Engineering Synthetic Microbial Communities through a Selective Biofilm Cultivation Device for the Production of Fermented Beverages

 and
and
Abstract
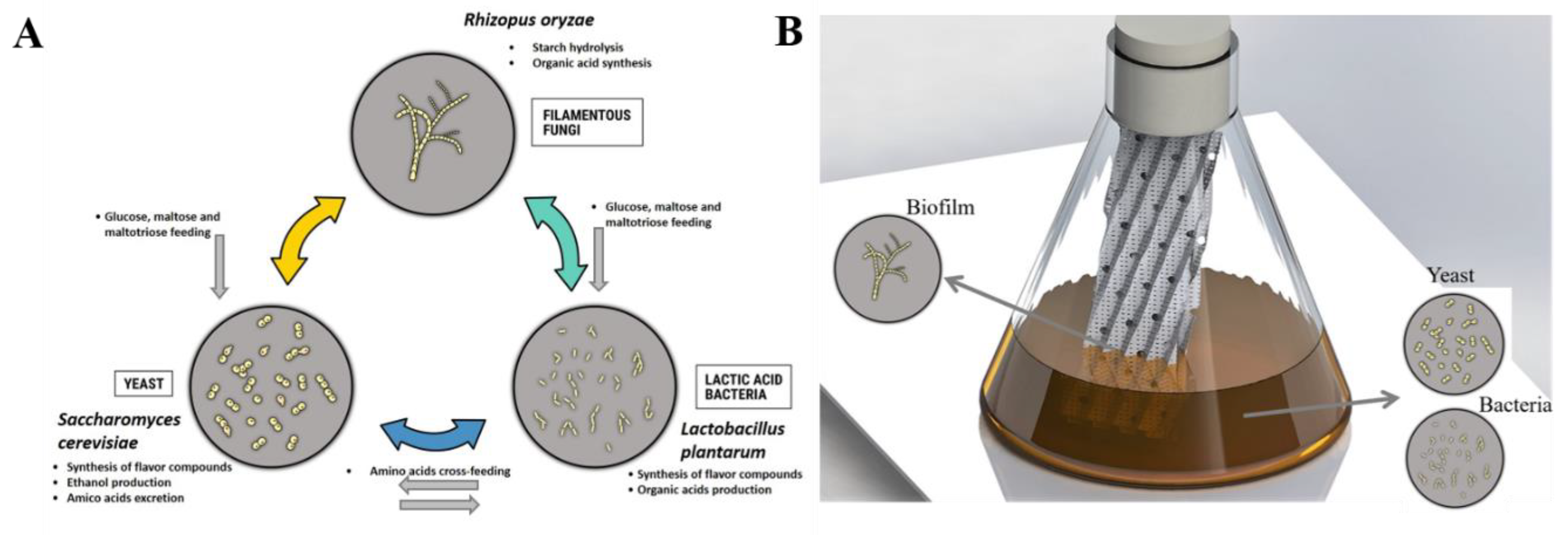
:1. Introduction
2. Materials and Methods
2.1. Strain and Medium Preparation
2.2. Fermentation Based on Red Rice
2.3. Fermentation Based on Synthetic Liquid Medium
2.4. Aromatic Compounds Analysis byHS-SPME-GC-MS
2.5. Sugar and Ethanol Analysis by HPLC
2.6. Microbiological Analysis
2.7. Statistical Aanalysis
3. Results
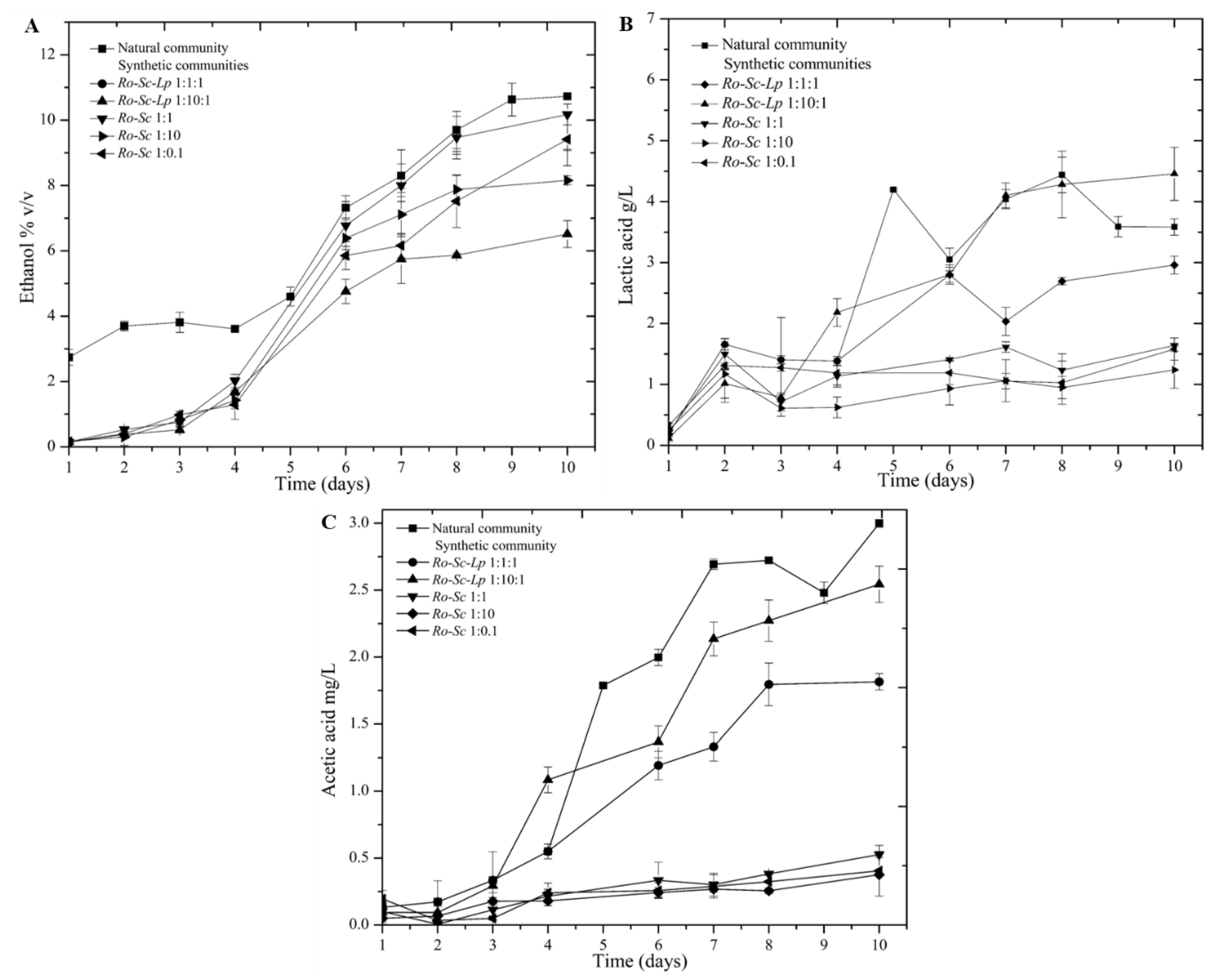
3.1. Comparison of Natural and Synthetic Community for Red Rice Wine Fermentation
3.1.1. Ethanol and Organic Acid Production during Red Rice Wine Processing
3.1.2. Comparative Analysis of the Impact of Natural and Synthetic Communities on the Flavor Profile during Red Rice Wine Fermentation
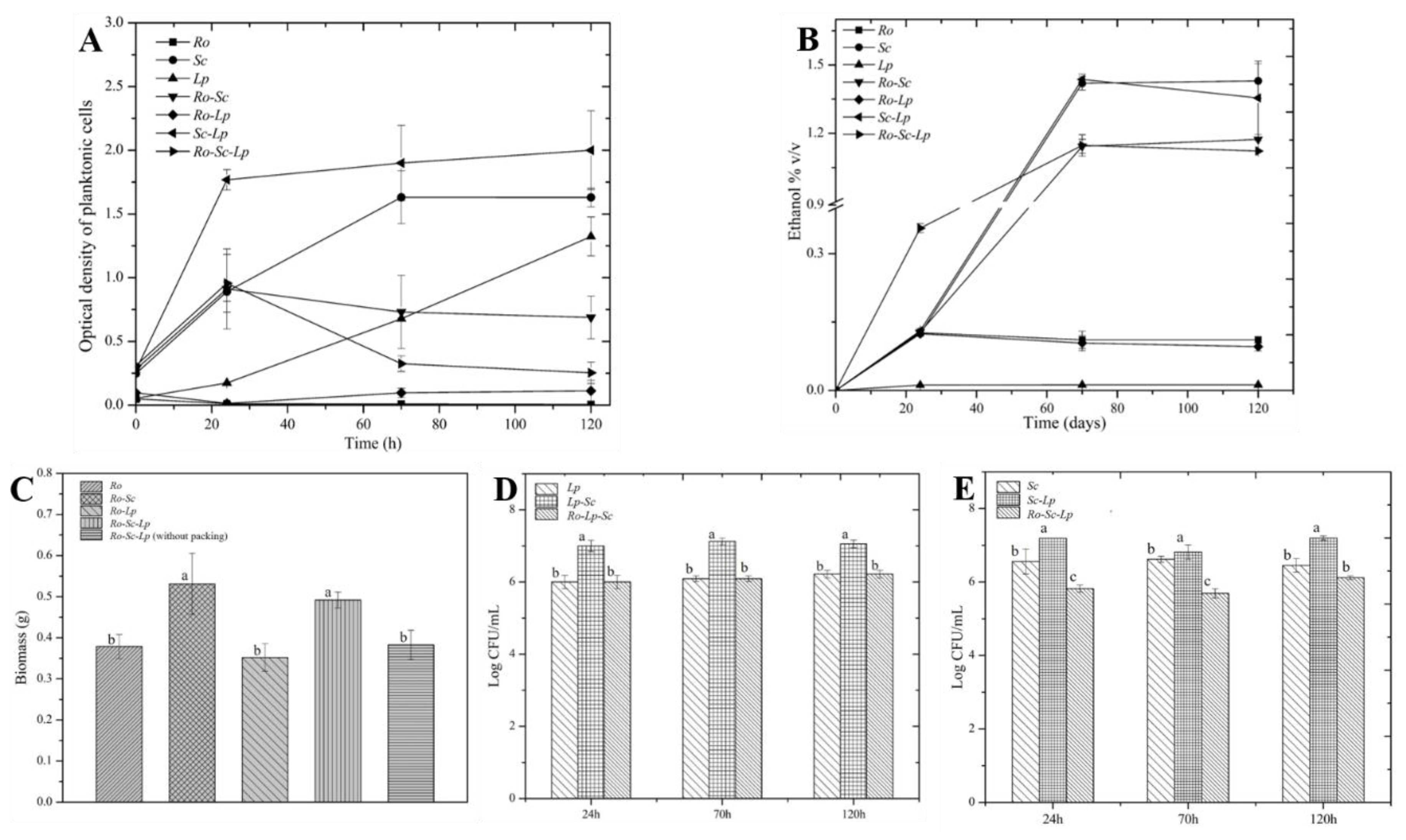
3.2. Spatial Structuration of Synthetic Communities Based on Biofilm Ccultivation.
3.2.1. Impact of Submerged and Biofilm Cultivation on Colonization Efficiency
3.2.2. Impact of Biofilm Mode of Cultivation on Flavor Compound Using Natural and Synthetic Communities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Uzogara, S.G.; Agu, L.N.; Uzogara, E.O. A review of traditional fermented foods, condiments and beverages in nigeria: Their benefits and possible problems. Ecol. Food Nutr. 1990. [Google Scholar] [CrossRef]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Sujaya, I.N.; Antara, N.S.; Sone, T.; Tamura, Y.; Aryanta, W.R.; Yokota, a.; Asano, K.; Tomita, F. Identification and characterization of yeasts in brem, a traditional Balinese rice wine. World J. Microbiol. Biotechnol. 2004, 20, 143–150. [Google Scholar] [CrossRef]
- Dung, N.T.P.; Rombouts, F.M.; Nout, M.J.R. Characteristics of some traditional Vietnamese starch-based rice wine fermentation starters (men). LWT - Food Sci. Technol. 2007, 40, 130–135. [Google Scholar] [CrossRef]
- Ercolini, D. PCR-DGGE fingerprinting: novel strategies for detection of microbes in food. J. Microbiol. Methods 2004, 56, 297–314. [Google Scholar] [CrossRef]
- Jeyaram, K.; Singh, W.; Capece, A.; Romano, P. Molecular identification of yeast species associated with ‘Hamei’ — A traditional starter used for rice wine production in Manipur, India. Int. J. Food Microbiol. 2008, 124, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Thanh, V.N.; Mai, L.T.; Tuan, D.A. Microbial diversity of traditional Vietnamese alcohol fermentation starters (banh men) as determined by PCR-mediated DGGE. Int. J. Food Microbiol. 2008, 128, 268–273. [Google Scholar] [CrossRef]
- Lv, X.-C.; Weng, X.; Zhang, W.; Rao, P.-F.; Ni, L. Microbial diversity of traditional fermentation starters for Hong Qu glutinous rice wine as determined by PCR-mediated DGGE. Food Control 2012, 28, 426–434. [Google Scholar] [CrossRef]
- Lv, X.-C.C.; Cai, Q.-Q.Q.; Ke, X.-X.X.; Chen, F.; Rao, P.-F.F.; Ni, L. Characterization of fungal community and dynamics during the traditional brewing of Wuyi Hong Qu glutinous rice wine by means of multiple culture-independent methods. Food Control 2015, 54, 231–239. [Google Scholar] [CrossRef]
- Chao, S.H.; Huang, H.Y.; Kang, Y.H.; Watanabe, K.; Tsai, Y.C. The diversity of lactic acid bacteria in a traditional Taiwanese millet alcoholic beverage during fermentation. LWT - Food Sci. Technol. 2013, 51, 135–142. [Google Scholar] [CrossRef]
- Luangkhlaypho, A.; Pattaragulwanit, K.; Leepipatpiboon, N.; Yompakdee, C. Development of a defined starter culture mixture for the fermentation of sato, a Thai rice-based alcoholic beverage. ScienceAsia 2014, 40, 125–134. [Google Scholar] [CrossRef]
- Wang, P.-P.; Li, Z.; Qi, T.-T.; Li, X.-J.; Pan, S.-Y. Development of a method for identification and accurate quantitation of aroma compounds in Chinese Daohuaxiang liquors based on SPME using a sol–gel fibre. Food Chem. 2015, 169, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Ly, S.; Mith, H.; Tarayre, C.; Taminiau, B.; Daube, G.; Fauconnier, M.; Delvigne, F. Impact of Microbial Composition of Cambodian Traditional Dried Starters (Dombea) on Flavor Compounds of Rice Wine: Combining Amplicon Sequencing With HP-SPME-GCMS. Front. Microbiol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rousseaux, S.; Tourdot-Maréchal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine microbiome: A dynamic world of microbial interactions. Crit. Rev. Food Sci. Nutr. 2017, 57, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacón, J.J.; Ballester, J.; Vichi, S.; Guérin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnol. Adv. 2004, 22, 189–259. [Google Scholar] [CrossRef] [PubMed]
- Zune, Q.; Delepierre, A.; Gofflot, S.; Bauwens, J.; Twizere, J.C.; Punt, P.J.; Francis, F.; Toye, D.; Bawin, T.; Delvigne, F. A fungal biofilm reactor based on metal structured packing improves the quality of a Gla::GFP fusion protein produced by Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2015, 99, 6241–6254. [Google Scholar] [CrossRef] [Green Version]
- Klitgord, N.; Segrè, D. Environments that Induce Synthetic Microbial Ecosystems. PLoS Comput. Biol. 2010, 6, e1001002. [Google Scholar] [CrossRef]
- Hays, S.G.; Patrick, W.G.; Ziesack, M.; Oxman, N.; Silver, P.A. Better together: engineering and application of microbial symbioses. Curr. Opin. Biotechnol. 2015, 36, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Khalesi, M.; Zune, Q.; Telek, S.; Riveros-Galan, D.; Verachtert, H.; Toye, D.; Gebruers, K.; Derdelinckx, G.; Delvigne, F. Fungal biofilm reactor improves the productivity of hydrophobin HFBII. Biochem. Eng. J. 2014, 88, 171–178. [Google Scholar] [CrossRef]
- Vandermies, M.; Kar, T.; Carly, F.; Nicaud, J.-M.; Delvigne, F.; Fickers, P. Yarrowia lipolytica morphological mutant enables lasting in situ immobilization in bioreactor. Appl. Microbiol. Biotechnol. 2018, 102, 5473–5482. [Google Scholar] [CrossRef] [PubMed]
- Zune, Q.; Soyeurt, D.; Toye, D.; Ongena, M.; Thonart, P.; Delvigne, F. High-energy X-ray tomography analysis of a metal packing biofilm reactor for the production of lipopeptides by\n Bacillus subtilis. J. Chem. Technol. Biotechnol. 2014, 89, 382–390. [Google Scholar] [CrossRef]
- Zune, Q.; Telek, S.; Calvo, S.; Salmon, T.; Alchihab, M.; Toye, D.; Delvigne, F. Influence of liquid phase hydrodynamics on biofilm formation on structured packing: Optimisation of surfactin production from Bacillus amyloliquefaciens. Chem. Eng. Sci. 2016. [Google Scholar] [CrossRef]
- Sompong, R.; Siebenhandl-Ehn, S.; Linsberger-Martin, G.; Berghofer, E. Physicochemical and antioxidative properties of red and black rice varieties from Thailand, China and Sri Lanka. Food Chem. 2011, 124, 132–140. [Google Scholar] [CrossRef]
- Zune, Q.; Toye, D. Implementation of a Metal Structured Packing in a Fungal Biofilm Reactor for the Production of a Recombinant Protein By. Comm. Appl. Biol. Sci. 2013, 1–6. [Google Scholar]
- Yang, D.; Luo, X.; Wang, X. Characteristics of traditional Chinese shanlan wine fermentation. J. Biosci. Bioeng. 2014, 117, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Baumes, R.; Cordonnier, R.; Nitz, S.; Drawert, F. Identification and determination of volatile constituents in wines from different vine cultivars. J. Sci. Food Agric. 1986. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Vallés, S.; Manzanares, P. International Journal of Food Microbiology Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2009, 135, 68–74. [Google Scholar] [CrossRef]
- Lindemann, S.R.; Bernstein, H.C.; Song, H.S.; Fredrickson, J.K.; Fields, M.W.; Shou, W.; Johnson, D.R.; Beliaev, A.S. Engineering microbial consortia for controllable outputs. ISME J. 2016. [Google Scholar] [CrossRef]
- Xie, G.-F.; Li, W.-J.; Lu, J.; Cao, Y.; Fang, H.; Zou, H.-J.; Hu, Z.-M. Isolation and identification of representative fungi from shaoxing rice wine wheat Qu using a polyphasic approach of culture-based and molecular-based methods. J. Inst. Brew. 2007. [Google Scholar] [CrossRef]
- O’Brien, S.; Wang, Y.J. Susceptibility of annealed starches to hydrolysis by??-amylase and glucoamylase. Carbohydr. Polym. 2008, 72, 597–607. [Google Scholar] [CrossRef]
- Mukisa, I.M.; Byaruhanga, Y.B.; Muyanja, C.M.B.K.; Langsrud, T.; Narvhus, J.A. Production of organic flavor compounds by dominant lactic acid bacteria and yeasts from Obushera, a traditional sorghum malt fermented beverage. Food Sci. Nutr. 2017, 5, 702–712. [Google Scholar] [CrossRef]
- Ponomarova, O.; Gabrielli, N.; Sévin, D.C.; Mülleder, M.; Zirngibl, K.; Bulyha, K.; Andrejev, S.; Kafkia, E.; Typas, A.; Sauer, U.; et al. Yeast Creates a Niche for Symbiotic Lactic Acid Bacteria through Nitrogen Overflow. Cell Syst. 2017, 5, 345–357.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nout, M.J.R.; Sarkar, P.K. Lactic acid food fermentation in tropical climates. Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 1999, 76, 395–401. [Google Scholar] [CrossRef]
- Killian, E.; Ough, C.S. Fermentation Esters — Formation and Retention as Affected by Fermentation Temperature. Am. J. Enol. Vitic. 1979. [Google Scholar]
- Guitart, A.; Hernández Orte, P.; Ferreira, V.; Peña, C.; Cacho, J. Some observations about the correlation between the amino acid content of musts and wines of the Chardonnay variety and their fermentation aromas. Am. J. Enol. Vitic. 1999. [Google Scholar]
- Zaupa, M.; Calani, L.; Del Rio, D.; Brighenti, F.; Pellegrini, N. Characterization of total antioxidant capacity and (poly)phenolic compounds of differently pigmented rice varieties and their changes during domestic cooking. Food Chem. 2015, 187, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012. [Google Scholar] [CrossRef]
- Rosales, E.; Pazos, M.; Ángeles Sanromán, M. Solid-State Fermentation for Food Applications. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherland, 2018; pp. 319–355. ISBN 9780444639905. [Google Scholar]
- Dung, N.T.P.; Rombouts, F.M.; Nout, M.J.R. Development of defined mixed-culture fungal fermentation starter granulate for controlled production of rice wine. Innov. Food Sci. Emerg. Technol. 2005, 6, 429–441. [Google Scholar] [CrossRef]
- Scherlach, K.; Graupner, K.; Hertweck, C. Molecular Bacteria-Fungi Interactions: Effects on Environment, Food, and Medicine. Annu. Rev. Microbiol. 2013, 67, 375–397. [Google Scholar] [CrossRef]
- Braga, R.M.; Dourado, M.N.; Araújo, W.L. Microbial interactions: ecology in a molecular perspective. Brazilian J. Microbiol. 2016, 47, 86–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolinšek, J.; Goldschmidt, F.; Johnson, D.R. Synthetic microbial ecology and the dynamic interplay between microbial genotypes. FEMS Microbiol. Rev. 2016, 40, 961–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Røder, H.L.; Madsen, J.S.; Bjarnsholt, T.; Sørensen, S.J.; Burmølle, M. Interspecific bacterial interactions are reflected in multispecies biofilm spatial organization. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ponomarova, O.; Patil, K.R. Metabolic interactions in microbial communities: Untangling the Gordian knot. Curr. Opin. Microbiol. 2015, 27, 37–44. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration | Natural Community | Ro-Sc-Lp 1:1:1 | Ro-Sc-Lp 1:10:1 | Ro-Sc 1:1 | Ro-Sc 1:10 | Ro-Sc 1:0.1 |
|---|---|---|---|---|---|---|
| Ethanol (% v/v) | 10.73 ± 0.07 a | 9.95 ± 0.02 b | 6.5 ± 0.41 d | 10.17 ± 0.32 b | 7.95 ± 0.13 c | 8.41 ± 0.80 c |
| Acetic acid (g/L) | 2.99 ± 0.02 a | 1.81 ± 0.06 c | 2.54 ± 0.13 b | 0.52 ± 0.02 d | 0.37 ± 0.01 d | 0.40 ± 0.18 d |
| Lactic acid (g/L) | 3.58 ± 0.13 a | 2.95 ± 0.14 b | 4.45 ± 0.43 a | 1.64 ± 0.01 c | 1.23 ± 0.30 c | 1.58 ± 0.20 c |
| N | Compounds | RI | Natural Community | Synthetic Community | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ro-Sc-Lp 1:1:1 | Ro-Sc-Lp 1:10:1 | Ro-Sc 1:1 | Ro-Sc 1:10 | Ro-Sc 1:0.1 | ||||||||||||||
| Average | STD | Average | STD | Average | STD | Average | STD | Average | STD | Average | STD | |||||||
| Ester | 19506.02 | 24029.27 | 21738.23 | 28584.87 | 33152.54 | 19290.85 | ||||||||||||
| 1 | 2-Methybutyl acetate | 880 | Nd | 137.89 | 15.85 | Nd | 55.07 | 21.45 | 170.05 | 4.09 | Nd | |||||||
| 2 | Isoamyl acetate | 876 | 125.98 | 15.4 | 270.1 | 107.15 | 493.09 | 123.93 | 155.8 | 9.3 | 533.14 | 4.66 | 344.24 | 193.83 | ||||
| 3 | 1-Methoxy-2-propyl acetate | Nd | Nd | Nd | Nd | 157.95 | 33.39 | 18.83 | ||||||||||
| 4 | Ethyl linoleate | 60.44 | 36.9 | 41.39 | 9.75 | Nd | 106.44 | 213.92 | 195.68 | Nd | ||||||||
| 5 | 4-Methylbenzyl alcohol | Nd | Nd | Nd | 30.7 | 35.16 | 35.42 | |||||||||||
| 6 | Phenylethyl acetate | 1252 | 1383 | 1421.55 | 346.22 | 1718.85 | 40.95 | 427.9 | 371.34 | 683.62 | 71.9 | 98.05 | 62.35 | |||||
| 7 | Ethyl isobutyrate | 756 | Nd | 275.28 | 61.82 | Nd | 270.53 | 149.22 | 296.72 | 74.47 | 381.78 | 175.2 | ||||||
| 8 | Ethyl propionate | 713 | Nd | 131.79 | 55.24 | 172.86 | 100.09 | 186.71 | 63.47 | 416.39 | 294.43 | 101.98 | 10.34 | |||||
| 9 | Ethyl myristate | 1651 | 1206.16 | 63.67 | 1202.58 | 70.77 | 1296.14 | 123.41 | 80.21 | 113.44 | 216.05 | 56.89 | 61.84 | 31.72 | ||||
| 10 | Ethyl octanoate | 1041 | 294.15 | 5.68 | 134.73 | 18.25 | 154.17 | 7.44 | 38.33 | 54.21 | 41.68 | 1.137 | 20.75 | |||||
| 11 | Isobutyl lactate | 51.88 | 48.02 | 43.82 | 17.34 | 266.86 | 14.88 | 8.96 | 12.68 | 29.6 | Nd | |||||||
| 12 | Ethyl lactate | 1010 | 4436.35 | 1501 | 3781.81 | 1016.62 | 3941.33 | 635.91 | 941.16 | 479.84 | 2313.94 | 1345.6 | 1137.4 | |||||
| 13 | Propyl 9,12-octadecadienoate | Nd | 17.65 | 7.71 | 59.41 | 15.7 | Nd | Nd | 20.92 | 6.88 | ||||||||
| 14 | Ethyl stearate | 253.6 | 154.03 | 121.71 | 37.42 | 366.97 | 1.31 | 234.15 | 70.35 | 179.59 | 68.49 | 20.74 | ||||||
| 15 | Ethyl decanoate | 1398 | 63 | 1.42 | 65.25 | 7.81 | 269.24 | 204.4 | 160.2 | 106.35 | 154.6 | 3.35 | 90.31 | 18.23 | ||||
| 16 | Ethyl laurate | 1494 | 281.26 | 53.27 | 213.47 | 23.88 | 412.84 | 1.29 | 241 | 144.21 | 109.99 | 1.69 | 44.13 | 18.64 | ||||
| 17 | Ethyl 9-hexadecenoate | Nd | Nd | Nd | 122.79 | 64.19 | 61.07 | 86.37 | 24.47 | 12.34 | ||||||||
| 18 | Ethyl Acetate | 628 | 9205.5 | 1011.45 | 14189.71 | 4590.13 | 4593.61 | 391.19 | 20976.76 | 8697.86 | 24587.57 | 7650.87 | 15713.3 | 0 | ||||
| 19 | Ethyl Oleate | 189.26 | 58.42 | 196.77 | 86.69 | 148.4 | 28.87 | 594.82 | 164.86 | 669.91 | 776.57 | 103.1 | 53.28 | |||||
| 20 | Ethyl palmitate | 1765.79 | 162.58 | 1467.66 | 213.54 | 7218.18 | 883.39 | 3420.41 | 1023.08 | 1901.87 | 129.36 | 537.84 | 343.77 | |||||
| 21 | Isoamyl lactate | 149.34 | 14.15 | 34.34 | 14.56 | 134.23 | 15.56 | Nd | 32.67 | 23.1 | Nd | |||||||
| 22 | Diethyl succinate | 1167 | Nd | 5.43 | 1.6 | Nd | 28.61 | 14.22 | 22.1 | 0.78 | 12.27 | |||||||
| 23 | Isobutyl acetate | 776 | Nd | 214.98 | 105.78 | 388.61 | 13.36 | 87.83 | 2.41 | 242.27 | 38.11 | 57.66 | 45.7 | |||||
| Acids and Aldehyde | 7525.45 | 5551.64 | 7215.71 | 1274.4 | 8219.13 | 588.64 | ||||||||||||
| 24 | 2-Methylbutanoic acid | 254.71 | 28.66 | 158.23 | 56.37 | Nd | 204.84 | 76.25 | 352.89 | 67.68 | 79.54 | 82.67 | ||||||
| 25 | Isovaleric acid | 877 | 157.19 | 47.88 | 108.41 | 18.89 | Nd | 125.98 | 15.88 | 22.24 | 22.19 | 15.69 | ||||||
| 26 | Acetic acid | 600 | 6050.36 | 225.56 | 4602.05 | 75.1 | 6487.79 | 1225.43 | 85.13 | 111.22 | 7576.43 | 524.84 | Nd | |||||
| 27 | Isobutyric acid | 1215 | 677.61 | 82.74 | 536.91 | 83.6 | 632.95 | 16.75 | 420.74 | 205.59 | 75.61 | 120.04 | 138.59 | |||||
| 28 | Acetaldehyde | Nd | 6.07 | 5.5 | 39.12 | 1.15 | 387.05 | Nd | 366.87 | 219.63 | ||||||||
| 29 | Benzaldehyde | 960 | 134.63 | 14.02 | 40.06 | 7.93 | 22.17 | 14.06 | 16.61 | 23.25 | Nd | |||||||
| 30 | Benzeneacetaldehyde | 250.92 | 23.21 | 105.95 | 38.56 | 72.78 | 8.01 | 34.05 | 6.04 | 38.72 | 6.18 | Nd | ||||||
| Alcohol | 46685.3 | 47489.59 | 47434.43 | 56613.62 | 68742.36 | 51864.3 | ||||||||||||
| 31 | 1-Hexanol | 851 | 108.9 | 10.82 | 137.59 | 4.57 | 33.29 | 1.49 | 62.15 | 7.74 | 14.8 | 7.34 | 33.65 | 47.6 | ||||
| 32 | 3-ethoxy-Propanol | 833 | 40.23 | 2.67 | 55.19 | 34.44 | 64.25 | 17.715 | 29.33 | 7.05 | 82.57 | 12.27 | 98.85 | 52.61 | ||||
| 33 | Isobutanol | 647 | 9854.79 | 262.93 | 8041.47 | 2.28 | 22432.98 | 1803.43 | 17018.02 | 12945.72 | 2849.94 | 12593.74 | 1040.56 | |||||
| 34 | Methionol | 978 | 266.39 | 142.01 | 266.71 | 30.07 | Nd | 42.58 | 64.56 | 5.93 | 8.4 | |||||||
| 35 | Isoamyl alcohol | 31276.83 | 2546.51 | 30257.95 | 536.34 | 21182.44 | 1471.7 | 34607.6 | 11519.76 | 34191.78 | 4674.47 | 24232.3 | 12570.1 | |||||
| 36 | Guaiacol | 1089 | 131.53 | 4.58 | 85.42 | 13.26 | 93.52 | 12.09 | 115.24 | 44.51 | 125.01 | 28.39 | 49.07 | |||||
| 37 | Phenylethyl Alcohol | 1118 | 3294.85 | 534.53 | 3446.39 | 576.43 | 8502.17 | 1331.22 | 9527.54 | 2295.83 | 9903.82 | 4338.51 | 6696.66 | 3835.75 | ||||
| 38 | 1-Butanol | 675 | 167.66 | 28.22 | 530.19 | 92.92 | 343.05 | 311.04 | 294.54 | 81.73 | 895.69 | 507.3 | 143.73 | |||||
| 39 | 2,3-Butanediol | 806 | 1584.31 | 460.75 | 4668.68 | 736.44 | 4026.13 | 744.14 | 3648.25 | 755.42 | 10600.96 | 6533.65 | 7745.62 | |||||
| Total | 73716.77 | 77015.35 | 85567.57 | 94788.2 | 110114.03 | 71376.9 | ||||||||||||
| Compounds | Natural Community | Synthetic Community Ro-Sc-Lp | ||
|---|---|---|---|---|
| With Packing | Without Packing | With Packing | Without Packing | |
| Isoamyl acetate | 1.78 ± 0.3 | 2.00 ± 0.46 | Nd | Nd |
| Phenethyl acetate | 1159.08 ± 69.74 | 7.92 ± 0.95 | Nd | Nd |
| Ethyl propionate | 4.85 ± 0.15 | Nd | Nd | Nd |
| Ethyl myristate | 3.29 ± 0.1 | Nd | Nd | Nd |
| Ethyl stearate | 3.03 ± 0.8 | Nd | Nd | Nd |
| Ethyl laurate | Nd | 26.58 ± 16.80 | Nd | Nd |
| Ethyl Acetate | 3.16 ± 1.19 | 1.65 ± 0.12 | Nd | Nd |
| Ethyl lactate | Nd | Nd | 2.5 ± 0.2 | Nd |
| Ethyl Oleate | 9.12 ± 0.98 | 2.97±1.61 | Nd | Nd |
| Ethyl Isobutyrate | Nd | Nd | 16.62 ± 0.89 | Nd |
| Ethyl palmitate | 28.33 ± 17.45 | 3.64 ± 1.07 | Nd | Nd |
| Isobutyl acetate | 43.14 ± 6.43 | 34.01 ± 14.05 | Nd | Nd |
| Isovaleric acid | Nd | 23.69 ± 6.19 | Nd | 275.58 ± 45.67 |
| Pentanoic acid | Nd | Nd | 124.72 ± 13.45 | 593.58 ± 34.23 |
| Acetic acid | 23.31 ± 2.63 | 8.53 ± 6.03 | 4.13 ± 0.56 | 1017.38 ± 21.12 |
| 1-Hexanol | 40.66 ± 6.89 | Nd | Nd | 18.82 ± 0.93 |
| Isobutanol | 3.24 ± 0.3 | 210.87 ± 140.32 | Nd | Nd |
| Methionol | Nd | 4.35 ± 2.75 | Nd | Nd |
| Propanol | Nd | Nd | 10.41 ± 2.34 | Nd |
| Butanol | Nd | Nd | 10.72 ± 3.23 | Nd |
| 1-Methyl-butanol | Nd | Nd | 5.04 ± 0.12 | Nd |
| 2-methyl-butanol | Nd | Nd | 158.59 ± 13.89 | Nd |
| Isobutyl alcohol | Nd | Nd | 378.19 ± 16.78 | Nd |
| Isoamyl alcohol | 2374.62 ± 64.32 | 1703.32 ± 19.8 | 4247.63 ± 234.89 | 256.69 ± 13.89 |
| Guaiacol | 3.29 ± 0.5 | Nd | Nd | 3594.66 ± 84.91 |
| Phenylethyl Alcohol | 1902.86 ± 89.90 | 0.58 ± 0.41 | 2017.87 ± 89.78 | Nd |
| 2,3-Butanediol | 47.80 ± 1.21 | Nd | Nd | 2070.68 ± 34.37 |
| Benzaldehyde | 25.50 ± 5.25 | 136.39 ± 17.50 | 91.75 ± 7.62 | Nd |
| Acetaldehyde | 2.78 ± 0.67 | Nd | Nd | 559.68 ± 23.34 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ly, S.; Bajoul Kakahi, F.; Mith, H.; Phat, C.; Fifani, B.; Kenne, T.; Fauconnier, M.-L.; Delvigne, F. Engineering Synthetic Microbial Communities through a Selective Biofilm Cultivation Device for the Production of Fermented Beverages. Microorganisms 2019, 7, 206. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070206
Ly S, Bajoul Kakahi F, Mith H, Phat C, Fifani B, Kenne T, Fauconnier M-L, Delvigne F. Engineering Synthetic Microbial Communities through a Selective Biofilm Cultivation Device for the Production of Fermented Beverages. Microorganisms. 2019; 7(7):206. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070206
Chicago/Turabian StyleLy, Sokny, F. Bajoul Kakahi, Hasika Mith, Chanvorleak Phat, Barbara Fifani, Tierry Kenne, Marie-Laure Fauconnier, and Frank Delvigne. 2019. "Engineering Synthetic Microbial Communities through a Selective Biofilm Cultivation Device for the Production of Fermented Beverages" Microorganisms 7, no. 7: 206. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070206