Inhibition and Interactions of Campylobacter jejuni from Broiler Chicken Houses with Organic Acids

 , , , and
, , , and
Abstract
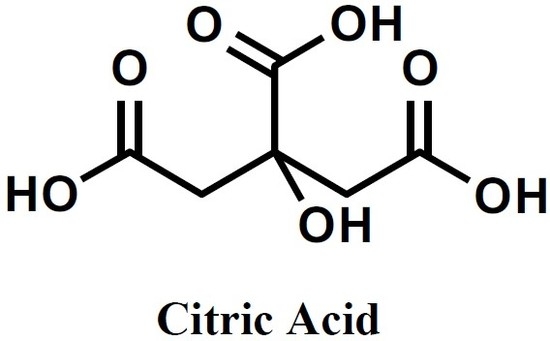
:
1. Introduction
2. Materials and Methods
2.1. Campylobacter jejuni
2.2. Susceptibility Testing of C. jejuni Strains with Organic Acids
2.3. Determination of pH in Wells of the 96-Well Microplates at the C. jejuni MICs
2.4. The Calculated Ratio of Undissociated/Dissociated OAs
2.5. Statistics
3. Results
3.1. Susceptibility Studies of OAs Against the C. jejuni strains
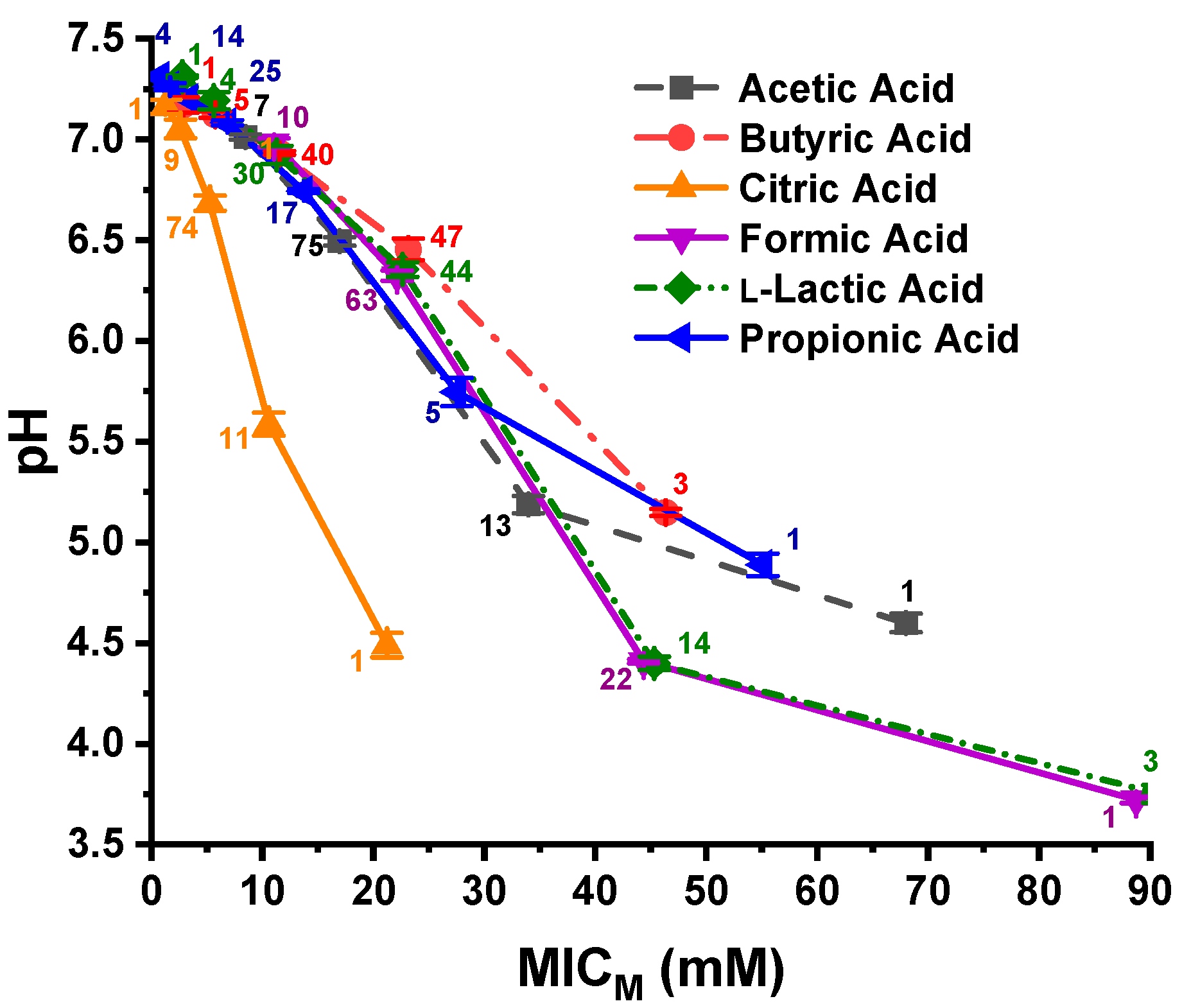
3.2. Experimentally Determined pH at the C. jejuni MICs for the 96 Strains Evaluated with the OAs
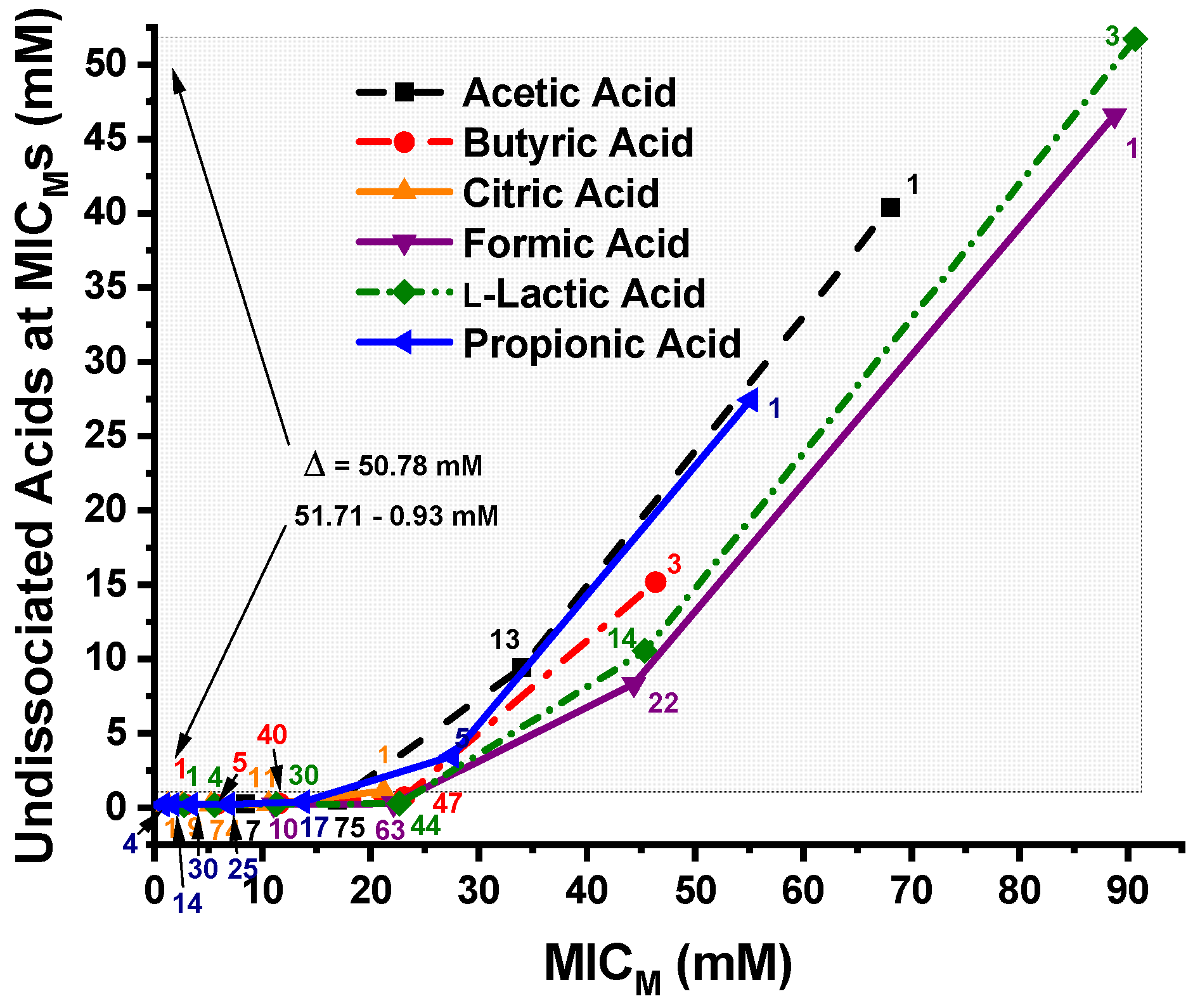
3.3. Calculated Undissociated OA Concentrations at the C. jejuni MICMs
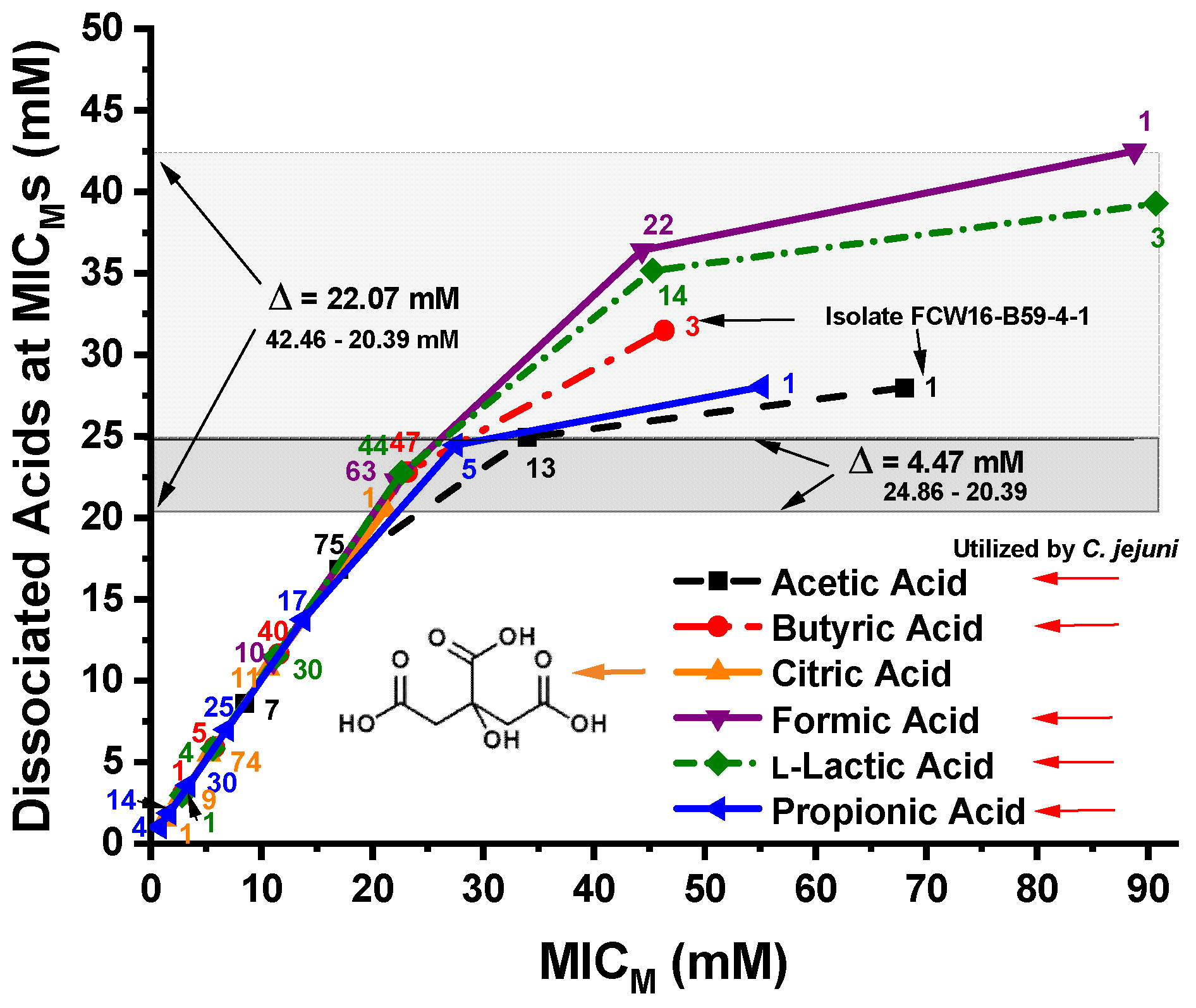
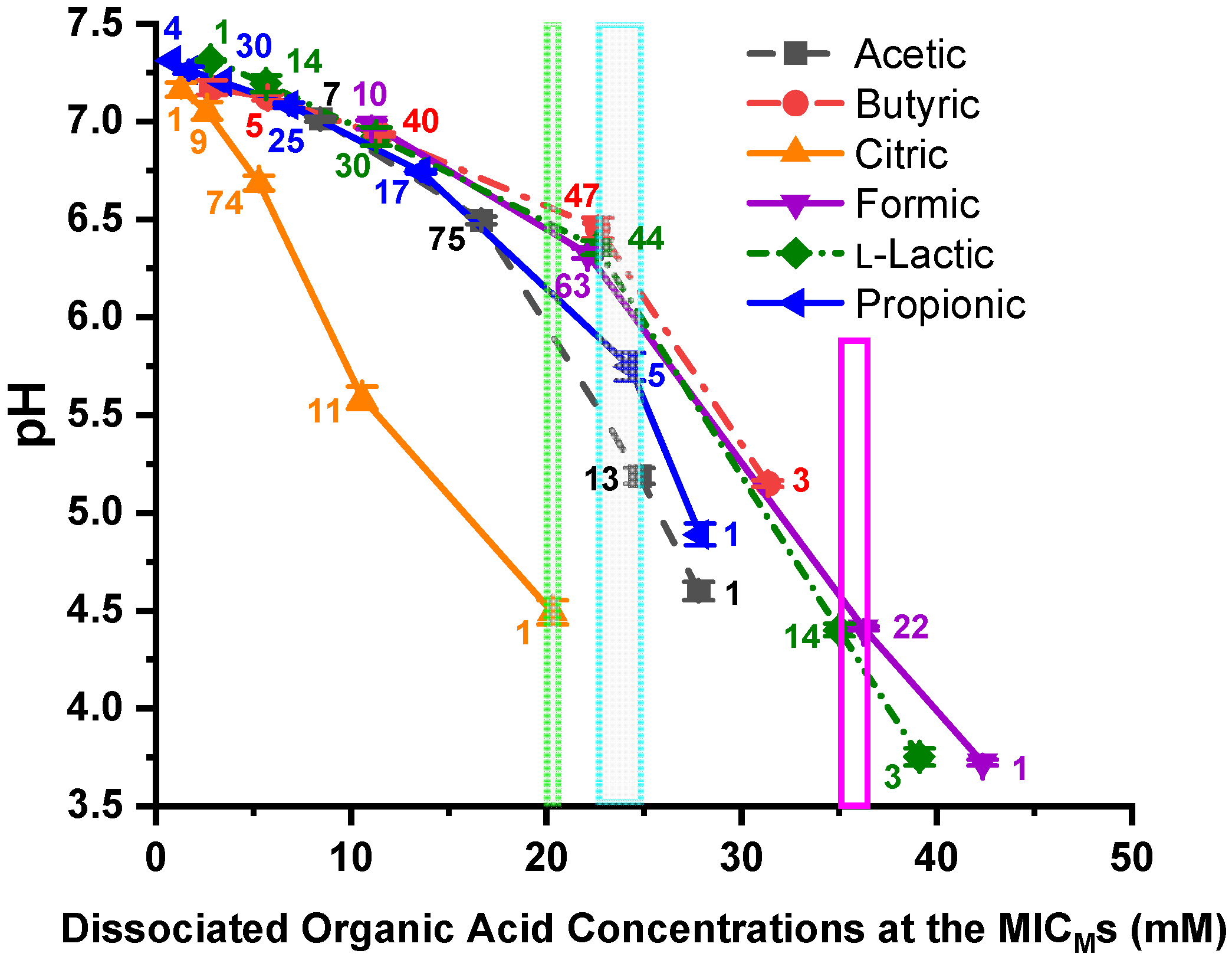
3.4. Calculated Dissociated OA Concentrations at the C. jejuni MICMs
4. Discussion
4.1. Differences in pH Between Solutions of Different OAs at the MICM
4.2. Relationships of MICM Concentrations and pH to the Undissociated OA Concentrations
4.3. Relationships of MICM Concentrations and pH to the Dissociated OA Concentrations
4.4. Mechanism by Which Organic Acids Control Bacteria Based on Our Studies
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moore, J.E.; Corcoran, D.; Dooley, J.S.G.; Fanning, S.; Lucey, B.; Matsuda, M.; McDowell, D.A.; Mégraud, F.; Millar, B.C.; O’Mahony, R.; et al. Campylobacter. Vet. Res. 2005, 36, 351–382. [Google Scholar] [CrossRef]
- Epps, S.V.R.; Harvey, R.B.; Hume, M.E.; Phillips, T.D.; Anderson, R.C.; Nisbet, D.J. Foodborne Campylobacter: Infections, metabolism, pathogenesis and reservoirs. Int. J. Environ. Res. Public Health 2013, 10, 6292–6304. [Google Scholar] [CrossRef]
- Hofreuter, D. Defining the metabolic requirements for the growth and colonization capacity of Campylobacter jejuni. Cellul. Infect. Microbiol. 2014, 4, 137. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2015. EFSA J. 2016, 14, 4634. [Google Scholar]
- World Health Organization (WHO). Campylobacter. Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter (accessed on 11 April 2019).
- Penner, J.L. The Genus Campylobacter: A decade of progress. Clin. Microbiol. Rev. 1988, 1, 157–172. [Google Scholar] [CrossRef]
- Blaser, M.J.; Taylor, D.N.; Feldman, R.A. Epidemiology of Campylobacter jejuni infections. Epidemiol. Rev. 1983, 5, 157–176. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Burden of foodborne illness: Findings. 2011. Available online: https://www.cdc.gov/foodborneburden/2011-foodborne-estimates.html (accessed on 13 April 2019).
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Briffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Nachamkin, I.; Szymanski, C.M.; Blaser, M.J. Campylobacter, 3rd ed.; ASM Press: Washington, DC, USA, 2008. [Google Scholar]
- Baer, A.A.; Miller, M.J.; Dilger, A.C. Pathogens of interest to the pork industry: A review of research on interventions to assure food safety. Compr. Rev. Food Sci. F. 2013, 12, 183–217. [Google Scholar] [CrossRef]
- Sifré, E.; Salha, B.A.; Ducournau, A.; Floch, P.; Chardon, H.; Mégraud, F.; Lehours, P. EUCAST recommendations for antimicrobial susceptibility testing applied to the three main Campylobacter species isolated in humans. J. Microbiol. Methods 2015, 119, 206–213. [Google Scholar] [CrossRef]
- Roop, R.M., II; Smibert, R.M.; Johnson, J.L.; Krieg, N.R. Differential characteristics of catalase-positive campylobacters correlated with DNA homology groups. Can. J. Microbiol. 1984, 30, 938–951. [Google Scholar] [CrossRef]
- Hébert, G.A.; Edmonds, P.; Brenner, D.J. DNA relatedness among strains of Campylobacter jejuni and Campylobacter coli with divergent serogroup and hippurate reactions. J. Clin. Microbiol. 1984, 20, 138–140. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Campylobacter, Salmonella led bacterial foodborne illnesses in 2016. 2017. Available online: https://www.cdc.gov/media/releases/2017/p0420-campylobacter-salmonella.html (accessed on 15 April 2019).
- Olson, C.K.; Ethelberg, S.; van Pelt, W.; Tauxe, R.V. Epidemiology of Campylobacter jejuni infections in industrialized nations. In Campylobacter, 3rd ed.; Nachamkin, I., Szymanski, C.M., Blaser, M.J., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 163–189. [Google Scholar]
- Kramer, J.M.; Frost, J.A.; Bolton, F.J.; Wareing, D.R. Campylobacter contamination of raw meat and poultry at retail sale: Identification of multiple types and comparison with isolates from human infection. J. Food Prot. 2000, 63, 1654–1659. [Google Scholar] [CrossRef]
- Kapperud, G.; Espeland, G.; Wahl, E.; Walde, A.; Herikstad, H.; Gustavsen, S.; Tveit, I.; Natås, O.; Bevanger, L.; Digranes, A. Factors associated with increased and decreased risk of Campylobacter infection: A prospective case-control study in Norway. Am. J. Epidemiol. 2003, 158, 234–242. [Google Scholar] [CrossRef]
- Humphrey, T.; Jørgensen, F. Pathogens on meat and infection in animals – Establishing a relationship using Campylobacter and Salmonella as examples. Meat Sci. 2006, 74, 89–97. [Google Scholar] [CrossRef]
- Lindmark, H.; Boqvist, S.; Ljungström, M.; Ågren, P.; Björkholm, B.; Engstrand, L. Risk factors for campylobacteriosis: An epidemiological surveilance study of patients and retail poultry. J. Clin. Microbiol. 2009, 47, 2616–2619. [Google Scholar] [CrossRef]
- Molatová, Z.; Skřivanová, E.; Macias, B.; McEwan, N.R.; Březina, P.; Marounek, M. Susceptibility of Campylobacter jejuni to organic acids and monoacylglycerols. Folia Microbiol. 2010, 55, 215–220. [Google Scholar] [CrossRef]
- Weinberger, M.; Moran-Gilad, J.; Rokney, A.; Davidov, Y.; Agmon, V.; Peretz, C.; Valinsky, L. Molecular epidemiology of Campylobacter jejuni infection in Israel—A nationwide study. Clin. Microbiol. Infect. 2016, 22, 1005.e9–1005.e15. [Google Scholar] [CrossRef]
- Coker, A.O.; Isokpehi, R.D.; Thomas, B.N.; Amisu, K.O.; Obi, C.L. Human campylobacteriosis in developing countries. Emerg. Infect. Dis. 2002, 8, 237–243. [Google Scholar] [CrossRef]
- Black, R.E.; Levine, M.M.; Clements, M.L.; Hughes, T.P.; Blaser, M.J. Experimental Campylobacter jejuni infection in humans. J. Infect. Dis. 1988, 157, 472–479. [Google Scholar] [CrossRef]
- Kothary, M.H.; Babu, U.S. Infective dose of foodborne pathogens in volunteers: A review. J. Food Saf. 2001, 21, 49–73. [Google Scholar] [CrossRef]
- Wachsmuth, I.K.; Sparling, P.H.; Barrett, T.J.; Potter, M.E. Enterohemorrhagic Escherichia coli in the United States. FEMS Immunol. Med. Microbiol. 1997, 18, 233–239. [Google Scholar] [CrossRef]
- Keener, K.M.; Bashor, M.P.; Curtis, P.A.; Sheldon, B.W.; Kathariou, S. Comprehensive review of Campylobacter and poultry processing. Comp. Rev. Food Sci. Food Saf. 2004, 3, 105–116. [Google Scholar] [CrossRef]
- Lee, M.D. Overview of avian Campylobacter infection, Merck Manual Veterinary Manual. Available online: https://merckvetmanual.com/poultry/avian-campylobacter-infection/overview-of-avian-campylobacter-infection (accessed on 15 April 2019).
- Sahin, O.; Kassem, I.I.; Shen, Z.; Lin, J.; Rajashekara, G.; Zhang, Q. Campylobacter in poultry: Ecololgy and potential interventions. Avian Dis. 2015, 59, 185–200. [Google Scholar] [CrossRef]
- Byrd, J.A.; Hargis, B.M.; Caldwell, D.J.; Bailey, R.H.; Herron, K.L.; McReynolds, J.L.; Brewer, R.L.; Anderson, R.C.; Bischoff, K.M.; Callaway, T.R.; et al. Effect of lactic acid administration in the drinking water during preslaughter feed withdrawal on Salmonella and Campylobacter contamination of broilers. Poult. Sci. 2001, 80, 278–283. [Google Scholar] [CrossRef]
- Chaveerach, P.; Keuzenkamp, D.A.; Urlings, H.A.P.; Lipman, L.J.A.; van Knapen, F. In vitro study on the effect of organic acids on Campylobacter jejuni/coli populations in mixtures of water and feed. Poult. Sci. 2002, 81, 621–628. [Google Scholar] [CrossRef]
- Chaveerach, P.; Keuzenkamp, D.A.; Lipman, L.J.A.; van Knapen, F. Effect of organic acids in drinking water for young broilers on Campylobacter infection, volatile fatty acid production, gut microflora and histological cell changes. Poult. Sci. 2004, 83, 330–334. [Google Scholar] [CrossRef]
- Skånseng, B.; Kaldhusdal, M.; Moen, B.; Gjevre, A.-G.; Johannessen, G.S.; Sekelja, M.; Trosvik, P. Prevention of intestinal Campylobacter jejuni colonization in broilers by combinations of in-feed organic acids. J. Appl. Microbiol. 2010, 109, 1265–1273. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.A.; Mohnl, M.; Schatzmayr, G.; Böhm, J. Control strategies for Campylobacter infection in poultry production. Worlds Poult. Sci. J. 2013, 69, 57–76. [Google Scholar] [CrossRef]
- Navarro, M.; Stanley, R.; Cusack, A.; Sultanbawa, Y. Combinations of plant-derived compounds against Campylobacter in vitro. J. Appl. Poult. Res. 2015, 24, 352–363. [Google Scholar] [CrossRef]
- Birk, T.; Grønlund, A.C.; Christensen, B.B.; Knøchel, S.; Lohse, K.; Rosenquist, H. Effect of organic acids and marination ingredients on the survival of Campylobacter jejuni on meat. J. Food Protect. 2010, 73, 258–265. [Google Scholar] [CrossRef]
- Ricke, S.C. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Sofos, J.N.; Busta, F.F. Antimicrobial activity of sorbate. J. Food Protect. 1981, 44, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Blocher, J.C.; Busta, F.F.; Sofos, J.N. Influence of potassium sorbate and pH on ten strains of type A and B Clostridium botulinum. J. Food Sci. 1982, 47, 2028–2032. [Google Scholar] [CrossRef]
- Ray, B.; Sandine, W.E. Acetic, propionic, and lactic acids of starter culture bacteria as biopreservatives. In Food Biopreservatives of Microbial Origin; Ray, B., Daeschel, M., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1992; pp. 103–136. [Google Scholar]
- Breidt, F., Jr.; Hayes, J.S.; McFeeters, R.F. Independent effects of acetic acid and pH on survival of Escherichia coli in simulated acidified pickle products. J. Food Protect. 2004, 67, 12–18. [Google Scholar] [CrossRef]
- Leeson, S. Balancing science versus societal issues in poultry nutrition. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 071. [Google Scholar] [CrossRef]
- Van Deun, K.; Haesebrouch, F.; Van Immerseel, F.; Ducatelle, R.; Pasmans, F. Short-chain fatty acids and l-lactate as feed additives to control Campylobacter jejuni infections in broilers. Avian Pathol. 2008, 37, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Presser, K.A.; Ross, T.; Ratkowsky, D.A. Modelling the growth limits (growth/no growth interface) of Escherichia coli as a function of temperature, pH, lactic acid concentration, and water activity. Appl. Environ. Microbiol. 1998, 64, 1773–1779. [Google Scholar]
- Carpenter, C.E.; Broadbent, J.R. External concentration of organic acid anions and pH: Key independent variables for studying how organic acids inhibit growth of bacteria in mildly acidic foods. J. Food Sci. 2009, 74, R12–R15. [Google Scholar] [CrossRef]
- Bushell, F.M.L.; Tonner, P.D.; Jabbari, S.; Schmid, A.K.; Lund, P.A. Synergistic impacts of organic acids and pH on growth of Pseudomonas aeruginosa: A comparison of parametric and Bayesian non-parametric methods to model growth. Front. Microbiol. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Beier, R.C.; Poole, T.L.; Brichta-Harhay, D.M.; Anderson, R.C.; Bischoff, K.M.; Hernandez, C.A.; Bono, J.L.; Arthur, T.M.; Nagaraja, T.G.; Crippen, T.L.; et al. Disinfectant and antibiotic susceptibility profiles of Escherichia coli O157:H7 strains from cattle carcasses, feces, and hides and ground beef from the United States. J. Food Protect. 2013, 76, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Franz, E.; Bono, J.L.; Mandrell, R.E.; Fratamico, P.M.; Callaway, T.R.; Andrews, K.; Poole, T.L.; Crippen, T.L.; Sheffield, C.L.; et al. Disinfectant and antimicrobial susceptibility profiles of the big six non-O157 Shiga toxin-producing Escherichia coli strains from food animals and humans. J. Food Protect. 2016, 79, 1355–1370. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Callaway, T.R.; Andrews, K.; Poole, T.L.; Crippen, T.L.; Anderson, R.C.; Nisbet, D.J. Interactions of organic acids with Salmonella strains from feedlot water-sprinkled cattle. J. Food Chem. Nanotechnol. 2017, 3, 60–66. [Google Scholar] [CrossRef]
- Beier, R.C.; Harvey, R.B.; Poole, T.L.; Hume, M.E.; Crippen, T.L.; Highfield, L.D.; Alali, W.Q.; Andrews, K.; Anderson, R.C.; Nisbet, D.J. Interactions of organic acids with vancomycin-resistant Enterococcus faecium isolated from community wastewater in Texas. J. Appl. Microbiol. 2018, 126, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Harvey, R.B.; Hernandez, C.A.; Hume, M.E.; Andrews, K.; Droleskey, R.E.; Davidson, M.K.; Bodeis-Jones, S.; Young, S.; Duke, S.E.; et al. Interactions of organic acids with Campylobacter coli from swine. PLoS ONE 2018, 13, e0202100. [Google Scholar] [CrossRef]
- Alakomi, H.-L.; Skyttä, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic acid permeabilizes Gram-negative bacteria by disrupting the outer membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef]
- Stern, N.J.; Wojton, B.; Kwiatek, K. A differential-selective medium and dry ice-generated atmosphere for recovery of Campylobacter jejuni. J. Food Protect. 1992, 55, 514–517. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria; Approved Guideline—2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010; M45-A2; Volume 30, No 18. [Google Scholar]
- TREK Diagnostic Systems. TREK materials and methods for sensititre susceptibility plates for Campylobacter. Available online: http://www.uniscience.co.kr/data/trds/sensi_manuals/Campylobacter_panel.pdf (accessed on 18 April 2019).
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Beier, R.C.; Bischoff, K.M.; Ziprin, R.L.; Poole, T.L.; Nisbet, D.J. Chlorhexidine susceptibility, virulence factors, and antibiotic resistance of beta-hemolytic Escherichia coli isolated from neonatal swine with diarrhea. Bull. Environ. Contam. Toxicol. 2005, 75, 835–844. [Google Scholar] [CrossRef]
- Beier, R.C.; Duke, S.E.; Ziprin, R.L.; Harvey, R.B.; Hume, M.E.; Poole, T.L.; Scott, H.M.; Highfield, L.D.; Alali, W.Q.; Andrews, K.; et al. Antibiotic and disinfectant susceptibility profiles of vancomycin-resistant Enterococcus faecium (VRE) isolated from community wastewater in Texas. Bull. Environ. Contam. Toxicol. 2008, 80, 188–194. [Google Scholar] [CrossRef]
- Beier, R.C.; Anderson, P.N.; Hume, M.E.; Poole, T.L.; Duke, S.E.; Crippen, T.L.; Sheffield, C.L.; Caldwell, D.J.; Byrd, J.A.; Anderson, R.C.; et al. Characterization of Salmonella enterica isolates from turkeys in commercial processing plants for resistance to antibiotics, disinfectants, and a growth promoter. Foodborne Pathog. Dis. 2011, 8, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Callaway, T.R.; Andrews, K.; Poole, T.L.; Crippen, T.L.; Anderson, R.C.; Nisbet, D.J. Disinfectant and antimicrobial susceptibility profiles of Salmonella strains from feedlot water-sprinkled cattle: Hides and feces. J. Food Chem. Nanotechnol. 2017, 3, 50–59. [Google Scholar] [CrossRef]
- Helmenstine, A.M. Henderson-Hasselbalch Equation and Example. 2014. Available online: http://chemistry.about.com/od/acidsbase1/a/hendersonhasselbalch.htm (accessed on 22 April 2019).
- Shaheen, B.W.; Miller, M.E.; Oyarzabal, O.A. In vitro survival at low pH and acid adaptation response of Campylobacter jejuni and Campylobacter coli. J. Food Saf. 2007, 27, 326–343. [Google Scholar] [CrossRef]
- Lund, P.; Tramonti, A.; De Biase, D. Coping with low pH: Molecular strategies in neutralophilic bacteria. FEMS Microbiol. Rev. 2014, 38, 1091–1125. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.N.; Pandey, R.; Palyada, K.; Naikare, H.; Stintzi, A. Identification of Campylobacter jejuni genes involved in the response to acidic pH and stomach transit. Appl. Environ. Microbiol. 2008, 74, 1583–1597. [Google Scholar] [CrossRef] [PubMed]
- Bearson, B.L.; Lee, I.S.; Casey, T.A. Escherichia coli O157:H7 glutamate- and arginine-dependent acid-resistance systems protect against oxidative stress during extreme acid challenge. Microbiology 2009, 155, 805–812. [Google Scholar] [CrossRef]
- Birk, T.; Wik, M.T.; Lametsch, R.; Knøchel, S. Acid stress response and protein induction in Campylobacter jejuni isolates with different acid tolerance. BMC Microbiol. 2012, 12, 174. [Google Scholar] [CrossRef]
- Varsaki, A.; Murphy, C.; Barczynska, A.; Jordan, K.; Carroll, C. The acid adaptive tolerance response in Campylobacter jejuni induces a global response, as suggested by proteomics and microarrays. Microb. Biotechnol. 2015, 8, 974–988. [Google Scholar] [CrossRef]
- Axelsson-Olsson, D.; Svensson, L.; Olofsson, J.; Salomon, P.; Waldenström, J.; Ellström, P.; Olsen, B. Increase in acid tolerance of Campylobacter jejuni through coincubation with amoebae. Appl. Environ. Microbiol. 2010, 76, 4194–4200. [Google Scholar] [CrossRef]
- Kiggins, E.M.; Plastridge, W.N. Some metabolic activities of Vibrio fetus of bovine origin. J. Bacteriol. 1958, 75, 205–208. [Google Scholar]
- Elharrif, Z.; Mégraud, F. Characterization of thermophilic Campylobacter. I. Carbon-substrate utilization tests. Curr. Microbiol. 1986, 13, 117–122. [Google Scholar] [CrossRef]
- Beier, R.C.; Foley, S.L.; Davidson, M.K.; White, D.G.; McDermott, P.F.; Bodeis-Jones, S.; Zhao, S.; Andrews, K.; Crippen, T.L.; Sheffield, C.L.; et al. Characterization of antibiotic and disinfectant susceptibility profiles among Pseudomonas aeruginosa veterinary isolates recovered during 1994–2003. J. Appl. Microbiol. 2014, 118, 326–342. [Google Scholar] [CrossRef] [PubMed]
- Hirshfield, I.N.; Terzulli, S.; O’Byrne, C. Weak organic acids: A panoply of effects on bacteria. Sci. Prog. 2003, 86, 245–269. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, A.H.M.; Ketley, J.M.; Park, S.F.; Penn, C.W. The role of iron in Campylobacter gene regulation, metabolism and oxidative stress defense. FEMS Microbiol. Rev. 2002, 26, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Shaw, F.L.; Mulholland, F.; Le Gall, G.; Porcelli, I.; Hart, D.J.; Pearson, B.M.; van Vliet, A.H.M. Selenium-dependent biogenesis of formate dehydrogenase in Campylobacter jejuni is controlled by the fdhTU accessory genes. J. Bacteriol. 2012, 194, 3814–3823. [Google Scholar] [CrossRef] [PubMed]
- Kassem, I.I.; Candelero-Rueda, R.A.; Esseili, K.A.; Rajashekara, G. Formate simultaneously reduces oxidase activity and enhances respiration in Campylobacter jejuni. Sci. Rep. (Nat.) 2017, 6, 40117. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.A.; Grant, A.J.; Hurd, D.; Harrison, M.; Guccione, E.J.; Kelly, D.J.; Maskell, D.J. Metabolite and transcriptome analysis of Campylobacter jejuni in vitro growth reveals a stationary-phase physiological switch. Microbiology 2009, 155, 80–94. [Google Scholar] [CrossRef]
- Luethy, P.M.; Huynh, S.; Ribardo, D.A.; Winter, S.E.; Parker, C.T.; Hendrixson, D.R. Microbiota-derived short-chain fatty acids modulate expression of Campylobacter jejuni determinants required for commensalism and virulence. mBio 2017, 8, e00407–e00417. [Google Scholar] [CrossRef]
- Thomas, M.T.; Shepherd, M.; Poole, R.K.; van Vliet, A.H.M.; Kelly, D.J.; Pearson, B.M. Two respiratory enzyme systems in Campylobacter jejuni NCTC 11168 contribute to growth on l-lactate. Environ. Microbiol. 2011, 13, 48–61. [Google Scholar] [CrossRef]
- Wagley, S.; Newcombe, J.; Laing, E.; Yusul, E.; Sambles, C.M.; Studholme, D.J.; La Ragione, R.M.; Titball, R.W.; Champion, O.L. Differences in carbon source utilisation distinguish Campylobacter jejuni from Campylobacter coli. BMC Microbiol. 2014, 14, 262. [Google Scholar] [CrossRef]
- Shin, S.-Y.; Hwang, H.-J.; Kim, W.J. Inhibition of Campylobacter jejuni in chicken by ethanol, hydrogen peroxide, and organic acids. J. Microbiol. Biotechnol. 2001, 11, 418–422. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Acids | MIC (μg/mL) | MICM (mM) | No. of C. jejuni Strains |
|---|---|---|---|
| Acetic Acid | 4096 | 68.21 | 1 |
| 2048 | 34.10 | 13 | |
| 1024 | 17.05 | 75 | |
| 512 | 8.53 | 7 | |
| Butyric Acid | 4096 | 46.49 | 3 |
| 2048 | 23.25 | 47 | |
| 1024 | 11.62 | 40 | |
| 512 | 5.81 | 5 | |
| 256 | 2.91 | 1 | |
| Citric Acid | 4096 | 21.32 | 1 |
| 2048 | 10.66 | 11 | |
| 1024 | 5.33 | 74 | |
| 512 | 2.67 | 9 | |
| 256 | 1.33 | 1 | |
| Formic Acid | 4096 | 89.00 | 1 |
| 2048 | 44.5 | 22 | |
| 1024 | 22.25 | 63 | |
| 512 | 11.12 | 10 | |
| l-Lactic Acid | 8192 | 90.94 | 3 |
| 4096 | 45.47 | 14 | |
| 2048 | 22.74 | 44 | |
| 1024 | 11.37 | 30 | |
| 512 | 5.68 | 4 | |
| 256 | 2.84 | 1 | |
| Propionic Acid | 4096 | 55.29 | 1 |
| 2048 | 27.65 | 5 | |
| 1024 | 13.82 | 17 | |
| 512 | 6.91 | 25 | |
| 256 | 3.46 | 30 | |
| 128 | 1.73 | 14 | |
| 64 | 0.86 | 4 |
| Organic Acid | Median | Mode | Range | 90th Percentile |
|---|---|---|---|---|
| Acetic Acid | ||||
| MIC (μg/mL) | 1024 | 1024 | 512–4096 | 2048 |
| MICM (mM) | 17.05 | 17.05 | 8.53–68.21 | 34.1 |
| Butyric Acid | ||||
| MIC (μg/mL) | 2048 | 2048 | 256–4096 | 2048 |
| MICM (mM) | 23.24 | 23.25 | 2.91–46.49 | 23.25 |
| Citric Acid | ||||
| MIC (μg/mL) | 1024 | 1024 | 256–4096 | 2048 |
| MICM (mM) | 5.33 | 5.33 | 1.33–21.32 | 10.66 |
| Formic Acid | ||||
| MIC (μg/mL) | 1024 | 1024 | 512–4096 | 2048 |
| MICM (mM) | 22.25 | 22.25 | 11.12–89.00 | 44.5 |
| l-Lactic Acid | ||||
| MIC (μg/mL) | 2048 | 2048 | 256–8192 | 4096 |
| MICM (mM) | 22.74 | 22.74 | 2.84–90.94 | 45.47 |
| Propionic Acid | ||||
| MIC (μg/mL) | 256 | 256 | 64–4096 | 1024 |
| MICM (mM) | 3.46 | 3.46 | 0.86–55.29 | 13.82 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beier, R.C.; Byrd, J.A.; Caldwell, D.; Andrews, K.; Crippen, T.L.; Anderson, R.C.; Nisbet, D.J. Inhibition and Interactions of Campylobacter jejuni from Broiler Chicken Houses with Organic Acids. Microorganisms 2019, 7, 223. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080223
Beier RC, Byrd JA, Caldwell D, Andrews K, Crippen TL, Anderson RC, Nisbet DJ. Inhibition and Interactions of Campylobacter jejuni from Broiler Chicken Houses with Organic Acids. Microorganisms. 2019; 7(8):223. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080223
Chicago/Turabian StyleBeier, Ross C., J. Allen Byrd, Denise Caldwell, Kathleen Andrews, Tawni L. Crippen, Robin C. Anderson, and David J. Nisbet. 2019. "Inhibition and Interactions of Campylobacter jejuni from Broiler Chicken Houses with Organic Acids" Microorganisms 7, no. 8: 223. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080223