Upgrading Grape Pomace through Pleurotus spp. Cultivation for the Production of Enzymes and Fruiting Bodies

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Inoculum Preparation
2.2. Raw Materials and Fermentation Media
2.3. SSF, SmF and SLF Conditions
2.4. Cultivation Conditions for Fruiting Bodies Production
2.5. Analytical Methods
2.5.1. Hydrolysis of Grape Pomace and Biomass Production
2.5.2. Determination of Glucosamine Content
2.5.3. Indirect Estimation of Biomass in Fermentations
2.5.4. Crude enzyme extraction and determination of enzyme activities
2.5.5. Determination of Phenolic Compounds
2.6. Statistical Analysis
3. Results and Discussion
3.1. Biomass Production
3.2. Consumption of Phenolic Compounds
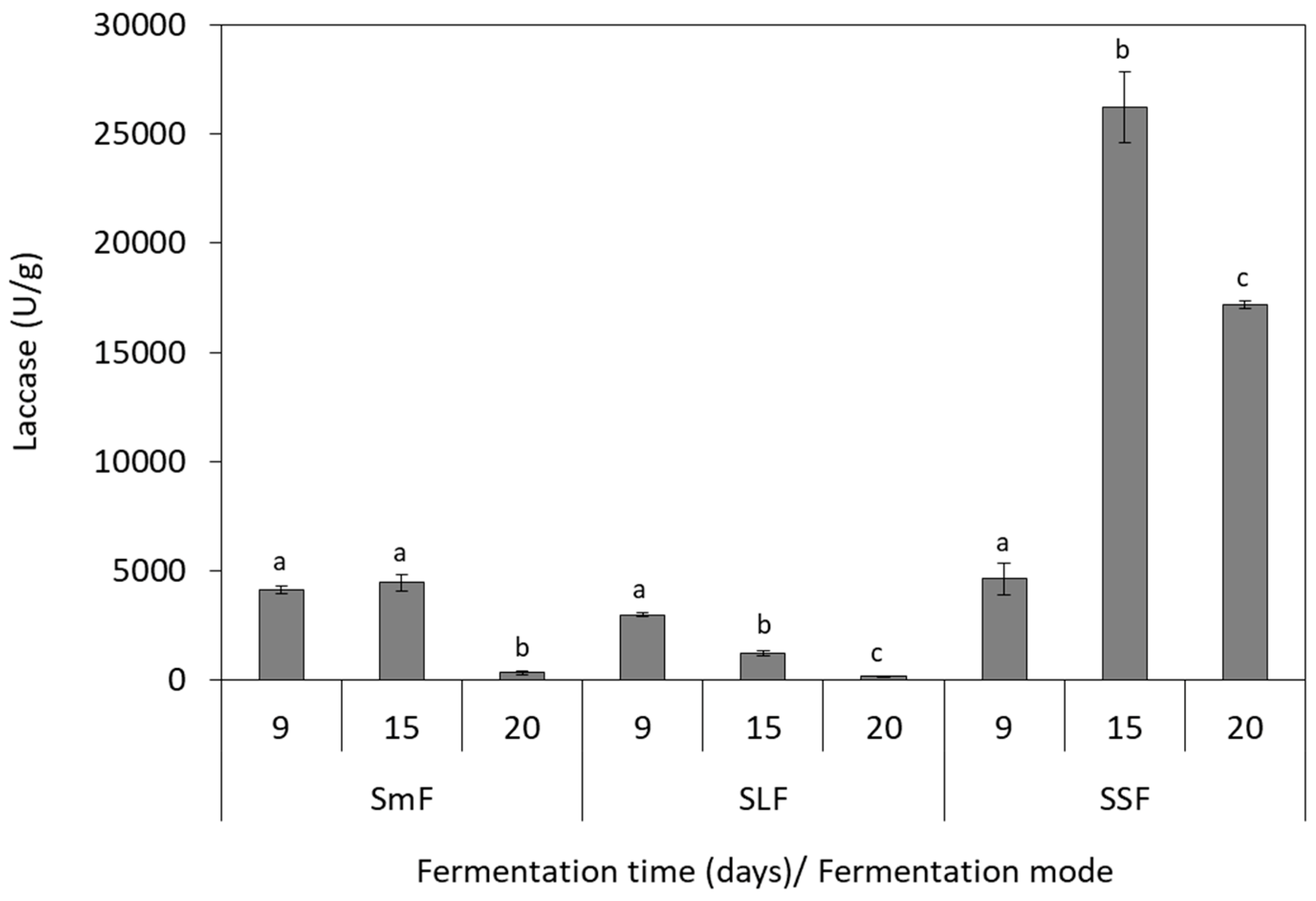
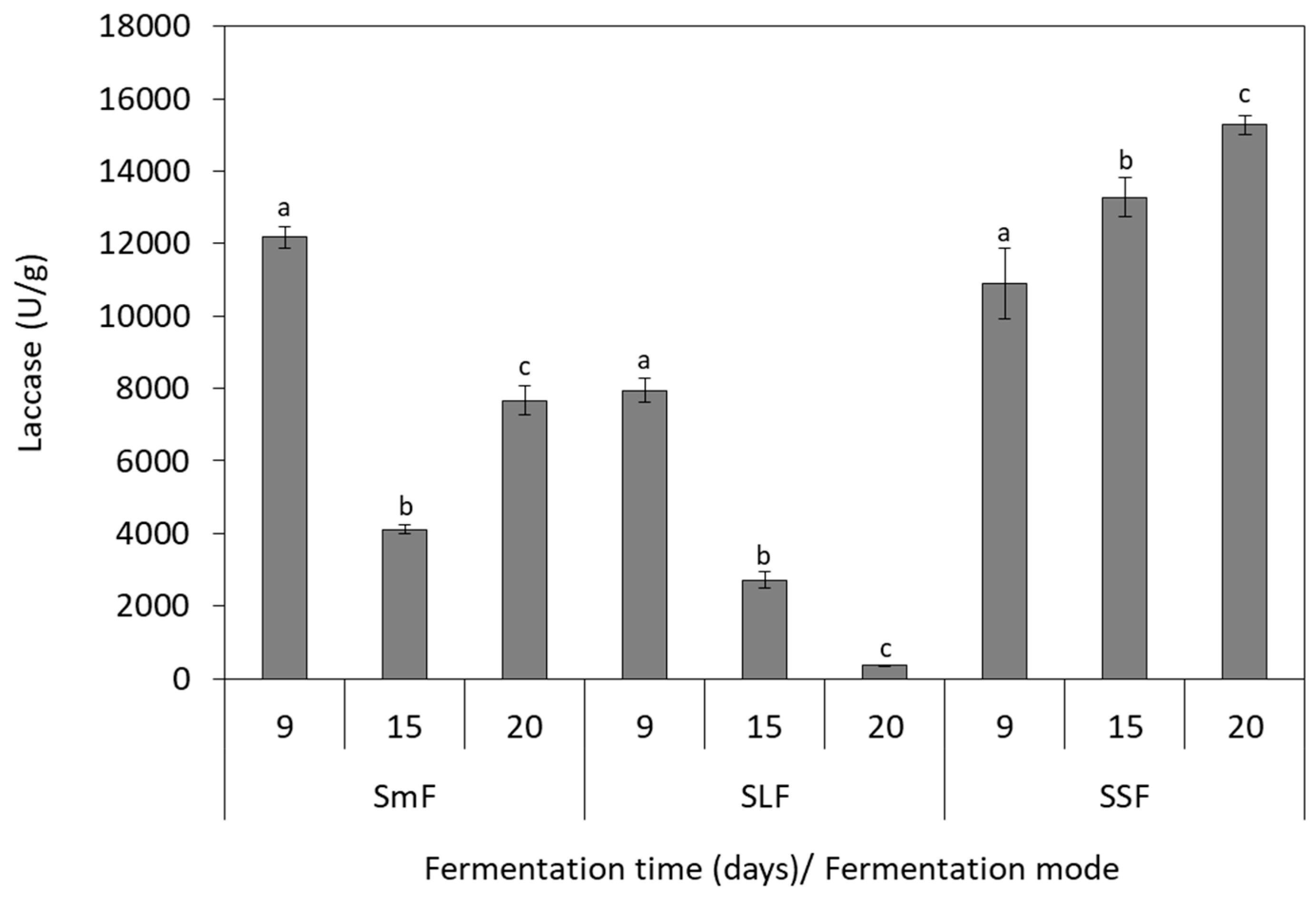
3.3. Laccase Production
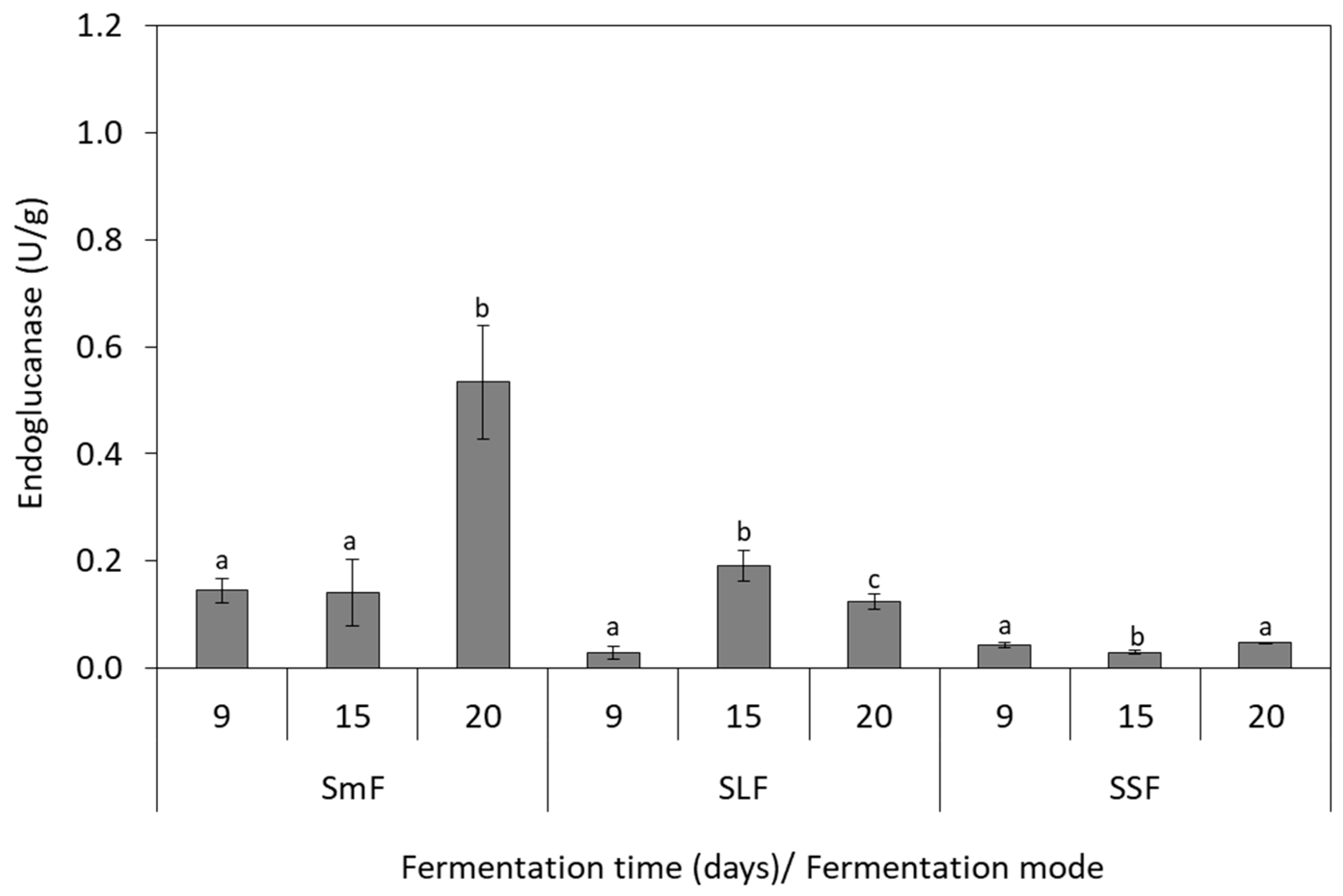
3.4. Endoglucanase Production
3.5. Evaluation of Grape Pomace for Fruiting Bodies Production
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef]
- Ghorai, S.; Banik, S.P.; Verma, D.; Chowdhury, S.; Mukherjee, S.; Khowala, S. Fungal biotechnology in food and feed processing. Food Res. Int. 2009, 42, 577–587. [Google Scholar] [CrossRef]
- Jayachandran, M.; Xiao, J.; Xu, B. A Critical Review on Health Promoting Benefits of Edible Mushrooms through Gut Microbiota. Int. J. Mol. Sci. 2017, 18, 1934. [Google Scholar] [CrossRef] [PubMed]
- Philippoussis, A.; Zervakis, G.; Diamantopoulou, P. Bioconversion of lignocellulosic wastes through the cultivation of the edible mushrooms Agrocybe aegerita, Volvariella volvacea and Pleurotus spp. World J. Microbiol. Biotechnol. 2001, 17, 191–200. [Google Scholar] [CrossRef]
- Chowdhary, P.; More, N.; Yadav, A.; Bharagava, R.N. Ligninolytic Enzymes: An Introduction and Applications in the Food Industry. In Enzymes in Food Biotechnology, 1st ed.; Kuddus, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 181–195. [Google Scholar]
- Fillat, Ú.; Ibarra, D.; Eugenio, M.; Moreno, A.; Tomás-Pejó, E.; Martín-Sampedro, R. Laccases as a Potential Tool for the Efficient Conversion of Lignocellulosic Biomass: A Review. Fermentation 2017, 3, 17. [Google Scholar] [CrossRef]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of Microbial Enzymes in Food Industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef]
- Chen, H.; Gan, J.; Ji, A.; Song, S.; Yin, L. Development of Double Network Gels Based on Soy Protein Isolate and Sugar Beet Pectin Induced by Thermal Treatment and Laccase Catalysis. Food Chem. 2019, 292, 188–196. [Google Scholar] [CrossRef]
- Rodríguez-Couto, S. Solid-State Fermentation for Laccases Production and Their Applications. In Current Developments in Biotechnology and Bioengineering; Pandey, A., Larroche, C., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 211–234. [Google Scholar]
- Chen, P.; Chen, W.; Jiang, S.; Zhong, Q.; Chen, H.; Chen, W. Synergistic Effect of Laccase and Sugar Beet Pectin on the Properties of Concentrated Protein Emulsions and Its Application in Concentrated Coconut Milk. Molecules 2018, 23, 2591. [Google Scholar] [CrossRef]
- Díaz, A.B.; Alvarado, O.; de Ory, I.; Caro, I.; Blandino, A. Valorization of grape pomace and orange peels: Improved production of hydrolytic enzymes for the clarification of orange juice. Food Bioprod. Process. 2013, 91, 580–586. [Google Scholar] [CrossRef]
- Economou, C.N.; Diamantopoulou, P.A.; Philippoussis, A.N. Valorization of spent oyster mushroom substrate and laccase recovery through successive solid state cultivation of Pleurotus, Ganoderma, and Lentinula strains. Appl. Microbiol. Biotechnol. 2017, 101, 5213–5222. [Google Scholar] [CrossRef]
- Bellettini, M.B.; Fiorda, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Maccari Junior, A.; Ribani, R.H. Factors affecting mushroom Pleurotus spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef] [PubMed]
- OIV, Organization Internationale de la Vigne et du Vin. Available online: http://www.oiv.int/public/medias/6307/oiv-press-release-global-economic-vitiviniculture-data-octob.pdf (accessed on 10 May 2019).
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of polyphenol-rich grape by-products in monogastric nutrition. A review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Lores, M.; Insam, H.; Domínguez, J. Strategies for recycling and valorization of grape marc. Crit. Rev. Biotechnol. 2019, 39, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Corbin, K.R.; Hsieh, Y.S.Y.; Betts, N.S.; Byrt, C.S.; Henderson, M.; Stork, J.; DeBolt, S.; Fincher, G.B.; Burton, R.A. Grape marc as a source of carbohydrates for bioethanol: Chemical composition, pre-treatment and saccharification. Bioresour. Technol. 2015, 193, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Petre, M.; Pătrulescu, F.; Teodorescu, R.I. Controlled Cultivation of Mushrooms on Winery and Vineyard Wastes. In Mushroom Biotechnology, 1st ed.; Petre, M., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 31–47. [Google Scholar]
- Commission Regulation (EC) No 152/2009 of 27 January 2009 Laying Down the Methods of Sampling and Analysis for the Official Control of Feed. Available online: http://data.europa.eu/eli/reg/2009/152/oj (accessed on 10 September 2012).
- Saeman, J.F.; Moore, W.E.; Mitchell, R.L.; Millett, M.A. Techniques for the determination of pulp constituents by quantitative paper chromatography. Tappi J. 1954, 34, 336–365. [Google Scholar]
- Rodríguez-Couto, S.; Longo, M.A.; Cameselle, C.; Sanromán, M.Á. Ligninolytic enzymes from corncob cultures of Phanerochaete chrysosporium in semi-solid state conditions. Acta Biotechnol. 1999, 19, 17–25. [Google Scholar] [CrossRef]
- Elisashvili, V.; Kachlishvili, E.; Tsiklauri, N.; Metreveli, E.; Tamar, K.; Agathos, S.N. Lignocellulose-degrading enzyme production by white-rot Basidiomycetes isolated from the forests of Georgia. World J. Microbiol. Biotechnol. 2009, 25, 331–339. [Google Scholar] [CrossRef]
- Waldron, K.W.; Selvendran, R.R. Composition of cell walls from different asparagus (Asparagus officinalis) tissues. Physiol. Plant. 1990, 80, 568–575. [Google Scholar] [CrossRef]
- Femenia, A.; Sánchez, E.S.; Sim al, S.; Rosselló, C. Effects of drying pretreatments on the cell wall composition of grape tissues. J. Agric. Food Chem. 1998, 46, 271–276. [Google Scholar] [CrossRef]
- Hoebler, C.; Barry, J.L.; David, A.; Delort-Laval, J. Rapid acid hydrolysis of plant cell wall polysaccharides and simplified quantitative determination of their neutral monosaccharides by gas-liquid chromatography. J. Agric. Food Chem. 1989, 37, 360–367. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Papadaki, A.; Diamantopoulou, P.; Papanikolaou, S.; Philippoussis, A. Evaluation of Biomass and Chitin Production of Morchella Mushrooms Grown on Starch-Based Substrates. Foods 2019, 8, 239. [Google Scholar] [CrossRef]
- Philippoussis, A.; Diamantopoulou, P.; Papadopoulou, K.; Lakhtar, H.; Roussos, S.; Parissopoulos, G.; Papanikolaou, S. Biomass, laccase and endoglucanase production by Lentinula edodes during solid state fermentation of reed grass, bean stalks and wheat straw residues. World J. Microbiol. Biotechnol. 2011, 27, 285–297. [Google Scholar] [CrossRef]
- Puoci, F.; Iemma, F.; Spizzirri, U.G.; Restuccia, D.; Pezzi, V.; Sirianni, R.; Manganaro, L.; Curcio, M.; Parisi, O.I.; Cirillo, G.; et al. Antioxidant activity of a Mediterranean food product: “fig syrup”. Nutrients 2011, 3, 317–329. [Google Scholar] [CrossRef]
- Mishra, A.; Kumar, S. Cyanobacterial biomass as N-supplement to agro-waste for hyper production of laccase from Pleurotus ostreatus in solid state fermentation. Process Biochem. 2007, 42, 681–685. [Google Scholar] [CrossRef]
- Sánchez, A.; Ysunza, F.; Beltrán-Garcıá, M.J.; Esqueda, M. Biodegradation of viticulture wastes by Pleurotus: A source of microbial and human food and its potential use in animal feeding. J. Agric. Food Chem. 2002, 50, 2537–2542. [Google Scholar] [CrossRef]
- Gaitán-Hernández, R.; Esqueda, M.; Gutiérrez, A.; Sánchez, A.; Beltrán-García, M.; Mata, G. Bioconversion of agrowastes by Lentinula edodes: The high potential of viticulture residues. Appl. Microbiol. Biotechnol. 2006, 71, 432–439. [Google Scholar] [CrossRef]
- Lakhtar, H.; Ismaili-Alaoui, M.; Philippoussis, A.; Perraud-Gaime, I.; Roussos, S. Screening of strains of Lentinula edodes grown on model olive mill wastewater in solid and liquid state culture for polyphenol biodegradation. Int. Biodeterior. Biodegrad. 2010, 64, 167–172. [Google Scholar] [CrossRef]
- Martinkova, L.; Kotik, M.; Markova, E.; Homolka, L. Biodegradation of phenolic compounds by Basidiomycota and its phenol oxidases: A review. Chemosphere 2016, 149, 373–382. [Google Scholar] [CrossRef]
- Patel, H.; Gupte, A.; Gupte, S. Effect of different culture conditions and inducers on production of laccase by a basidiomycete fungal isolate Pleurotus ostreatus HP-1 under solid state fermentation. Bioresources 2009, 4, 268–284. [Google Scholar]
- De Souza, D.F.; Tychanowicz, G.K.; Marques de Souza, C.G.; Peralta, R.M. Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheat bran solid state cultures. J. Basic Microbiol. 2006, 46, 126–134. [Google Scholar] [CrossRef]
- Stajić, M.; Persky, L.; Friesem, D.; Hadar, Y.; Wasser, S.P.; Nevo, E.; Vukojević, J. Effect of different carbon and nitrogen sources on laccase and peroxidases production by selected Pleurotus species. Enzym. Microb. Technol. 2006, 38, 65–73. [Google Scholar] [CrossRef]
- Tsioulpas, A.; Dimou, D.; Ikonomou, D.; Aggelis, G. Phenolic removal in olive oil mill waste-water by strains of Pleurotus spp. in respect to their phenol oxidase (laccase) activity. Bioresour. Technol. 2002, 84, 251–257. [Google Scholar] [CrossRef]
- Velázquez-Cedeño, M.A.; Mata, G.; Savoi, J.-M. Waste-reducing cultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffee pulp: Changes in the production of some lignocellulosic enzymes. World J. Microbiol. Biotechnol. 2002, 18, 201–207. [Google Scholar] [CrossRef]
- Prasad, K.K.; Mohan, S.V.; Bhaskar, Y.V.; Ramanaiah, S.V.; Babu, V.L.; Pati, B.R.; Sarma, P.N. Laccase production using Pleurotus ostreatus 1804 immobilized on PUF cubes in batch and packed bed reactors: Influence of culture conditions. J. Microbiol. 2005, 43, 301–307. [Google Scholar]
- Elisashvili, V.; Penninckx, M.; Kachlishvili, E.; Asatiani, M.; Kvesitadze, G. Use of Pleurotus dryinus for lignocellulolytic enzymes production in submerged fermentation of mandarin peels and tree leaves. Enzym. Microb. Technol. 2006, 38, 998–1004. [Google Scholar] [CrossRef]
- Stajić, M.; Persky, L.; Hadar, Y.; Friesem, D.; Duletić-Laušević, S.; Wasser, S.P.; Nevo, E. Effect of copper and manganese ions on activities of laccase and peroxidases in three Pleurotus species grown on agricultural wastes. Appl. Biochem. Biotechnol. 2006, 128, 87–96. [Google Scholar] [CrossRef]
- Akpinar, M.; Ozuturk Urek, R. Production of ligninolytic enzymes by solid-state fermentation using Pleurotus eryngii. Prep. Biochem. Biotechnol. 2012, 42, 582–597. [Google Scholar] [CrossRef]
- Elisashvili, V.; Kachlisvili, E.; Penninckx, M. Effect of growth substrate, method of fermentation and nitrogen source on lignocelluloses-degrading enzymes production by white rot basidiomycetes. J. Ind. Microbiol. Biotechnol. 2008, 35, 1531–1538. [Google Scholar] [CrossRef]
- Kurt, S.; Buyukalaca, S. Yield performances and changes in enzyme activities of Pleurotus spp. (P. ostreatus and P. sajor-caju) cultivated on different agricultural wastes. Bioresour. Technol. 2010, 101, 3164–3169. [Google Scholar] [CrossRef]
- Karpe, A.V.; Harding, I.H.; Palombo, E.A. Comparative degradation of hydrothermal pretreated winery grape wastes by various fungi. Ind. Crop. Prod. 2014, 59, 228–233. [Google Scholar] [CrossRef]
- Philippoussis, A.; Diamantopoulou, P. Agro-food industry wastes and agricultural residues conversion into high value products by mushroom cultivation. In Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Products (ICMBMP7), Arcachon, France, 4–7 October 2011. [Google Scholar]
- Ananbeh, K.; Almomany, A. Production of Oyster Mushroom (Pleurotus ostreatus) on Olive Cake Agro Waste. Agric. Sci. 2005, 32, 64–69. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pleurotus spp. | Biomass g (y)/glucosamine mg (x) | R2 |
|---|---|---|
| P. ostreatus | y = 0.0529x − 0.0735 | 0.99 |
| P. pulmonarius | y = 0.0579x − 0.0715 | 0.99 |
| Time (Days) | P. ostreatus | P. pulmonarius | ||||
|---|---|---|---|---|---|---|
| SSF | SmF | SLF | SSF | SmF | SLF | |
| 9 | 0.31 ± 0.01 a | 0.38 ± 0.01 a | 0.36 ± 0.01 a | 0.34 ± 0.01 a | 0.44 ± 0.01 a | 0.40 ± 0.01 a |
| 15 | 0.32 ± 0.02 a | 0.40 ± 0.01 a | 0.36 ± 0.01 a | 0.37 ± 0.01 b | 0.46 ± 0.00 b | 0.40 ± 0.01 a |
| 20 | 0.42 ± 0.01 b | 0.50 ± 0.02 b | 0.43 ± 0.01 b | 0.40 ± 0.01 c | 0.54 ± 0.01 c | 0.45 ± 0.02 b |
| Pleurotus spp. | Flushes | Earliness (Days) | Mushroom Number | Average Fresh Weight (g) | Total Yield (%) | BE (%) |
|---|---|---|---|---|---|---|
| P. ostreatus | 1 | 42 | 162 ± 3.7 | 25.9 ± 2.3 | 7.4 | 16.2 |
| P. pulmonarius | 1 | 35 | 176 ± 9.5 | 24.0 ± 1.8 | 14.4 | 31.4 |
| 2 | 45 | 43 ± 3.1 | 14.4 ± 1.4 | |||
| 3 | 55 | 12 ± 2.6 | 11.9 ± 1.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, A.; Kachrimanidou, V.; Papanikolaou, S.; Philippoussis, A.; Diamantopoulou, P. Upgrading Grape Pomace through Pleurotus spp. Cultivation for the Production of Enzymes and Fruiting Bodies. Microorganisms 2019, 7, 207. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070207
Papadaki A, Kachrimanidou V, Papanikolaou S, Philippoussis A, Diamantopoulou P. Upgrading Grape Pomace through Pleurotus spp. Cultivation for the Production of Enzymes and Fruiting Bodies. Microorganisms. 2019; 7(7):207. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070207
Chicago/Turabian StylePapadaki, Aikaterini, Vasiliki Kachrimanidou, Seraphim Papanikolaou, Antonios Philippoussis, and Panagiota Diamantopoulou. 2019. "Upgrading Grape Pomace through Pleurotus spp. Cultivation for the Production of Enzymes and Fruiting Bodies" Microorganisms 7, no. 7: 207. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070207