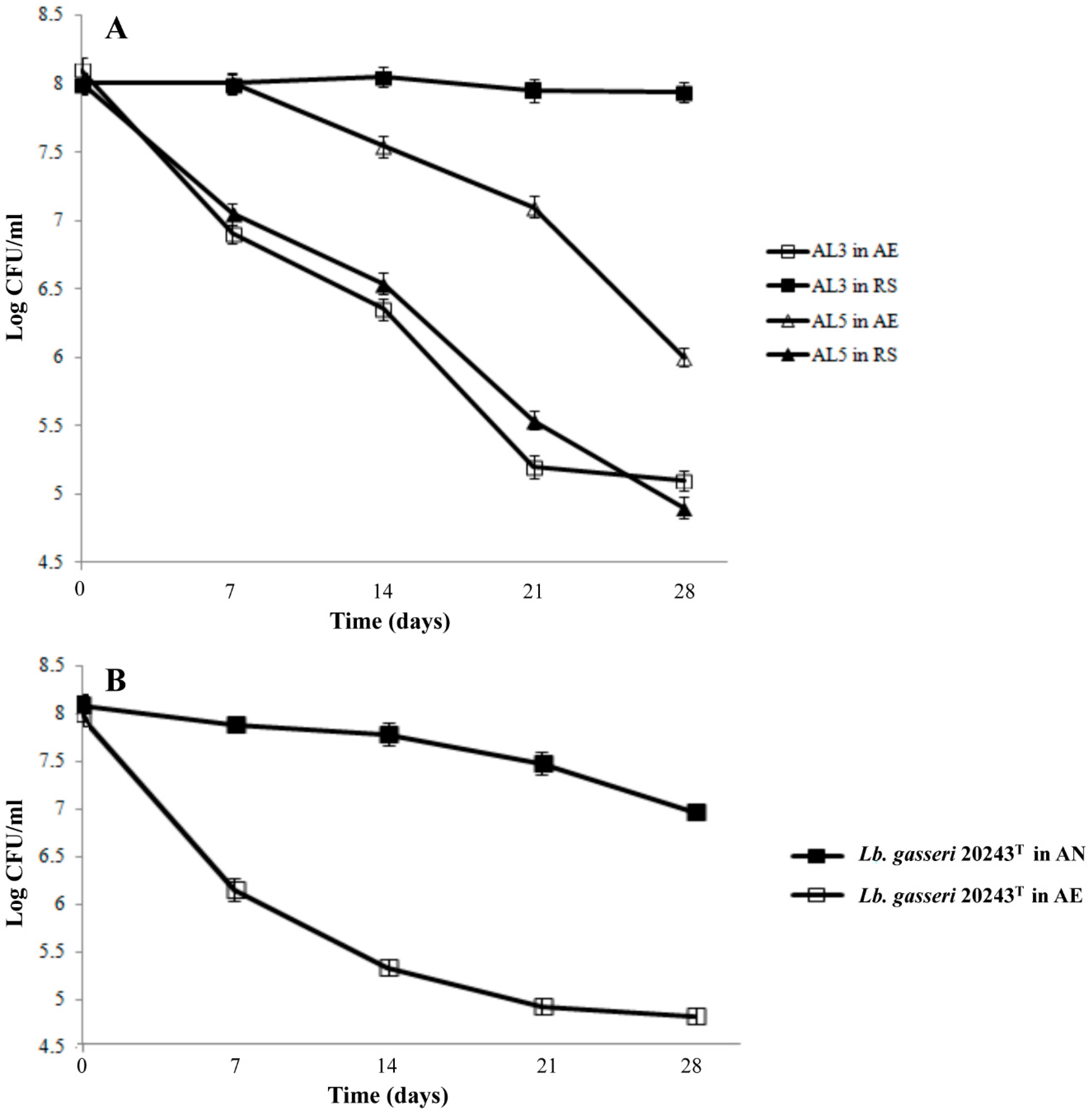
3.2. Survival of the Cells Under Cold-Starvation Stress
3.3. Comparative in Silico Analysis
4. Discussion
Author Contributions
Conflicts of Interest
References
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic metabolism in the genus Lactobacillus: Impact on stress response and potential application in the food industry. J. Appl. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Sonomoto, K.; Atsushi, Y. Oxidative stress and oxygen metabolism in lactic acid bacteria. In Lactic Acid Bacteria and Bifidobacteria; Caister Academic Press: Poole, UK, 2011. [Google Scholar]
- Pedersen, M.B.; Gaudu, P.; Lechardeur, D.; Petit, M.A.; Gruss, A. Aerobic respiration metabolism in lactic acid bacteria and uses in biotechnology. Annu. Rev. Food. Sci. Technol. 2012, 3, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Ianniello, R.G.; Zheng, J.; Zotta, T.; Ricciardi, A.; Ganzle, M.G. Biochemical analysis of respiratory metabolism in the heterofermentative Lactobacillus spicheri and Lactobacillus reuteri. J. Appl. Microbiol. 2015, 119, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Ricciardi, A.; Ianniello, R.G.; Parente, E.; Reale, A.; Rossi, F.; Iacumin, L.; Comi, G.; Coppola, R. Assessment of aerobic and respiratory growth in the Lactobacillus casei group. PLoS ONE 2014, 9, e99189. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, A.; Ianniello, R.G.; Parente, E.; Zotta, T. Modified chemically defined medium for enhanced respiratory growth of Lactobacillus casei and Lactobacillus plantarum groups. J. Appl. Microbiol. 2015, 119, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Brooijmans, J.W.; de Vos, W.M.; Hugenholtz, J. Lactobacillus plantarum electron transport chain. Appl. Environ. Microbiol. 2009, 75, 3580–3585. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Van der Veen, S.; Nakajima, H.; Abee, T. Effect of respiration and manganese on oxidative stress resistance of Lactobacillus plantarum WCFS1. Microbiology 2012, 158, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Ricciardi, A.; Guidone, A.; Sacco, M.; Muscariello, L. Inactivation of ccpA and aeration affect growth, metabolite production and stress tolerance Lactobacillus plantarum WCFS1. Int. J. Food Microbiol. 2012, 155, 51–59. [Google Scholar] [CrossRef]
- Zotta, T.; Ianniello, R.G.; Guidone, A.; Parente, E.; Ricciardi, A. Selection of mutants tolerant of oxidative stress from respiratory cultures of Lactobacillus plantarum C17. J. Appl. Microbiol. 2014, 116, 632–643. [Google Scholar] [CrossRef]
- Reale, A.; Di Renzo, T.; Zotta, T.; Preziuso, M.; Boscaino, F.; Rocco, I.; Livia, V.S.; Patrizio, T.; Raffaele, C. Effect of respirative coltures of Lactobacillus casei on model sourdough fermentation. LWT Food. Sci. Technol. 2016, 73, 622–629. [Google Scholar] [CrossRef]
- Reale, A.; Ianniello, R.G.; Ciocia, F.; di Renzo, T.; Boscaino, B.; Ricciardi, A.; Coppola, R.; Parente, E.; Zotta, T.; McSweeney, P.L.H. Effect of respirative and catalase-positive Lactobacillus casei adjuncts on the production and quality of Cheddar-type cheese. Int. Dairy. J. 2016, 63, 78–87. [Google Scholar] [CrossRef]
- Neeser, J.; Granato, D.; Rouret, M.; Servin, A.; Teneberg, S.; Karlsson, K. Lactobacillus johnsonii La1 shares carbohydrate binding specificities with several enterophatogenic bacteria. Glycobiol. 2000, 10, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Sakai, F.; Hosoya, F.; Ono-Ohmachi, A.; Ukibe, K.; Ogawa, A.; Moriya, T.; Kadooka, Y.; Shiozaki, T.; Nakagawa, H.; Nakayama, Y.; et al. Lactobacillus gasseri SBT2055 Induces TGF-b expression in dendritic cells and activates TLR2 signal to produce IgA in the small intestine. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Wood, B.J.B. The genus Lactobacillus. In Biodiversity and Taxonomy; Wiley: New York, NY, USA, 2014. [Google Scholar]
- Hertzberger, R.Y.; Pridmore, R.D.; Gysler, C.; Kleerebezem, M.; de Mattos, M. Oxygen relieves the CO2 and acetate dependency of Lactobacillus johnsonii NCC 533. PLoS ONE 2013, 8, e57235. [Google Scholar] [CrossRef] [PubMed]
- Hertzberger, R.Y.; Arents, J.; Dekker, H.; Pridmore, R.D.; Gysler, C.; Kleerebezem, M.; de Mattos, M.J. H2O2 production in species of the Lactobacillus acidophilus group: A central role for a novel NADH dependent flavin reductase. Appl. Environ. Microbiol. 2014, 80, 2229–2239. [Google Scholar] [CrossRef]
- Maresca, D.; Zotta, T.; Mauriello, G. Adaptation to aerobic environment of Lactobacillus johnsonii/gasseri strains. Front. Microbiol. 2018, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Maresca, D.; De Filippis, F.; Tytgat, H.L.P.; de Vos, W.M.; Mauriello, G. Draft genome sequences of the aerobic strains Lactobacillus gasseri AL3 and AL5. Genome Announc. 2017, 5, e00213–e00217. [Google Scholar] [CrossRef] [PubMed]
- Duwat, P.; Sourice, S.; Cesselin, B.; Lamberet, G.; Vido, K.; Gaudu, P. Respiration capacity of the fermenting bacterium Lactococcus lactis and its positive effects on growth and survival. J. Bacteriol. 2001, 183, 4509–4516. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.B.; Garrigues, C.; Tuphile, K.; Brun, C.; Vido, K.; Bennedsen, M.; Møllgaard, H.; Gaudu, P.; Gruss, A. Impact of aeration and heme-activated respiration on Lactococcus lactis gene expression: Identification of a heme-responsive operon. J. Bacteriol. 2008, 190, 4903–4911. [Google Scholar] [CrossRef] [PubMed]
- Guidone, A.; Ianniello, R.G.; Ricciardi, A.; Zotta, T.; Parente, E. Aerobic metabolism and oxidative stress tolerance in the Lactobacillus plantarum group. World J. Microbiol. Biotechnol. 2013, 29, 1713–1722. [Google Scholar] [CrossRef]
- Rezaiki, L.; Cesseli, B.; Yamamoto, Y.; Vido, K.; West, E.; Gaudu, P. Respiration metabolism reduces oxidative and acid stress to improve long-term survival of Lactococcus lactis. Mol. Microbiol. 2004, 53, 1331–1342. [Google Scholar] [CrossRef] [PubMed]
- Cesselin, B.; Derré-Bobillot, A.; Fernandez, A.; Lamberet, G.; Lachardeur, D.; Yamamoto, Y. Respiration, a strategy to avoid oxidative stress in Lactococcus lactis, is regulated by the heme status. Jpn. J. Lact. Acid Bact. 2010, 21, 10–15. [Google Scholar] [CrossRef]
- Kang, T.S.; Korber, D.R.; Tanaka, T. Influence of oxygen on NADH recycling and oxidative stress resistance systems in Lactobacillus panis PM1. AMB Express. 2013, 3, 10–13. [Google Scholar] [CrossRef]
- Mortera, P.; Pudlik, A.; Magni, C.; Alarcon, S.; Lolkema, J.S. Ca2+ Citrate uptake and metabolism in Lactobacillus casei ARCC 334. Appl. Environ. Microbiol. 2013, 79, 4603–4612. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Sivaraman, C. Bacterial citrate lyase. J. Biosci. 1984, 6, 379–401. [Google Scholar] [CrossRef]
- Kanao, T.; Fukui, T.; Atomi, H.; Imanaka, T. Kinetic and biochemical analyses on the reaction mechanism of a bacterial ATP-citrate lyase. Eur. J. Biochem. 2002, 269, 3409–3416. [Google Scholar] [CrossRef] [PubMed]
- Quintans, N.G.; Blancato, V.; Repizo, G.; Magni, C.; López1, P. Citrate metabolism and aroma compound production in lactic acid bacteria; Research Signpost: Irvine, CA, USA, 2008; ISBN 978-81-308-0250-3. [Google Scholar]
- Teysset, C.M.; de La Torre, F.; Garel, J.R. Increased production of hydrogen peroxide by Lactobacillus delbrueckii subsp. bulgaricus upon aeration: Involvement of an NADH oxidase in oxidative stress. Appl. Environ. Microbiol. 2000, 66, 262–267. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Genes | Strains | ||
|---|---|---|---|---|
| AL3 | AL5 | DSM 20243T | ||
| Aerobic metabolism | pox (pyruvate oxidase) | + | + | + |
| ack (acetate kinase) | + | + | + | |
| lox (lactate oxidase) | + | + | − | |
| nox (NADH oxidase) | + | + | − | |
| lao (L-amino acid oxidase) | + | − | − | |
| Respiratory metabolism | ndh (NADH dehydrogenase) | + | + | + |
| ubiE (ubiquinone/menaquinone biosynthesis methyltrasferase) | + | + | + | |
| cydABCD (cytochrome bd-I oxidase operon) | + | + | + | |
| Stress response | npr (NADH peroxidase) | + | + | + |
| gor (glutathione reductase) | + | + | + | |
| gop (glutathione peroxidase) | − | − | − | |
| GshA (γ-glutamylcystiene synthetase) | + | + | − | |
| GshB (glutathione synthetase) | − | − | − | |
| GshF (bifunctional glutamate-cysteine ligase/glutathione synthetase) | − | − | − | |
| TrxR (thioredoxin reductase) | + | + | + | |
| TrxP (thioredoxin peroxidase) | + | + | + | |
| SOD (superoxide dismutase) | + | + | − | |
| Kat (catalase) | − | − | − | |
| MnKat (Manganese-catalase) | − | − | − | |
| KatG (catalase-peroxidase) | − | − | − | |
| Dps (DNA binding protein from starved cells) | + | + | + | |
| Partial tricarboxylic acid (TCA) cycle | CitD (gamma subunits of citrate lyase) | + | + | − |
| CitE (beta subunits of citrate lyase) | + | + | − | |
| CitF (alpha subunits of citrate lyase) | + | + | − | |
| CitP (citrate permease) | + | + | − | |
| AOD (oxaloacetate decarboxylase) | − | − | − | |
| PYC (pyruvate carboxylase) | − | − | − | |
| MDH (malate dehydrogenase) | + | + | + | |
| FH (fumarate hydratase) | + | + | + | |
| SDH (succinate dehydrogenase) | + | + | − | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maresca, D.; De Filippis, F.; Robertiello, A.; Mauriello, G. Metabolic Profiling and Cold-Starvation Stress Response of Oxygen-Tolerant Lactobacillus gasseri Strains Cultured in Batch Bioreactor. Microorganisms 2019, 7, 200. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070200
Maresca D, De Filippis F, Robertiello A, Mauriello G. Metabolic Profiling and Cold-Starvation Stress Response of Oxygen-Tolerant Lactobacillus gasseri Strains Cultured in Batch Bioreactor. Microorganisms. 2019; 7(7):200. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070200
Chicago/Turabian StyleMaresca, Diamante, Francesca De Filippis, Alessandro Robertiello, and Gianluigi Mauriello. 2019. "Metabolic Profiling and Cold-Starvation Stress Response of Oxygen-Tolerant Lactobacillus gasseri Strains Cultured in Batch Bioreactor" Microorganisms 7, no. 7: 200. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070200