Genomic Insights of Dyadobacter tibetensis Y620-1 Isolated from Ice Core Reveal Genomic Features for Succession in Glacier Environment

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. General Features of the Dyadobacter Genomes
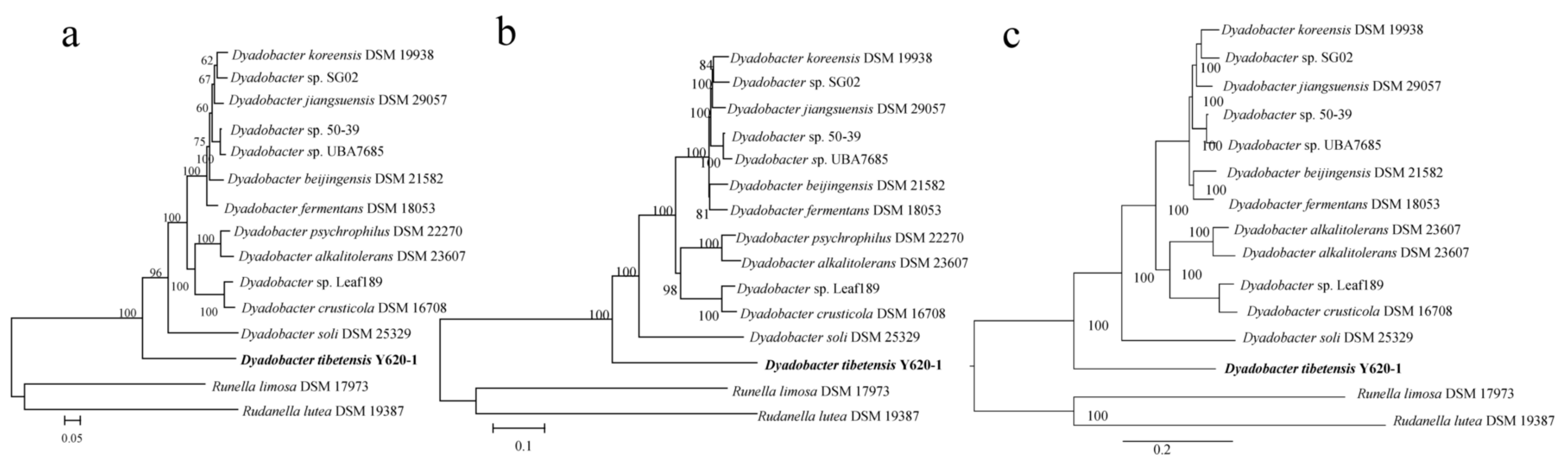
3.2. Distribution of Dyadobacter Strains in Their Phylogenomic Tree
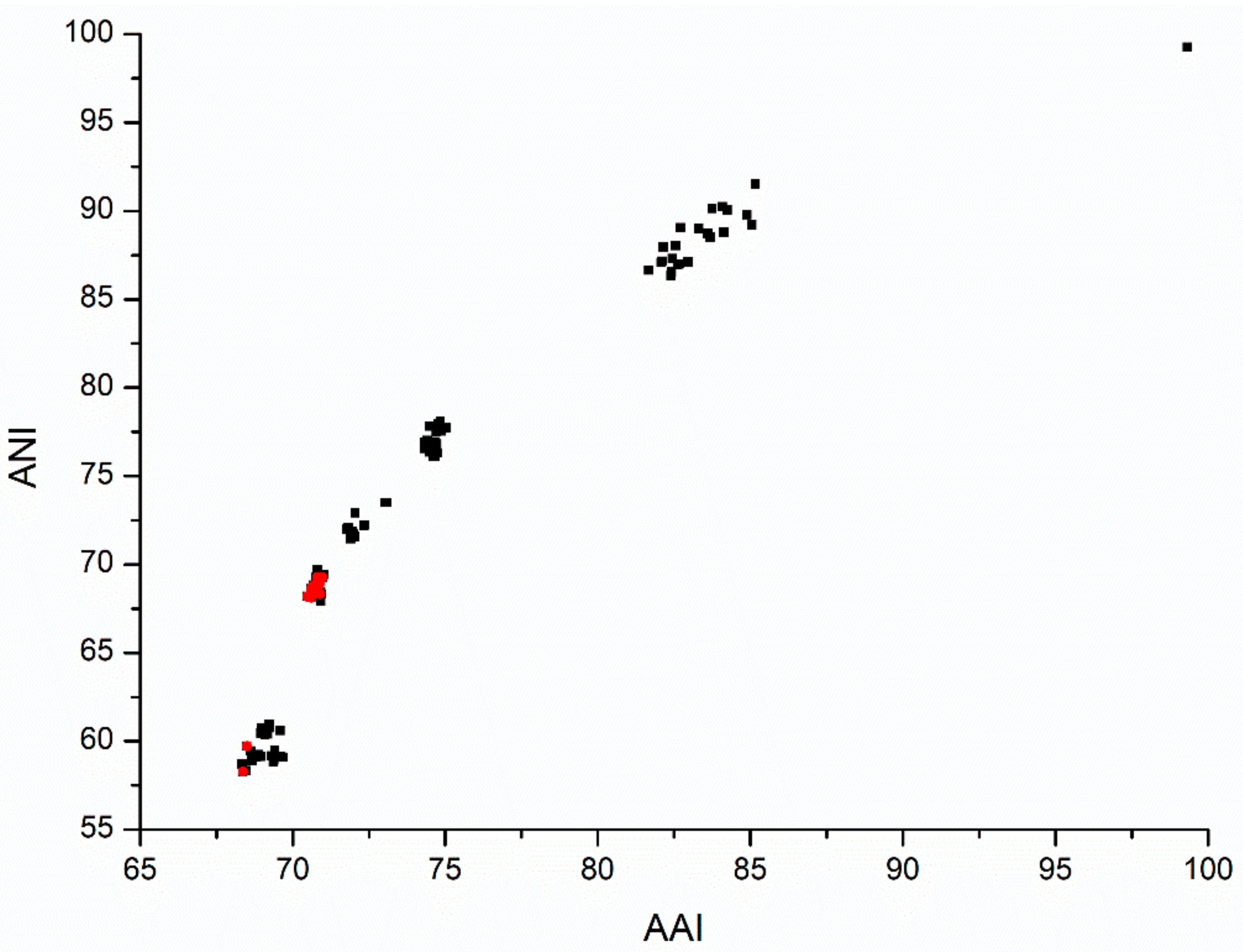
3.3. Average Nucleotide and Amino Acid Identity
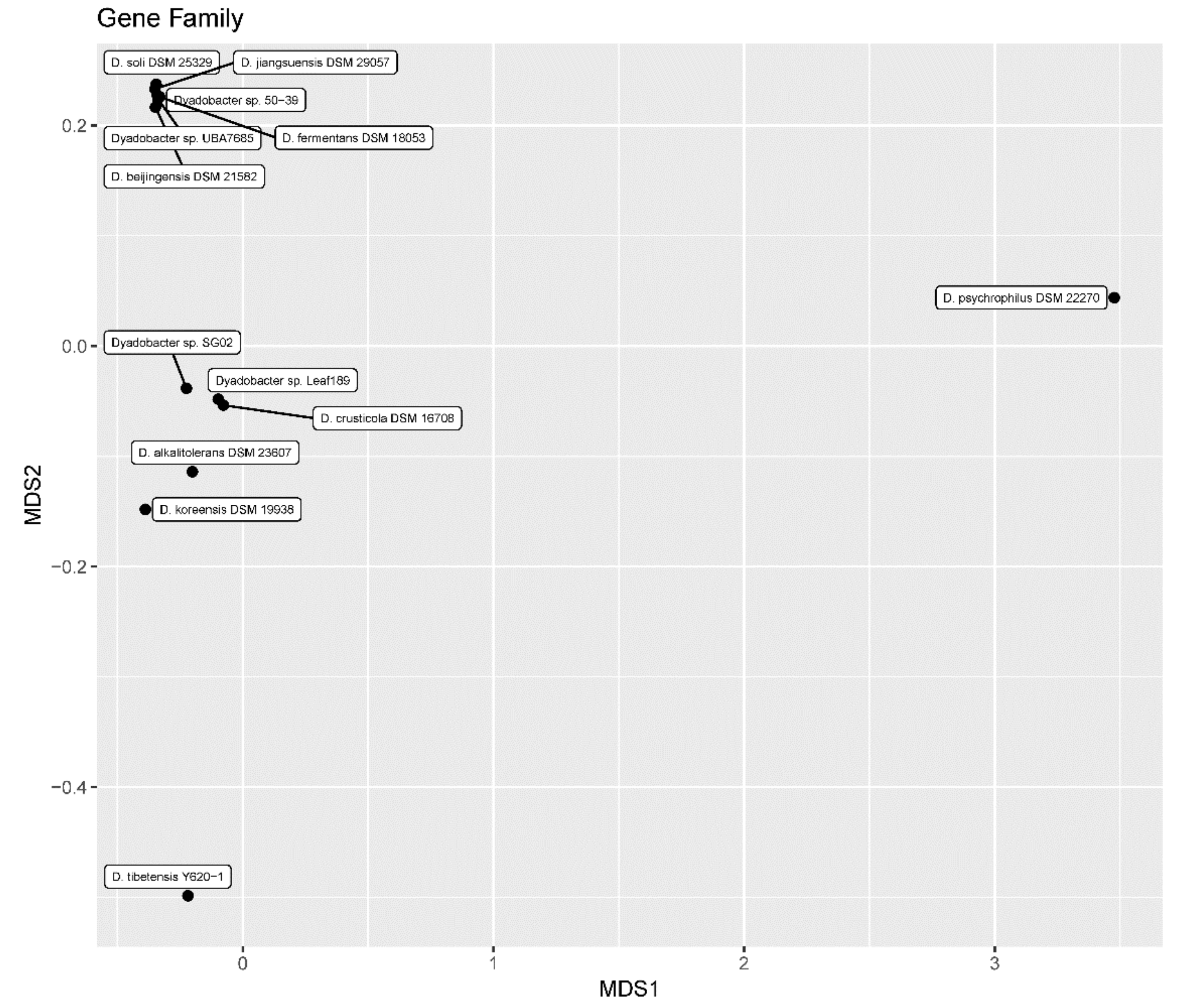
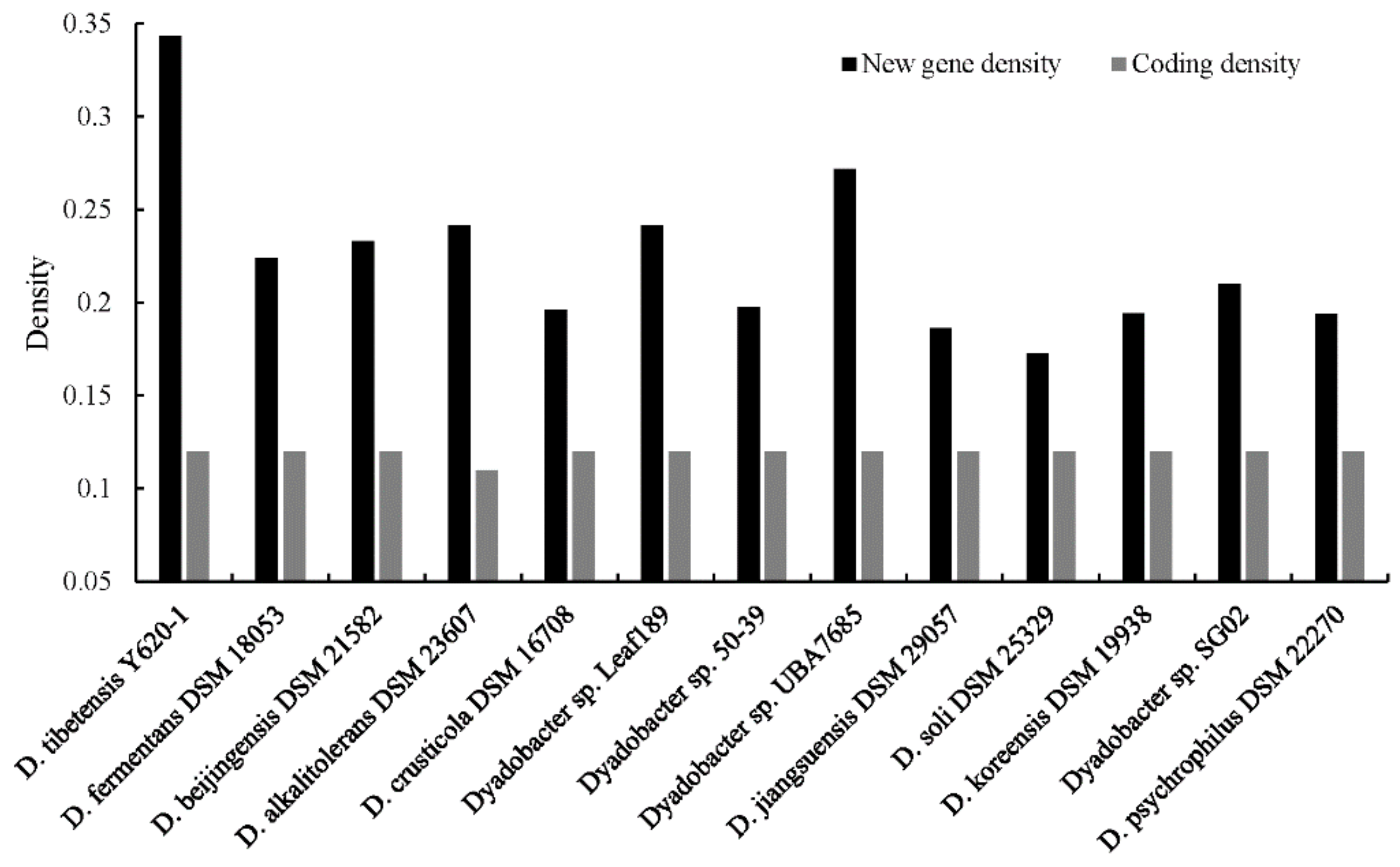
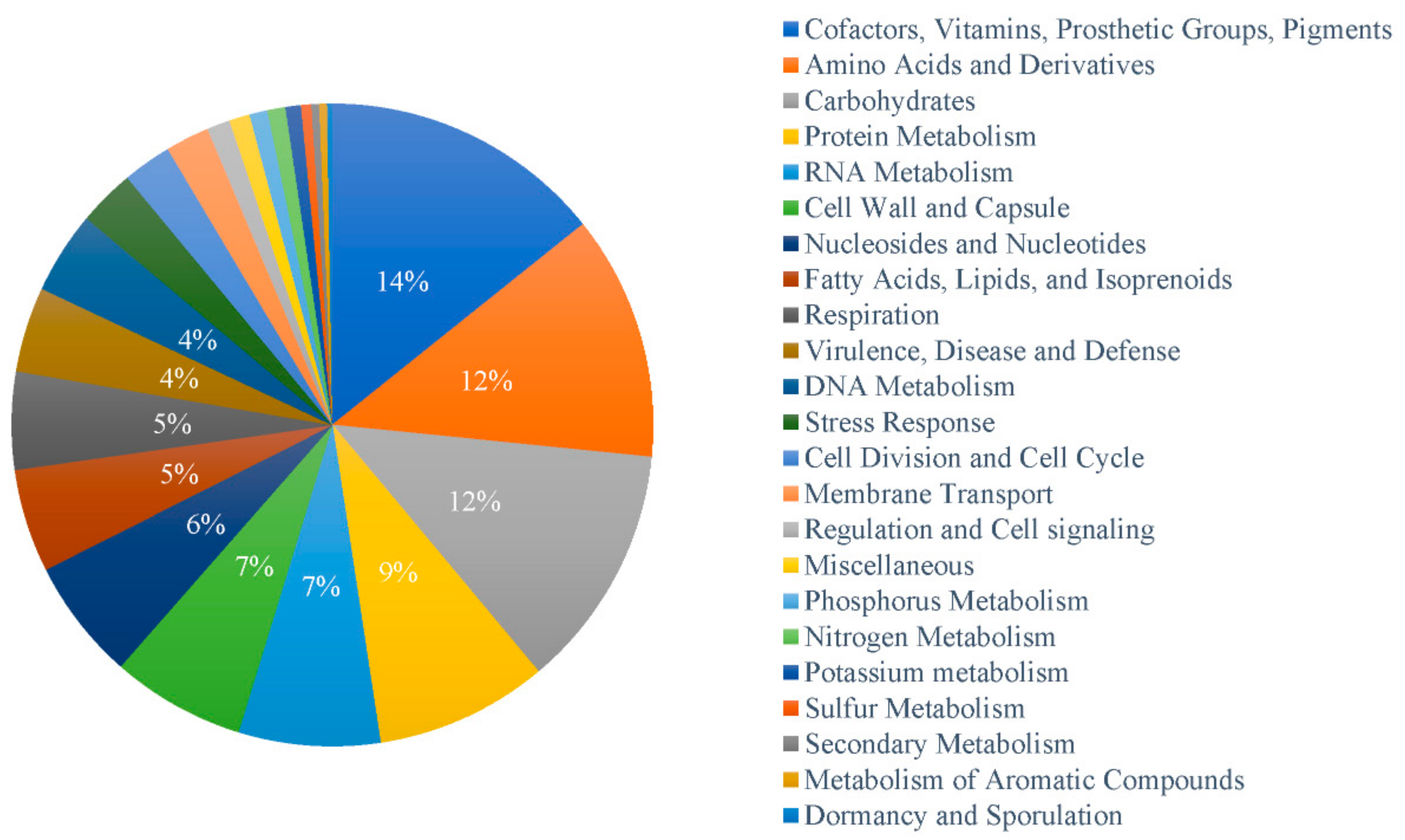
3.4. Distribution Pattern of Function Genes and Gene Families
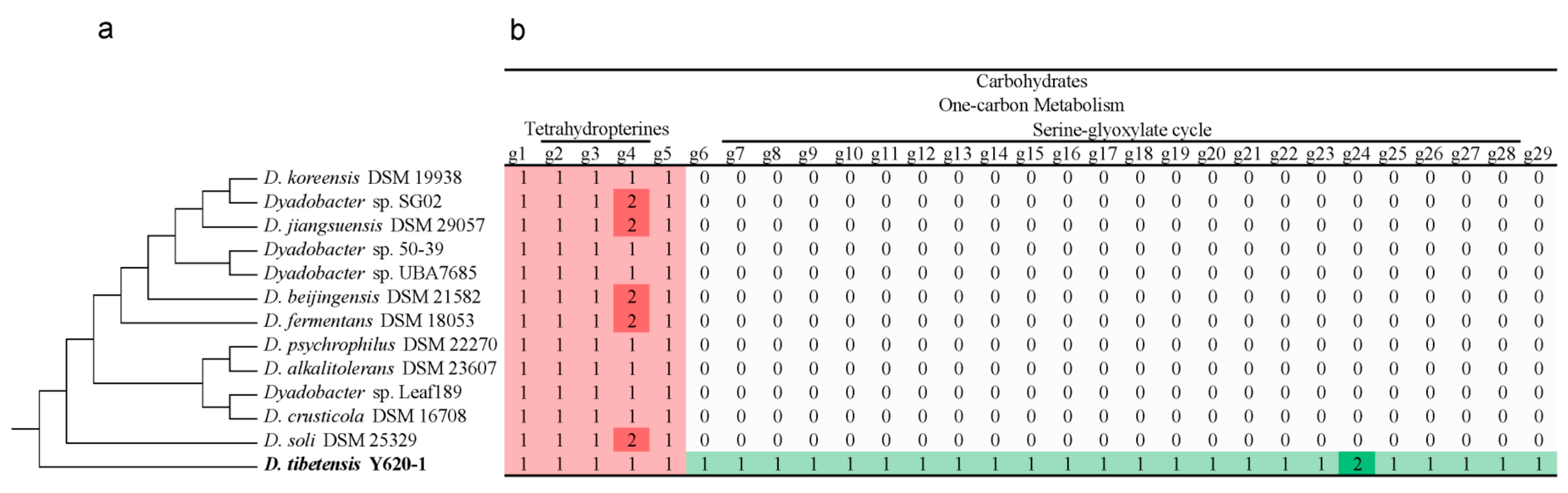
3.5. Specific Functions in One-Carbon Metabolism of D. tibetensis Y620-1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grinsted, A. An estimate of global glacier volume. Cryosphere 2013, 7, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Anesio, A.M.; Laybourn-Parry, J. Glaciers and ice sheets as a biome. Trends Ecol. Evol. 2012, 27, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Hodson, A.; Anesio, A.M.; Tranter, M.; Fountain, A.; Osborn, M.; Priscu, J.; Laybourn-Parry, J.; Sattler, B. Glacial ecosystems. Ecol. Monogr. 2008, 78, 41–67. [Google Scholar] [CrossRef]
- Larose, C.; Dommergue, A.; Vogel, T.M. Microbial nitrogen cycling in Arctic snowpacks. Environ. Res. Lett. 2013, 8, 035004. [Google Scholar] [CrossRef]
- Maccario, L.; Sanguino, L.; Vogel, T.M.; Larose, C. Snow and ice ecosystems: Not so extreme. Res. Microbiol. 2015, 166, 782–795. [Google Scholar] [CrossRef] [PubMed]
- Laybourn-Parry, J. No Place Too Cold. Science 2009, 324, 1521–1522. [Google Scholar] [CrossRef] [PubMed]
- Christner, B.C.; Mosley-Thompson, E.; Thompson, L.G.; Zagorodnov, V.; Sandman, K.; Reeve, J.N. Recovery and identification of viable bacteria immured in glacial ice. Icarus 2000, 144, 479–485. [Google Scholar] [CrossRef]
- Price, P.B. A habitat for psychrophiles in deep Antarctic ice. Proc. Natl. Acad. Sci. USA 2000, 97, 1247–1251. [Google Scholar] [CrossRef] [Green Version]
- Frasson, D.; Udovicic, M.; Frey, B.; Lapanje, A.; Zhang, D.C.; Margesin, R.; Sievers, M. Glaciimonas alpina sp. nov. isolated from alpine glaciers and reclassification of Glaciimonas immobilis Cr9-12 as the type strain of Glaciimonas alpina sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 1779–1785. [Google Scholar] [CrossRef]
- Margesin, R.; Schumann, P.; Zhang, D.C.; Redzic, M.; Zhou, Y.G.; Liu, H.C.; Schinner, F. Arthrobacter cryoconiti sp nov., a psychrophilic bacterium isolated from alpine glacier cryoconite. Int. J. Syst. Evol. Microbiol. 2012, 62, 397–402. [Google Scholar] [CrossRef]
- Margesin, R.; Sproer, C.; Schumann, P.; Schinner, F. Pedobacter cryoconitis sp. nov., a facultative psychrophile from alpine glacier cryoconite. Int. J. Syst. Evol. Microbiol. 2003, 53, 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, H.C.; Wen, Y.; Zhou, Y.G.; Xin, Y.H. Cryobacterium flavum sp. nov. and Cryobacterium luteum sp. nov., isolated from glacier ice. Int. J. Syst. Evol. Microbiol. 2012, 62, 1296–1299. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Kumari, M.; Kiran, S.; Salwan, R.; Mayilraj, S.; Chhibber, S.; Gulati, A. Chryseobacterium glaciei sp. nov., isolated from the surface of a glacier in the Indian trans-Himalayas. Int. J. Syst. Evol. Microbiol. 2018, 68, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Liu, H.C.; Zhang, J.L.; Zhou, Y.G.; Xin, Y.H. Flavobacterium xueshanense sp. nov. and Flavobacterium urumqiense sp. nov., two psychrophilic bacteria isolated from glacier ice. Int. J. Syst. Evol. Microbiol. 2012, 62, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, Y.Q.; Wang, N.L.; Yao, T.D.; Jiao, N.Z.; Liu, H.C.; Zhou, Y.G.; Xu, B.Q.; Liu, X.B. Massilia yuzhufengensis sp. nov., isolated from an ice core. Int. J. Syst. Evol. Microbiol. 2013, 63, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, Y.Q.; Yao, T.D.; Wang, N.L.; Xu, B.Q.; Jiao, N.Z.; Liu, H.C.; Zhou, Y.G.; Liu, X.B.; Wang, Y.N. Dyadobacter tibetensis sp. nov., isolated from glacial ice core. Int. J. Syst. Evol. Microbiol. 2013, 63, 3636–3639. [Google Scholar] [CrossRef] [PubMed]
- Bajerski, F.; Ganzert, L.; Mangelsdorf, K.; Lipski, A.; Busse, H.J.; Padur, L.; Wagner, D. Herbaspirillum psychrotolerans sp. nov., a member of the family Oxalobacteraceae from a glacier forefield. Int. J. Syst. Evol. Microbiol. 2013, 63, 3197–3203. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Jiang, F.; Chang, X.L.; Qiu, X.; Ren, L.Z.; Qu, Z.H.; Deng, S.S.; Da, X.Y.; Fang, C.X.; Peng, F. Flavobacterium collinsense sp. nov., isolated from a till sample of an Antarctic glacier. Int. J. Syst. Evol. Microbiol. 2016, 66, 172–177. [Google Scholar] [CrossRef]
- Qiu, X.; Qu, Z.H.; Jiang, F.; Ren, L.Z.; Chang, X.L.; Kan, W.J.; Fang, C.X.; Peng, F. Pedobacter huanghensis sp. nov. and Pedobacter glacialis sp. nov., isolated from Arctic glacier foreland. Int. J. Syst. Evol. Microbiol. 2014, 64, 2431–2436. [Google Scholar] [CrossRef]
- Srinivas, T.N.R.; Manasa, P.; Begum, Z.; Sunil, B.; Sailaja, B.; Singh, S.K.; Prasad, S.; Shivaji, S. Iodobacter arcticus sp. nov., a psychrotolerant bacterium isolated from meltwater stream sediment of an Arctic glacier. Int. J. Syst. Evol. Microbiol. 2013, 63, 2800–2805. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Yu, Y.; Liu, Y.; Li, H.R. Psychrobacter glaciei sp. nov., isolated from the ice core of an Arctic glacier. Int. J. Syst. Evol. Microbiol. 2016, 66, 1792–1798. [Google Scholar] [CrossRef] [PubMed]
- Price, P.B. Microbial life in glacial ice and implications for a cold origin of life. FEMS Microbiol. Ecol. 2007, 59, 217–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Kaushik, R.; Saxena, A.K. Cold active hydrolytic enzymes production by psychrotrophic Bacilli isolated from three sub-glacial lakes of NW Indian Himalayas. J. Basic. Microbiol. 2016, 56, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.F.; Tiedje, J.M. Coping with our cold planet. Appl. Environ. Microbiol. 2008, 74, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- De Maayer, P.; Anderson, D.; Cary, C.; Cowan, D.A. Some like it cold: Understanding the survival strategies of psychrophiles. EMBO Rep. 2014, 15, 508–517. [Google Scholar] [CrossRef]
- Chelius, M.K.; Triplett, E.W. Dyadobacter fermentans gen. nov., sp. nov., a novel Gram-negative bacterium isolated from surface-sterilized Zea mays stems. Int. J. Syst. Evol. Microbiol. 2000, 50, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.L.; Sun, P.B.; Wang, X.M.; Qiu, T.L.; Lv, F.Y.; Yuan, M.; Yang, M.M.; Sun, J.G. Dyadobacter endophyticus sp. nov., an endophytic bacterium isolated from maize root. Int. J. Syst. Evol. Microbiol. 2016, 66, 4022–4026. [Google Scholar] [PubMed]
- Liu, Y.Q.; Hu, A.Y.; Shen, L.; Yao, T.D.; Jiao, N.Z.; Wang, N.L.; Xu, B.Q. Draft genome sequence of Dyadobacter tibetensis type strain (Y620-1) isolated from glacial ice. Stand. Genom. Sci. 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- CompareM. Available online: https://github.com/dparks1134/CompareM (accessed on 15 November 2018).
- ANI Calculator. Available online: http://enve-omics.ce.gatech.edu/ani/ (accessed on 20 January 2019).
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Torsten, S. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar]
- Yarza, P.; Richter, M.; Peplies, J.; Euzeby, J.; Amann, R.; Schleifer, K.H.; Ludwig, W.; Glöckner, F.O.; Rosselló-Móra, R. The All-Species Living Tree project: A 16S rRNA-based phylogenetic tree of all sequenced type strains. Syst. Appl. Microbiol. 2008, 31, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.H. Computational Molecular Evolution; Oxford University Press: Great Britain, UK, 2006. [Google Scholar]
- Segata, N.; Bornigen, D.; Morgan, X.C.; Huttenhower, C. PhyloPhlAn is a new method for improved phylogenetic and taxonomic placement of microbes. Nat. Commun. 2013, 4, 2304. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Rinke, C.; Chuvochina, M.; Chaumeil, P.A.; Woodcroft, B.J.; Evans, P.N.; Hugenholtz, P.; Tyson, G.W. Recovery of nearly 8000 metagenome-assembled genomes substantially expands the tree of life. Nat. Microbiol. 2017, 2, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- FastOrtho Software (--pv_cutoff 1-e5 --pi_cutoff 70 --pmatch_cutoff 70). Available online: http://enews.patricbrc.org/fastortho/ (accessed on 16 October 2018).
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Wang, L.; Chen, L.; Ling, O.; Li, C.C.; Tao, Y.; Wang, M. Dyadobacter jiangsuensis sp. nov., a methyl red degrading bacterium isolated from a dye-manufacturing factory. Int. J. Syst. Evol. Microbiol. 2015, 65, 1138–1143. [Google Scholar] [CrossRef]
- Lee, M.; Woo, S.G.; Park, J.; Yoo, S.A. Dyadobacter soli sp. nov., a starch-degrading bacterium isolated from farm soil. Int. J. Syst. Evol. Microbiol. 2010, 60, 2577–2582. [Google Scholar] [CrossRef]
- Zhang, D.C.; Liu, H.C.; Xin, Y.H.; Zhou, Y.G.; Schinner, F.; Margesin, R. Dyadobacter psychrophilus sp. nov., a psychrophilic bacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2010, 60, 1640–1643. [Google Scholar] [CrossRef]
- Dong, Z.; Guo, X.Y.; Zhang, X.X.; Qiu, F.B.; Sun, L.; Gong, H.L.; Zhang, F.Y. Dyadobacter beijingensis sp. nov, isolated from the rhizosphere of turf grasses in China. Int. J. Syst. Evol. Microbiol. 2007, 57, 862–865. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Dai, J.; Zhang, L.; Mo, Z.Y.; Wang, Y.; Li, Y.W.; Ji, S.M.; Fang, C.X.; Zheng, C.Y. Dyadobacter alkalitolerans sp. nov., isolated from desert sand. Int. J. Syst. Evol. Microbiol. 2009, 59, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.S.N.; Garcia-Pichel, F. Dyadobacter crusticola sp. nov., from biological soil crusts in the Colorado Plateau, USA, and an emended description of the genus Dyadobacter Chelius and Triplett 2000. Int. J. Syst. Evol. Microbiol. 2005, 55, 1295–1299. [Google Scholar] [CrossRef] [PubMed]
- Baik, K.S.; Kim, M.S.; Kim, E.M.; Kim, H.R.; Seong, C.N. Dyadobacter koreensis sp. nov., isolated from fresh water. Int. J. Syst. Evol. Microbiol. 2007, 57, 1227–1231. [Google Scholar] [CrossRef]
- Rawat, S.R.; Bromberg, Y.; Häggblom, M.M.; Männistö, M.K. Comparative genomic and physiological analysis provides insights into the role of Acidobacteria in organic carbon utilization in Arctic tundra soils. FEMS Microbiol. Ecol. 2012, 82, 341–355. [Google Scholar] [CrossRef]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic insights that advance the species definition for prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, K.T.; Tiedje, J.M. Trends between gene content and genome size in prokaryotic species with larger genomes. Proc. Natl. Acad. Sci. USA 2004, 101, 3160–3165. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Seshadri, R.; Varghese, N.J.; Eloe-Fadrosh, E.A.; Meier-Kolthoff, J.P.; Goker, M.; Coates, R.C.; Hadjithomas, M.; Pavlopoulos, G.A.; Paez-Espino, D.; et al. 1003 reference genomes of bacterial and archaeal isolates expand coverage of the tree of life. Nat. Biotechnol. 2017, 35, 676–683. [Google Scholar] [CrossRef]
- Sawatdeenarunat, C.; Nguyen, D.; Surendra, K.C.; Shrestha, S.; Rajendran, K.; Oechsner, H.; Xie, L.; Khanal, S.K. Anaerobic biorefinery: Current status, challenges, and opportunities. Bioresour. Technol. 2016, 215, 304–313. [Google Scholar] [CrossRef]
- Smejkalova, H.; Erb, T.J.; Fuchs, G. Methanol assimilation in Methylobacterium extorquens AM1: Demonstration of all enzymes and their regulation. PLoS ONE 2010, 5, e13001. [Google Scholar] [CrossRef]
- Murakami, T.; Segawa, T.; Bodington, D.; Dial, R.; Takeuchi, N.; Kohshima, S.; Hongoh, Y. Census of bacterial microbiota associated with the glacier ice worm Mesenchytraeus solifugus. FEMS Microbiol. Ecol. 2015, 91, fiv003. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Greening, C.; Vanwonterghem, I.; Carere, C.R.; Bay, S.K.; Steen, J.A.; Montgomery, K.; Lines, T.; Beardall, J.; van Dorst, J.; et al. Atmospheric trace gases support primary production in Antarctic desert surface soil. Nature 2017, 552, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Singh, S.M.; Dhakephalkar, P. Diversity, cold active enzymes and adaptation strategies of bacteria inhabiting glacier cryoconite holes of High Arctic. Extremophiles 2014, 18, 229–242. [Google Scholar] [CrossRef] [PubMed]
- McCrimmon, D.O.; Bizimis, M.; Holland, A.; Ziolkowski, L.A. Supraglacial microbes use young carbon and not aged cryoconite carbon. Org. Geochem. 2018, 118, 63–72. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Mueller, D.; Vincent, W.F. Arctic Ice Shelves and Ice Islands; Springer Netherlands: New York, NY, USA, 2017; pp. 227–260. [Google Scholar]
- Smith, G.J.; Foster, R.A.; McKnight, D.M.; Lisle, J.T.; Littmann, S.; Kuypers, M.M.M.; Foreman, C.M. Microbial formation of labile organic carbon in Antarctic glacial environments. Nat. Geosci. 2017, 10. [Google Scholar] [CrossRef]
- Anesio, A.M.; Bellas, C.M. Are low temperature habitats hot spots of microbial evolution driven by viruses? Trends Microbiol. 2011, 19, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Bahl, J.; Lau, M.C.; Smith, G.J.; Vijaykrishna, D.; Cary, S.C.; Lacap, D.C.; Lee, C.K.; Papke, R.T.; Warren-Rhodes, K.A.; Wong, F.K.; et al. Ancient origins determine global biogeography of hot and cold desert cyanobacteria. Nat. Commun. 2011, 2, 163. [Google Scholar] [CrossRef] [PubMed]
- Choudoir, M.J.; Buckley, D.H. Phylogenetic conservatism of thermal traits explains dispersal limitation and genomic differentiation of Streptomyces sister-taxa. ISME J. 2018, 12, 2176–2186. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Assembly No. | Isolation Sources | Completeness | Contamination | GC | Size (Mbp) | CDS | CRISPRs | rRNAs | tRNAs | CspA | CspG | New Gene Dendity | Coding Density |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D. alkalitolerans DSM 23607 | GCA_000428845.1 | Desert sand | 100.00 | 0.00 | 45.67 | 6.29 | 5496 | 0 | 3 | 35 | 3 | 1 | 0.24 | 0.11 |
| D. beijingensis DSM 21582 | GCA_000382205.1 | Soil | 99.69 | 0.30 | 52.08 | 7.38 | 6030 | 0 | 6 | 40 | 4 | 1 | 0.23 | 0.12 |
| D. crusticola DSM 16708 | GCA_000701505.1 | Soil | 100.00 | 0.00 | 46.73 | 6.07 | 5141 | 0 | 3 | 40 | 2 | 1 | 0.20 | 0.12 |
| D. fermentans DSM 18053 | GCA_000023125.1 | Plant | 99.70 | 0.30 | 51.54 | 6.97 | 5853 | 0 | 12 | 43 | 2 | 1 | 0.22 | 0.12 |
| D. jiangsuensis DSM 29057 | GCA_003014695.1 | Soil | 100.00 | 0.60 | 50.26 | 8.27 | 6854 | 0 | 2 | 38 | 2 | 1 | 0.19 | 0.12 |
| D. koreensis DSM 19938 | GCA_900108855.1 | Fresh water | 99.70 | 0.89 | 41.26 | 7.34 | 6140 | 0 | 7 | 40 | 1 | 1 | 0.19 | 0.12 |
| D. psychrophilus DSM 22270 | GCA_900167945.1 | Soil | 99.70 | 0.30 | 45.05 | 6.74 | 5722 | 0 | 4 | 34 | 2 | 1 | 0.19 | 0.12 |
| D. soli DSM 25329 | GCA_900101885.1 | Soil | 99.70 | 0.00 | 50.47 | 8.74 | 7339 | 0 | 6 | 40 | 1 | 1 | 0.17 | 0.12 |
| D. tibetensis Y620-1 | GCA_000566685.1 | Ice core | 99.70 | 0.30 | 43.45 | 5.31 | 4275 | 6 | 3 | 37 | 5 | 2 | 0.34 | 0.12 |
| Dyadobacter sp. 50-39 | GCA_001898145.1 | Bioreactor | 99.70 | 0.60 | 50.24 | 7.72 | 6563 | 5 | 2 | 40 | 4 | 1 | 0.20 | 0.12 |
| Dyadobacter sp. Leaf189 | GCA_001424405.1 | Leaf | 99.70 | 0.60 | 47.00 | 6.07 | 5141 | 0 | 3 | 40 | 3 | 1 | 0.24 | 0.12 |
| Dyadobacter sp. SG02 | GCA_900109045.1 | Root | 99.70 | 0.74 | 50.23 | 8.48 | 7043 | 0 | 2 | 38 | 6 | 1 | 0.21 | 0.12 |
| Dyadobacter sp. UBA7685 | GCA_002482895.1 | Water | 97.02 | 0.00 | 50.58 | 5.18 | 4436 | 0 | 0 | 30 | 2 | 1 | 0.27 | 0.12 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Liu, Y.; Wang, N.; Adhikari, N.P. Genomic Insights of Dyadobacter tibetensis Y620-1 Isolated from Ice Core Reveal Genomic Features for Succession in Glacier Environment. Microorganisms 2019, 7, 211. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070211
Shen L, Liu Y, Wang N, Adhikari NP. Genomic Insights of Dyadobacter tibetensis Y620-1 Isolated from Ice Core Reveal Genomic Features for Succession in Glacier Environment. Microorganisms. 2019; 7(7):211. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070211
Chicago/Turabian StyleShen, Liang, Yongqin Liu, Ninglian Wang, and Namita Paudel Adhikari. 2019. "Genomic Insights of Dyadobacter tibetensis Y620-1 Isolated from Ice Core Reveal Genomic Features for Succession in Glacier Environment" Microorganisms 7, no. 7: 211. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070211