Exploration of Plant-Microbe Interactions for Sustainable Agriculture in CRISPR Era



1
Division of Applied Life Science (BK21 Plus Program), Plant Molecular Biology and Biotechnology Research Center, Gyeongsang National University, Jinju 660-701, Korea
2
Division of Life Science (CK1 Program), Gyeongsang National University, Jinju 660-701, Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Microorganisms 2019, 7(8), 269; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080269
Submission received: 19 July 2019
/
Revised: 8 August 2019
/
Accepted: 14 August 2019
/
Published: 17 August 2019
(This article belongs to the Special Issue Plant Microbial Interactions)
Abstract
:Plants and microbes are co-evolved and interact with each other in nature. Plant-associated microbes, often referred to as plant microbiota, are an integral part of plant life. Depending on the health effects on hosts, plant–microbe (PM) interactions are either beneficial or harmful. The role of microbiota in plant growth promotion (PGP) and protection against various stresses is well known. Recently, our knowledge of community composition of plant microbiome and significant driving factors have significantly improved. So, the use of plant microbiome is a reliable approach for a next green revolution and to meet the global food demand in sustainable and eco-friendly agriculture. An application of the multifaceted PM interactions needs the use of novel tools to know critical genetic and molecular aspects. Recently discovered clustered regularly interspaced short palindromic repeats (CRISPR)/Cas-mediated genome editing (GE) tools are of great interest to explore PM interactions. A systematic understanding of the PM interactions will enable the application of GE tools to enhance the capacity of microbes or plants for agronomic trait improvement. This review focuses on applying GE techniques in plants or associated microbiota for discovering the fundamentals of the PM interactions, disease resistance, PGP activity, and future implications in agriculture.
1. Introduction
In nature, plants and animals continuously interact with numerous microbial species during all the stages of the life cycle. From early times of evolution, humans are exposed to a rich microbial world that extends the human capacity to adapt to a healthy life [1]. Similarly, plants cohabit with microbes, including archaea, protists, bacteria, and fungi, together called microbiota [2]. The beginning of microbial life dated back to the beginning of life (more than 3.5 billion years), suggesting that microbe–microbe interactions have evolved and diversified over time, long before the adaptation of plants to the land life, i.e., before 450 million years [3]. Higher plants and photosynthetic algae assimilated cyanobacterial endosymbionts in the process of evolution, now we know them as chloroplasts or plastids [4]. Thus, the evolutionary history of plant and microbes share common origins, and their survival is interdependent. Consequently, the “plant microbiota” has gained more attention that exists within or nearby surfaces of the plant parts [5].
Profiling of the plant-associated microbiome (genome assemblies of all microbes) is an emerging concept to understand the plant–microbe (PM) interactions. Microbiota extends the plant capacity to acclimatize fluctuating environmental conditions through several mechanisms. Beneficial PM interactions include plant growth promotion (PGP), protection against biotic and abiotic stresses through the priming of plant immune system or induction of plant defence pathways, adaptation to a variable environment, mycorrhizal symbiosis, nutrient uptake, and conversion of the unavailable nutrient forms into plant-accessible form (summarized in Reference [3]). The PM interactions are bidirectional, and microbes also obtain nutrients from the host plants. The trade-off between plant and microbe may develop into distinguishing partnerships depending on its impact on plant health, i.e., mutualistic (beneficial to both the partners, symbiotic), neutral (beneficial to only one partner, commensalistic), or harmful (deleterious to the host plant, pathogenic) [6,7]. These PM interactions are crucial in sustainable agriculture and the environment for food production and health management, respectively.
An investigation of the host plant together with associated microbiome (also called holobiont) suggests the coevolution of plant–microbe, plant–plant, and microbe–microbe interactions [8]. Modern technologies such as next-generation sequencing (NGS), omics approaches (metagenomics, transcriptomics, proteomics, metabolomics), and computational tools enable the understanding of community-level molecular aspects of the PM interactions governing the plant traits. Recently, several reports investigated the diverse aspects of plant microbiota and the influence of host genotype on different facets of the microbiome (Table 1).
Genetic information about the PM interactions is becoming available for several crops and associated microbes. Understanding the molecular aspects of PM interactions at gene level will be a crucial step toward the better use of microbiome in agriculture [14,32]. In this regard, revolutionary techniques such as CRISPR (clustered regularly interspaced short palindromic repeats)-based genome editing (GE) capable of inducing precise genetic modifications [33], are an ideal platform to know the basics of the PM interactions in a fast-forward way and enable precise genetic modifications for higher crop productivity and disease resistance [34]. In this review, we summarize a critical assessment of recent updates about the PM interactions regarding composition, structure, and factors shaping the formation of plant microbiota. We further discuss the CRISPR-based tools and their applications in the beneficial (symbiotic) or harmful (pathogenic) PM interactions towards the development of sustainable agricultural practices. We also elaborate the limitations, regulatory issues, and possible future paths while applying CRISPR-mediated techniques for agricultural purposes in plants or microbes.
2. Composition and Driving Factors of the Plant–Microbe (PM) Interactions
2.1. Composition
Below-ground microbial habitat consists of the rhizosphere (soil close to the root surface), rhizoplane (root surface), and endosphere (root interior). On the other and, above-ground habitat (phyllosphere) of microbes comprises of leaves (phylloplane), stem (caulosphere), flowers (anthosphere), seeds (spermosphere), and fruits (carposphere) (Figure 1). Plants actively recruit microbes from the environment through the soil or air [35]. Some microbes such as endophytes (dwell inside the plant tissues without causing any harm to host plants) follow either horizontal (acquiring from the environment with each new generation) or vertical (transfer from parental seeds) routes of transmission [36]. The NGS-mediated profiling of microbial composition residing below- and above-ground habitats of several plants have provided thorough details about community structure, including agave [9], Arabidopsis [10,11,12,13,21], wild and cultivated barley [14], citrus [15], grapevine [16,17], maize [18,19,20], petunia [21], potato [22], rice [23,24], soybean [25], sugar beet [26], sugarcane [27], tomato [28,29], wheat [25,30], cucumber [30], wild mustard [31], and lettuce [37].
Although several metagenomic studies imply the abundance of the bacterial population in microbiota composition, other microbes such as viruses, protozoa, fungi, oomycetes, nematodes, and algae are also vital contributors (Table 1). Due to the use of diverse sampling methods, primers, and sequencing techniques, it is not possible to compare data from these studies [38]. Nonetheless, all of them suggest that the composition of bacterial communities contain only a few dominant phyla (Actinobacteria, Proteobacteria, Firmicutes, and Bacteroidetes). Several microbial species were found to be common between leaf- and root-associated microbiomes in A. thaliana, agave, grapevine, and wild mustard [9,11,14,17]. The composition of leaf microbiome, but not root, is genetically controlled by host plants and several bacterial species of leaf microbiomes are shared with root microbiomes, suggesting their acquisition from soil [31]. Overall, several reports imply that the structure of microbiota is context-dependent and consists of conserved microbial taxa dwelling in a given plant part across multiple host species and environments. However, more studies are needed to understand the effect of driving factors on microbe–microbe, plant–plant, and PM interactions during plant growth and development.
2.2. Factors Influencing Microbial Communities and PM Interactions
The microbial composition of a plant microbiota is not the result of random selection, and instead, it determined by assembly rules [3,38,39]. Microbial diversity decreases sequentially from bulk soil to rhizosphere. Plant-associated factors promote the preferable growth of a set of microorganisms and inhibit the other. Recent advancement in the mapping of plant microbiota with NGS provides deep insights about biotic (plant-related factors, microbial factors, anthropogenic activities) and abiotic factors (soil properties, environmental factors) affecting the composition and structure of the microbial community (Figure 2) [3,20,38,40]. The profound effects of various factors governing the PM associations have been thoroughly reported in the model plant Arabidopsis, trees, and crops (Table 1).
2.2.1. Biotic Factors
Plant factors include host genotype, the immune system, plant compartment, metabolite secretions, plant age, plant–plant interactions, root morphology, and root exudates. Among them, plant genotype is the major driving factor governing the composition and structure of the microbial community in the rhizosphere [41]. Different plant species vary in their rhizosphere and phyllosphere communities. It is not surprising given that plant genotype decides the properties of root and leaf surface, the type of exudates secreted by roots, chemical signalling pathways, and nutrient quantity plus quality available for microbes [14]. Plant metabolites such as coumarins affect the host microbiota, the assembly of root microbiome, and act as semiochemicals in PM interactions [42]. Microbial species available at specific geographical locations per se also influence the composition of the microbiota and their interactions with plants. The joint action of the plant–plant, microbe–microbe, and PM interactions determines the distinct microbiome assemblies [14]. Symbiotic microbial species are abundant in the niche of rhizosphere and phyllosphere due to positive and selective plant pressure. On the other hand, plant-pathogenic microbes cause a dramatic shift in the population of antagonistic microbes as well as plant immune responses, leading to the control of pathogen [13,43].
Anthropogenic factors such as agricultural practices, including higher dosages of fertilizers, pesticide sprays, cultivation practices pollution, and several other human activities disturb the quality of soil, air, and water, thereby influencing microbial structures and PM associations [12,21,23]. Regarding fertilizer use, the composition of root microbiota of petunia and Arabidopsis varied significantly in response to the phosphorus (P) application and plant species responded differently to low-phosphate conditions [21]. Furthermore, similar effects were observed for leaf microbiota in maize and soybean in response to nitrogen (N) dosages [44,45]. In the future, it will be exciting to study the impact of intensive agricultural practices on changes in PM associations and structure of plant microbiota.
2.2.2. Abiotic Factors
Soil properties have a profound impact on the composition of bacterial and fungal communities in the rhizosphere [22]. The soil is the natural resource of nutrients and hence acts as a microbial seed bank for the rhizosphere community. Soil properties such as soil pH, soil type, macronutrient distribution, soil organic matter, salinity, soil structure, and moisture content drive the microbial community formation [24,25]. Plant species recruit distinct microbial communities in both rhizosphere, rhizoplane (epiphytes, colonize plant surfaces), and endosphere (endophyte, colonize internal plant parts) even if grown in a similar soil environment. On the other hand, certain plant species or genotype recruit the matching group of microorganisms irrespective of environmental and soil conditions, known as the core plant microbiota [46]. Environmental factors also significantly influence the assemblies of phyllosphere microbes that include climate, light, water, ultraviolet (UV) radiation, and geographic location [10,16,18,30]. Generally, plant phenotype is the outcome of interactions between plant genotype, associated microbiota, and environmental factors. Overall, plant microbiota is vertically (through seed, propagation material) or horizontally (through soil, air) acquired and resides on or in the inside of plant tissues where all of the above-discussed factors shape the structure of the microbial community.
3. Role of Plant Microbiota in Sustainable Agriculture
3.1. Beneficial PM Interactions in Agriculture
Plant–microbe interactions regulate the process of soil carbon sequestration by the modulating of the terrestrial carbon cycle [47]. The plant microbiota includes beneficial, neutral, or pathogenic microbial species that decompose the plant residues and dead animals. The beneficial plant microbiota is vital for plant growth, flowering time, and crop yields directly or indirectly [41,48,49]. Furthermore, microbial responses drive the impact of climate changes on agriculture, so there is growing interest to use plant-associated microbiota to mitigate the influence of climate change on sustainable agricultural practices and food production [32]. The consequence of a specific PM interaction is reliant on its effect on plant health, and it may be beneficial under the distinct set of conditions and damaging under the others. Well-known examples of beneficial PM associations include symbiotic interactions of legumes with N-fixing rhizobia and arbuscular mycorrhizal (AM) fungal taxa that helps host plants to access N and P respectively, under nutrient-deficit environments [50,51]. Symbiotic behavior helps PGP microbes to dominate the population of other microbial species. Many PGP bacteria affect plant growth via the production of phytohormones (auxin, cytokinin, gibberellin) and plant-beneficial enzymes (1-aminocyclopropane-1-carboxylate deaminase). Some PM interactions are beneficial under heavy-metal stress through enhanced uptake, and detoxification by either or both the partners, i.e., plant or microbe [52].
3.2. Harmful PM Interactions
Some microbes are harmful to plants causing disease symptoms, for example, Pseudomonas syringae, Erwinia amylovora, Ralstonia solanacearum, Xanthomonas sp., and Xylella fastidiosa. Plant-pathogenic microbes infect the plant tissues through natural openings or wounds for nutrient acquisition and trigger the immune responses [53]. Various factors regulate the outcome of plant–pathogen interaction like population size, the host vulnerability, the climate, and biotic factors like plant microbiota [54]. Several members of plant microbiome known to enhance plant resistance against phytopathogenic microbes are called biocontrol agents. Some non-pathogenic microbes can act as pathogens under some circumstances such as change of host plants [35] or alteration in microbial population size. Therefore, modern tools could be an ideal platform to understand such mysterious behavior of PM associations. In general, precise information of PM interactions at the molecular level is needed.
4. Modern Tools to Explore PM Interactions
Understanding the basic mechanisms of the plant-specific microbiome is a suitable approach for its use in agriculture since the plant-associated microbiota greatly influences the host’s phenotype, as described above. More precisely, the investigation of microbial and plant genes involved in the PM interactions is vital for the future application of plant microbiome. Molecular biology, omics tools (genomics, transcriptomics, proteomics, metabolomics), and NGS technologies have significantly improved our understanding of plant microbiome that includes the PGP microbes as well as phytopathogens [55,56,57,58]. For example, a shift in the composition of the beneficial microbiota community reported in plant–pathogenic interactions [19,29,59]. Although some metagenomic and proteogenomic reports provided assembly level data of augmented functional categories, alternate methods are required to gain functional insights at the gene or protein level.
One of the research themes about PM application in agriculture deals with the use of microbial consortia (a group of species) whereas another research theme involves a precise genetic modification (GM) of either plant or microbe. In the past, the GM methods, together with gene silencing, are widely used to study gene functions or trait improvement. Transgenic technology is a promising approach to accomplish a faster outcome, but the integration of foreign genetic material limits its widespread use due to regulatory issues [60]. In this regard, GE tools are of great interest that allows scientists to edit genomic sequences in a more precise manner without the integration of a foreign gene [33]. Genome editing technology employs the engineered endonucleases to create a double-strand break (DSB) that undergo DNA repair by endogenous mechanisms and generate different types of mutations [61]. DSB repair mechanisms occur through two major pathways (Figure 3A), non-homologous end-joining (NHEJ) and homology-directed repair (HDR). HDR is precise but less common than NHEJ, and applicable in specific donor-dependent gene replacement.
Targeted genetic modifications can be accomplished through several ways, but three meganucleases (or site-specific nucleases (SSNs) or site-directed nucleases (SDN)), are the most commonly used recently that consist of transcription activator-like effector nucleases (TALENs), zinc finger nucleases (ZFNs), and CRISPR/Cas (Cas, CRISPR-associated) system, [62]. Genome editing by ZNFs and TALENs is based on the ability of DNA-binding domains that can specifically recognize almost any target DNA sequence (Figure 3B,C). Therefore, the GE ability of ZNF/TALEN is mainly governed by the DNA-binding affinity and specificity of the assembled zinc-finger and TALE proteins [61]. The CRISPR/Cas system is adventitious compared to ZFNs and TALENs in terms of simple designing, versatility, cost-effective, higher efficiency, multiplexing, and specificity [62]. Since the adaptation of CRISPR/Cas systems for GE in eukaryotes [63,64,65,66], it has emerged as the most popular GE tool that is also described as nature’s toolbox and ‘magic wand’ of genome engineering [67,68]. Thus, primarily CRISPR-mediated GE tools and their applications in PM studies are discussed in the following sections.
5. Components of the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/Cas System
The CRISPR/Cas is a Type II bacterial immune system found in several prokaryotes including bacteria and archaea. Ishino et al. [69] were the first to report some components of CRISPR in Escherichia coli, but the function of these components was not known at that time. A decade ago, the function and mode of action of CRISPR arrays as a programmed immune prokaryotic system against phages was characterized [70,71]. Soon, the components of the CRISPR immune system were reprogrammed for CRISPR-mediated GE. The two main components of CRISPR-based tool include the single gRNA (sgRNA) and Cas endonuclease [63]. The chimeric sgRNA include guide RNA (crRNA) and trans-activating RNA (tracrRNA) (Figure 3D). The key factor for using CRISPR machinery is the presence of protospacer adjacent motif (PAM), a short recognition sequence adjacent to gRNA. The sgRNA designing for the chosen target gene is simple. Several companies offer services for sgRNA synthesis, and plasmid constructs (Cas endonuclease) are available at Addgene repository (https://www.addgene.org/) that make CRISPR technology available for scientists with affordable prices. Also, multiplexing with CRISPR tools is a convenient way to perform editing at multiple targeted loci in a single cell [64]. Cas9 from Streptococcus pyogenes is the first Cas enzyme harnessed for GE, and it is also the most commonly used for development and application of CRISPR-based tools because of robust efficiency and PAM availability [33].
5.1. Cas9 and Cpf1 Orthologs
The gRNA-driven-Cas9 endonuclease scan the target genome for its complementary sequence along with PAM, after recognition, Cas endonuclease produces DSBs [66]. By designing the gRNA next to PAM, this synthetic CRISPR/Cas system can be programmed to target theoretically any desired DNA sequence for genetic modifications. Apart from Cas9, many Cas9 orthologs with the diverse PAM recognition sequences have been discovered. Some of the examples include SpCas9 from Streptococcus pyogenes (5′-NGG-3′) [63], StCas9 from Streptococcus thermophiles (5′-NNAGAAW-3′) [72,73], NmCas9 from Neisseria meningitidis (5′-NNNNGMTT-3′) [74], SaCas9 from Staphylococcus aureus (5′-NNNRRT-3′) [75], FnCas9 from Francisella novivida (5′-NGG-3′) [76], CjCas9 of Campylobacter jejuni (5′-NNNVRYAC-3′) [77], ScCas9 from Streptococcus canis (5′-NNG-3′) [78], and CasX (5′-TTCN-3′) [79], whereas N is any nucleotide, R is A/G, M is A/C, and W is A/T, V is G/C/A, R is A/G, Y is C/T. Among them, native spCas9 is widely used in plants [80]. Another popular Cas9 ortholog is Cpf1. Unlike Cas9, the trans-activating RNA (tracrRNA) is not required for pre-crRNA processing in Cpf1 [81]. Also, Cas9 cleavage produces blunt ends, whereas Cpf1 cutting generate staggered ends (Figure 3D,E). The reprogrammed Cpf1 orthologs for GE include FnCpf1 from Francisella tularensis subsp. novicida U112 (5′-TTV/TTTV/KYTV-3′) [81], LbCpf1 from Lachnospiraceae bacterium ND2006 (5′-TTTV-3′) [81], AsCpf1 from Acidaminococcus sp. BV3L6 (5′-TTTV-3′) [81], and MbCpf1 from Moraxella bovoculi 237 (5′-TTV/TTTV-3′) [82].
5.2. Cas9 and Cpf1 Variants
Two mutant versions of Cas9, catalytically inactive cas9 (dCas9, D10A, and H840A double mutant) [63,78] and nickase Cas9 (nCas9, either D10A or H840A single mutant) [83], have been manipulated for diverse applications. For instance, base editors fused with dCas9 or nCas9 have been developed for specific C-to-T [84] or A-to-G [85] mutations in DNA. Besides, SpCas9 and Cpf1 variants have been engineered either for higher specificity or variable PAM recognition (Figure 4). Some examples of the SpCas9 variants include: SpCas9(D1135E) [64], SpCas9(VQR) [86], SpCas9(EQR) [86], SpCas9(VRER) [86], SpCas9(QQR) [87], eSpCas9(1.0) [88], eSpCas9(1.1) [88], SpCas9-HF1 [89], HeFSpCas9 [90], HypaCas9 [91], evoCas9 [92] xCas9 [93], Cas9-NG [94], HiFiCas9 [95], Sniper Cas9 [96], iSpmacCas9 [95], eHF1-Cas9 [97] and eHypaCas9 [97]. Also, some Cpf1 variants include AsCpf1(RR) [98], AsCpf1(RVR) [98], enAsCpf1 [99], LbCpf1(RR) [98], LbCpf1(RVR) [98], FnCpf1(RR) [82], FnCpf1(RVR) [82], MbCpf1(RR) [82], and MbCpf1(RVR) [82].
These Cas9/Cpf1 variants and orthologs provide alternatives to access different targets in the genome that are otherwise not possible to target with 5′-NGG-3′ or 5′-TTTV-3′ as PAM using native SpCas9 and Cpf1, respectively. Even though higher specificity of Cas9 may not be a major concern in plant or microbe, it is critical for clinical applications to avoid off-target effects [100].
5.3. RNA-Targeting Endonucleases
Some Cas9 cousins from bacterial immune systems also have been redesigned for RNA-editing, such as C2c2 or C2c6 (Cas13a from Leptotrichia shahii or Cas13b from Porphyromonas Prevotella) [101,102]. The Cas13-gRNA system has also been developed for RNA base editing, where catalytically inactive Cas13b fused to the adenosine deaminase domain of ADAR2 (Adenosine deaminases acting on RNA) for programmable A-to-Inosine (G) conversion in transcripts in human [103]. A most recent addition to CRISPR toolbox is the C-to-U RNA editor designed by directed evolution of ADAR2 into a cytidine deaminase [104]. This RNA-editing system is also applied for knockdown of RNA transcripts in plants, for example, in rice (Oryza sativa) protoplasts [105]. Recently, the Cas13-based molecular detection system, described as Specific High-Sensitivity Enzymatic Reporter Unlocking (SHERLOCK), was developed for pathogen identification and genotyping [106,107] as well as for detection of multiple plant genes in a single reaction [108].
6. CRISPR-Based Programmed Tools and Applications
Apart from the use of the CRISPR/Cas system as GE technology, programmable tools have been developed using modified Cas9 versions (i.e., nCas9 and dCas9). These CRISPR-based tools demonstrated its use in single base editing, gene regulation, epigenetic editing, chromatin engineering, imaging, and many more yet to come (Figure 5) [109,110,111]. The next generation CRISPR-based tools expanded beyond DSB-based GE and imparted the capability to these tools to precisely target the DNA region.
The endonuclease capability of native (wild-type) SpCas9 has been explored in basic research to generate knock-out or knock-in mutants to study the gene function [63,64,65,66], and also to improve HR-based donor DNA integration at specific locus [112]. To understand the complex regulatory network, the multiplex CRISPR system is more powerful and allows targeting multiple sites with the desired manner [113]. EvolvR is a novel, targeted mutagenesis tool, designed to incorporate semi-random mutations at a dCas9/gRNA-targeted site in plasmid or prokaryotic genome [114]. The fusion of impaired Cas9 (nCas9 or dCas9) with cytidine deaminase or adenine deaminase was employed by David Liu’s group to generate cytidine base editor (CBE) and adenine base editor (ADE), respectively [84,85]. The CBE and ABE make nucleotide substitutions from C-to-T and A-to-G, which is already applied in plants [115] and microbes [116]. Additionally, mRNA trafficking was visualized in living cells by RNA-targeted Cas9 (RCas9) that could facilitate in vivo RNA tracking of desired transcripts [117].
CRISPR-based screening tools such as nucleic acid detection [106,107], gene tagging [118], barcoding [119], and lineage tracing [120], along with functional-specific genomic library [121] allows screening of the target precisely from a large population. Live-cell imaging CRISPR tools with single or multiple fluorescent proteins are highly efficient to understand the chromatin structure [122], chromatin dynamics [123,124], and topology by manipulating chromatin loops between regulatory genomic regions [125]. The CRISPR/cas9 system has been re-programmed to regulate the gene expression via modulating the transcription, translation, and epigenome.
In the CRISPR activation (CRISPRa) tool, dCas9 was fused with activator (VP64 domain) proteins to activate the transcription [66] that is also effectively applied in plants [126] and bacterial species [127], whereas the dCas9-based CRISPRi (interference) system uses the repressor complex to suppress the gene transcription [126,128]. The Cas13 has been shown to knock-out mRNA of the targeted gene, thereby suppressing the gene function [103]. Furthermore, dCas9-based epigenetic effectors are successfully applied to reactivate silenced genes or transposons through histone modification and catalysis of DNA demethylation in human [129] and plants [130].
Overall, pertaining to expanding the CRISPR toolbox, the CRISPR has emerged as a user-friendly GE tool for a safer and faster alternative to transgenic GM technology and time-consuming traditional breeding methods, respectively.
7. CRISPR-Mediated PM Applications in Agriculture
Several reports have documented the use of CRISPR-based tools in plants with particular emphasis on plant–pathogen interactions to develop disease resistance [34,80,131,132]. Here, we highlight the potential use of CRISPR technology in PM interactions to meet the food demand of the growing population sustainably through discovering the critical mechanisms of the PM interactions, developing disease resistance varieties, and enhancement of PGP activity.
7.1. Understanding the Fundamentals of the PM Interactions
The PM interactions are reliant on the genetics of both the microbiome and the host [133]. Some pathways of the plant immune system evolved to recognize molecules secreted by associated microbiota that serve as signals to trigger the host protection against phytopathogens [134]. Also, plant hormones such as ethylene, jasmonate, and salicylic acid, act as signals that facilitate PM interactions [135]. Especially, understanding the ‘Red Queen’ dynamics (coevolutionary cycles perpetuated due to the conflicts between pathogens and plants) of the pathogenic PM interactions will enhance our knowledge about their vital evolutionary principles (reviewed in Reference [136]).
Standard tools such as 16S ribosomal RNA-sequencing facilitate to distinguish the makeup of a microbial community. Also, multiple omics tools (gen-, transcript-, prote-, and metabol-omics) shed light on the community-level functions by plant-associated genes and pathways. However, these tools cannot determine harmful, neutral, or beneficial interaction with the host plant. Also, omics data typically emphasizes single time points of PM interactions, and thus cannot determine spatiotemporal dynamic interplays [137]. Given the significant role of microbes in plant fitness, identifying essential genes in PM interaction regulating the agronomic trait will help to improve specific plant traits for sustainable agriculture and industries. One of the critical applications of CRISPR-based GE tools is to study the gene functions by genetic modification in plants or microbes. The complete knockdown of the target gene is a unique advantage of CRISPR-based tools compared to partial gene silencing by the RNA interference (RNAi) technique that produces partial phenotype [138]. Overall, using CRISPR-based tools, we can gain more precise genetic information, particularly gene function in PM interaction at the molecular level.
Investigation of model microbes, such as root-rhizobia and phytopathogen Pseudomonas syringae, has provided the mechanistic understanding of genetic factors that causes mutual and pathogenic interactions with hosts, respectively [139,140]. Recently, the CRISPR/Cas system along with single-stranded DNA recombineering was established in the rhizospheric bacterium Pseudomonas putida KT2440 for different genetic modifications, including gene deletion, insertion, replacement, and transcription repression [141,142]. However, mechanistic studies on non-model microbial isolates are necessary to explore gene-level connections in harmful or pathogenic PM interactions. Therefore, GE of non-model microorganisms with robust CRISPR/Cas tools enable the studies to establish links between genes and functions. Further, a recent trend of using biomaterials either in DNA, mRNA, or protein form offers a unique solution for CRISPR/Cas delivery in organisms that cannot be possible by conventional methods [143].
7.2. Plant Disease Resistance
Average global yield losses due to pathogenic PM and plant–insect interactions range from 11% to 30%, mostly in the regions already suffering from food scarcity [144]. So, developing resistant crop varieties against phytopathogens has been a continuous task for agricultural scientists. Classical methods have been applied to introduce genetic modifications for improved disease-resistant plant varieties, such as crossbreeding, natural mutations, hybridization, radiation or chemical mutagenesis, and biological mutagenesis [132]. However, these methods generate several non-targeted modifications, and screening remains a time-consuming and a laborious job. In GM and GE techniques, commonly used modes for phytopathogen resistance include blocking of pathogen entry, alteration of plant defense system, modulation of recessive traits or susceptibility genes (S-genes), activation of dominant resistance genes (R-genes), expression of antimicrobial peptides, and RNAi use [145]. As discussed earlier, transgene insertion by GM technology is viewed as problematic. Also, an integrated approach (use of GM technology along with biocontrol agents, chemical fertilizers, and pesticides) to combat phytopathogens has limitations, for example, several pesticides are not efficient after pathogen evolution, and higher doses may cause harm to humans and the environment [146].
In the last decade, a new set of tools collectively labeled as new plant breeding techniques (NPBTs or NBTs), including CRISPR/Cas, have developed as an alternative (rather complementary) to classical plant breeding and GM tools [34,132]. Examples of CRISPR-mediated genetic modifications of plant or pathogen, including bacteria, fungi, oocytes, and viruses to improve disease resistance summarized in Table 2. The primary step for successful application of GE is to know the molecular details about a target gene from the plant or the pathogen. Whole-genome sequencing of several crop species and associated microbiota is available. The CRISPR-mediated disease resistance can be achieved by targeting the gene either from the host plant or pathogen.
Unlike humans and other vertebrates, plants don’t have an adaptive immune system or mobile immune cells, and are reliant on the inter-connected two-tier innate immune system to tackle pathogenic interactions [147,148]. One branch employs cell surface pattern-recognition receptors (PRRs) to distinguish between “non-self” and “self,” i.e., pathogen-associated molecular patterns (PAMPs) and plant-derived damage-associated molecular patterns (DAMPs), and initiate the PAMP-triggered immunity (PTI) [149]. The other uses resistance (R)-proteins to tackle pathogen-derived effectors to prevent the pathogen entry into the host cell and activate effector-triggered immunity (ETI) [150]. Receptor-like kinases and receptor-like proteins are known to acts as PRRs, whereas R-genes encode intracellular nucleotide-binding leucine-rich repeat receptor (NLRs) proteins. Such essential genes of plant innate immunity involved in the host susceptibility (S-genes) or resistance (R-genes) produced against a pathogenic response (virulence proteins or effectors) are well studied. The GE techniques targeted either of the above candidate plant genes (from PTI or ETI) or pathogen genes to confer resistance against bacteria [151,152,153,154,155,156,157,158], fungi and oocytes [159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177], and DNA/RNA viruses [178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193] (refer to Table 2 for details). Besides, the CRISPR/Cas system is also introduced in some microorganisms for targeted GE that act as biocontrol agents, such as insect-pathogenic fungus Beauveria bassiana, filamentous saprobic fungus Purpureocillium lilacinum controlling plant nematodes, and filamentous fungi like the Trichoderma species [194,195,196].
7.3. Plant Growth Promotion and Nutrient Uptake
Plant-beneficial free-living microbes are known as plant-growth-promoting microbes (PGPM). Among them, the most studied species include N-fixing rhizobia and arbuscular mycorrhiza (AM) fungi. For example, the plant root-associated AM fungi from phylum Glomeromycota inhabit 80%–90% of terrestrial plants and mobilize nutrients from the soil to plant [197]. Similarly, root endophytic fungus Colletotrichum tofieldiae isolated from Arabidopsis exhibits PGP activity and mobilize P to the host plant under phosphate-deficient conditions [198]. While pathogenic PM interactions have been intensely investigated to develop disease resistance in plants, little is known about several PGPM from plant microbiota involved in the beneficial PM interactions. However, the beneficial PM interactions are frequently specific to a species or a cultivar. Plant signaling mechanisms engaged in beneficial effects are very similar to the pathogenic PM interactions. But, it is not yet clear what decides the outcome of particular PM interactions [7]. The beneficial PM interaction is the result of complex processes, like enhancing the nutrient accessibility through iron uptake, N-fixation, and potassium or phosphate solubilization. These also include the activation of plant defense pathways against biotic stresses by signaling molecules, mainly systemic acquired resistance and induced systemic resistance [7].
Some legume root-associated bacteria and archaea have evolved the ability to convert the non-bioavailable (atmospheric) N into available (ammonium) form. This symbiotic plant-rhizobia association involves the trade of food in exchange for fixed-N. A systematic study of this symbiotic interaction about nodule organogenesis may offer ways for engineering non-legume crops to host N-fixing bacteria [199]. In this regard, GE tools are already established in some model legume species, for example, Lotus japonicus [200], Medicago truncatula [201], Glycine max [202] and Vigna unguiculata L.Walp [203]. Moreover, CRISPR protocols for some rhizospheric PGPMs also designed for genetic studies that include Bacillus mycoides and B. subtilis [204]. Overall, expanding the CRISPR toolbox could revolutionize the breeding of food legumes and non-legumes to acquire efficient N-fixing rhizobia and P-solubilizing microbes.
Detailed molecular exploration of beneficial PM interactions with GE will facilitate their field application to non-host species, organic farms, or unfertilized soils for enhanced crop productivity. Also, the use of such GE-modified beneficial microbes in agriculture is the best alternative to agrochemicals, which is notoriously incompetent [205]. Out of field-applied micronutrients and pesticides, up to 95% and 99.9% are wasted and never reach to target [206,207], causing environmental pollution. Therefore, PGPM could be better options to decrease the cost in an eco-friendly way. The application of such genetically engineered microbial inoculants can avoid the rapid decline in introduced microbial population and subsequent benefit to the crops [208]. This also facilitates the plant microbiota-mediated remediation of contaminated soils [56].
7.4. Metabolic Engineering
Secondary metabolites (SMs) are generally described as natural products manufactured by an organism that is not indispensable to support life and growth [209]. The plant and associated microbes produce different SMs, mostly through some chemical pathways. While these SMs are critical in plant or microbial defense pathways, many of them have been utilized as nutrients, medicines, repellants, fragrances, flavors, and coloring agents [209]. Numerous approaches have been adopted for native or heterologous production of SMs in plants [210] and microbes [211,212].
Several plants microbiome studies provide an interesting insight into microbial texa supporting the activity of plant SMs production. Particularly, seed microbiome of medicinal plants such as Salvia miltiorrhiza [213] shows an overlapping set of bacterial and fungal genera with that of maize [214], bean [215], rice [216], and rapeseed [210]. Also, PM-associated microbes share common terpenoid metabolic pathways with the host plant, signifying their potential as a repository of SM-related genes. Furthermore, the SMs production in plants varies depending on geographical location, partly due to altered microbiota suggesting a direct link between plant metabolome and associated microbiome [217]. In this regard, the CRISPR-mediated editing of PM-associated genes involved in the SM pathway provides an innovative and attractive strategy to accomplish higher production of stable and bioactive SMs.
Recently, CRISPR-based metabolic engineering studies in either microbes or plants were performed regarding the basic understanding of SMs pathways or enhanced production of SMs [218,219]. Some studies in microbes include Beauveria bassiana (uridine synthesis) [196], Trichoderma reesei (uridine synthesis) [194], Escherichia coli (flavonoid synthesis) [220], Myceliophthora (cellulase production) [221], Aspergillus niger (galactaric acid production) [222], Aspergillus oryzae (pigment production) [223], Shiraia bambusicola (hypocrellin production) [224], and Aspergillus fumigatus (trypacidin biosynthesis) [225]. Besides, some examples of CRISPR-based metabolic engineering in plants include opium poppy (morphine biosynthesis) [226], tomato (γ-aminobutyric acid, GABA) [227], S. miltiorrhiza (tanshinone biosynthesis) [228], Dendrobium sp. (lignocellulose biosynthesis) [229], Camelina sativa (triacylglycerol synthesis) [230], tobacco (glycan biosynthesis) [231], and S. miltiorrhiza (phenolic acid) [232]. Hence, it will be exciting to use CRISPR technology for editing PM interactions to discover the novel aspects of SMs pathways and modulation of those pathways for better SM productivity.
Overall, the application of CRISPR technology holds immense potential for understanding the fundamental aspects as well as applications of PM interactions in sustainable agriculture (summarized in Table 3 along with potential CRISPR tools) in the future.
8. Limitations and Possible Solutions
Despite its several unprecedented advantages, the CRISPR/Cas technology still has limits while using it in different studies of PM interactions and agriculture. In general, nuclease protein size, nuclease efficiency, and PAM availability can all influence the overall CRISPR outcome, including delivery, off-target effects, and lack of viable targets [233]. As most of the CRISPR-based tools are based on SpCas9, novel Cas9 orthologs are being mined and programmed to overcome the current limitations. Some of the critical issues are discussed in the following text, along with possible solutions and future implications.
8.1. Culture, Species Isolation, and Transformation Protocols
Research exclusively based on culture-dependent experiments have ignored the diversity of plant microbiota. Profiling of plant microbiome by culture-independent tools has transformed our understanding of the diversity and population of microbes at the community level [2,9,12,17]. Also, culturing and isolation of individual microbes are critical steps toward developing transformation, genome sequencing, improved gene, and protein details to design CRISPR-based protocols for further study. Therefore, culture media such as abundance-based synthetic inoculants (resembling in vivo habitats) can be used to recover specific microbes from complex microbiota [234]. A recent innovative approach to isolate single microbial candidates includes the culturing microbiota reliant on host-based media components by adjusting various parameters (reviewed by Sarhan et al. [235]). Tissue culture protocols for major crops are either well established or being developed in recent times. Therefore, plant-based culturonomics tools might provide impetus to grow previously uncultivable plant-associated microbes and ultimately, the application of CRISPR-based tools to study PM interactions. This could also enhance the knowledge about the availability of target genes governing the desired trait in plant/microbe.
8.2. Delivery of CRISPR-Based Tools
One of the major hurdles in CRISPR application is CRISPR/Cas delivery in plant and microbial cells. Agrobacterium-mediated T-DNA transformation is the traditional delivery method being used in plants. Present delivery methods are limited to specific genotypes, species, and tissues. There is an immediate need to design novel methods for CRISPR/Cas delivery in non-model plants and microbes [236]. Non-tissue culture delivery methods will facilitate genotype-independent GE, especially using meristematic or germline plant cells. The CRISPR/Cas components can be delivered as DNA, RNA, and protein.
One of the methods comprises the split of Cas enzyme into two inactive parts that must re-assemble into catalytically active Cas to do editing at the target DNA site [237,238]. This split-Cas9 technology is offered spatiotemporal control of Cas activity, decreased frequency of off-targets using transient methods or induced promoters, and reduced size of plasmid vectors.
Recently, the CRISPR-associated transposon-based method was reprogrammed for specific DNA insertion at target sites in bacteria [239,240]. Such pioneering techniques are a necessity for precise DNA insertion without the need for major endogenous DNA repair pathways from plant or microbe.
Lately, ribonucleoprotein complexes (RNPs) and biomaterials have emerged as promising nanotechnology approaches for the direct delivery because of their tunability, easy use, and higher efficacy that cannot be made possible by other techniques [143]. The RNPs provide better control over activity with less off-target effects compared to continuously produced CRISPR components by DNA methods. Also, novel delivery methods such as shoot meristem microinjection is a viable approach for transformation of recalcitrant plants [241].
Another problem is the delivery of several gRNAs for multiplex GE while using DNA methods. The in vivo production and processing of multiple gRNAs by the endogenous tRNA-processing system or a simple array of multiple gRNAs was achieved in plants and broadens the scope of genome engineering [242,243].
8.3. Transgene-Free Applications
Transgene-free GE is a great advantage of CRISPR-based technology. However, removal of the transgene is difficult from plants that are not propagated by seeds, because backcrossing is not possible. In this regard, delivery via RNP methods that do not include DNA is valuable. It will help to avoid the undesirable DNA footprints in the genome of the host. Already, Cas9 protein-gRNA as RNPs has successfully delivered and been verified to perform GE in plants [244] that could also avoid insertion of foreign DNA. Likewise, improved delivery methods based on geminiviral systems could enhance the HDR-based gene targeting in plants without any foreign DNA integration [245].
8.4. Off-Targets, Biosafety Laws, and Regulations
Transfer of technology from the lab-to-land is a prerequisite for successful implementation in the near future. Most of the studies are proofs of concept and need to validate similar outcomes at the field level. The great power of CRISPR/Cas technology needs to be harnessed with a greater responsibility [143]. One of the major concerns of CRISPR technology is the targeting of undesirable sites in the genome, popularly known as off-target effects [52]. These off-targets can be removed in agriculture by backcrossing the CRISPR-developed variety that allows segregation and removal of possible off-targets [91]. Also, there is a chance of gain-of-function mutations caused by potential off-targets leading to higher crop productivity. In some cases, the off-target effect may be a useful tool for mutagenesis of paralogous and homeobox genes using the same set of gRNAs. Moreover, designing highly efficient and specific gRNAs or modified gRNAs could produce a lower level of off-targets [246].
Regulatory issues of engineered plants or microbes via CRISPR are debatable in different parts of the world, primarily regulating the open-field applications of engineered plants or microbes would be a challenging task. It remains unclear whether GE products will be recognized as GM organisms (GMOs) in several countries even though SSNs introduce indels or substitutions at target sites that naturally occur in evolution process [247]. However, CRISPR-modified organisms might be recognized as GMOs in certain countries, which may need to reconsider by the regulatory authorities based on the consequence and kind of genetic modification included in plants or microbes.
9. Conclusions and Future Directions
Understanding the fundamentals of the PM interactions and their engineering for suitable application in sustainable agriculture is the most appropriate way to meet the food demand in the future. In the past, research about PGP microbes and microbe-mediated plant protection was performed using only a few representative species [58]. Therefore, in recent years, detailed molecular studies about microbe-mediated plant benefits have been conducted to broaden the horizon of PM engineering for agriculture. The CRISPR/Cas technology has enormous potential to help scientists to understand the basics of PM interactions and to develop ideal plant/microbes relevant for agricultural application. Consequently, studying a higher number of plant species, more in-depth sequencing analyses of the plant microbiome, and meta-transcriptomic data are supplementary to understand the community-level molecular mechanisms under field conditions. Identification of individual plant or microbial candidate genes governing agronomic traits will facilitate CRISPR-based applications in sustainable agricultural practices. Essential questions to be addressed are—which plant genes allow crops to shape the rhizosphere microbiota? What are the effects of microbes on the host plant? How do plants and microbes communicate with each other? These questions will establish the direct link between an agronomic trait and gene of the plant or microbe, allowing the design of synthetic microbial communities for higher crop productivity.
Author Contributions
R.M.S. and J.-Y.K. conceived the study. R.M.S. and D.P. performed the analyses. R.M.S., D.P. and J.-Y.K. wrote and edited the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (Grant NRF 2017R1A4A1015515) and by the Next-Generation BioGreen 21 Program (SSAC, Grant PJ01322601), Rural Development Administration (RDA), Republic of Korea. R.M.S. and D.P. were supported by Brain Korea 21 Plus program (BK21+).
Conflicts of Interest
The authors declare no conflict of interest that might be perceived as affecting the objectivity of this review.
References
- Kirjavainen, P.V.; Karvonen, A.M.; Adams, R.I.; Täubel, M.; Roponen, M.; Tuoresmäki, P.; Loss, G.; Jayaprakash, B.; Depner, M.; Ege, M.J.; et al. Farm-like indoor microbiota in non-farm homes protects children from asthma development. Nat. Med. 2019, 25, 1089–1095. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Boil. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J. The endosymbiotic origin, diversification and fate of plastids. Philos. Trans. R. Soc. B Boil. Sci. 2010, 365, 729–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waghunde, R.R.; Shelake, R.M.; Shinde, M.S.; Hayashi, H. Endophyte Microbes: A Weapon for Plant Health Management. In Microorganisms for Green Revolution; Springer: Singapore, 2017; pp. 303–325. [Google Scholar]
- Thrall, P.H.; Hochberg, M.E.; Burdon, J.J.; Bever, J.D. Coevolution of symbiotic mutualists and parasites in a community context. Trends Ecol. Evol. 2007, 22, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.A.; Rothballer, M.; Chowdhury, S.P.; Nussbaumer, T.; Gutjahr, C.; Falter-Braun, P. Systems biology of plant-microbiome interactions. Mol. Plant 2019, 12, 804–821. [Google Scholar] [CrossRef]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef]
- Coleman-Derr, D.; Desgarennes, D.; Fonseca-Garcia, C.; Gross, S.; Clingenpeel, S.; Woyke, T.; North, G.; Visel, A.; Partida-Martinez, L.P.; Tringe, S.G. Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 2016, 209, 798–811. [Google Scholar] [CrossRef]
- Schlaeppi, K.; Dombrowski, N.; Oter, R.G.; Ver Loren van Themaat, E.; Schulze-Lefert, P. Quantitative divergence of the bacterial root microbiota in Arabidopsis thaliana relatives. Proc. Natl. Acad. Sci. USA 2014, 111, 585–592. [Google Scholar] [CrossRef]
- Bai, Y.; Müller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Münch, P.C.; Spaepen, S.; Remus-Emsermann, M.; et al. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 2015, 528, 364–369. [Google Scholar] [CrossRef]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; De Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D.; et al. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Durán, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 2018, 175, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and Function of the Bacterial Root Microbiota in Wild and Domesticated Barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, Y.; Zhang, P.; Trivedi, P.; Riera, N.; Wang, Y.; Liu, X.; Fan, G.; Tang, J.; Coletta-Filho, H.D.; et al. The structure and function of the global citrus rhizosphere microbiome. Nat. Commun. 2018, 9, 4894. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [PubMed]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S.; et al. The Soil Microbiome Influences Grapevine-Associated Microbiota. mBio 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [Green Version]
- Niu, B.; Paulson, J.N.; Zheng, X.; Kolter, R. Simplified and representative bacterial community of maize roots. Proc. Natl. Acad. Sci. USA 2017, 114, E2450–E2459. [Google Scholar] [CrossRef] [Green Version]
- Walters, W.A.; Jin, Z.; Youngblut, N.; Wallace, J.G.; Sutter, J.; Zhang, W.; González-Peña, A.; Peiffer, J.; Koren, O.; Shi, Q.; et al. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proc. Natl. Acad. Sci. USA 2018, 115, 7368–7373. [Google Scholar] [CrossRef] [Green Version]
- Bodenhausen, N.; Somerville, V.; Desirò, A.; Walser, J.-C.; Borghi, L.; Van Der Heijden, M.G.A.; Schlaeppi, K. Petunia- and Arabidopsis-Specific Root Microbiota Responses to Phosphate Supplementation. Phytobiomes J. 2019, 3, 112–124. [Google Scholar] [CrossRef]
- Inceoğlu, Ö.; Abu Al-Soud, W.; Salles, J.F.; Semenov, A.V.; Van Elsas, J.D. Comparative Analysis of Bacterial Communities in a Potato Field as Determined by Pyrosequencing. PLoS ONE 2011, 6, e23321. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breidenbach, B.; Pump, J.; Dumont, M.G. Microbial Community Structure in the Rhizosphere of Rice Plants. Front. Microbiol. 2016, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Rascovan, N.; Carbonetto, B.; Perrig, D.; Díaz, M.; Canciani, W.; Abalo, M.; Alloati, J.; González-Anta, G.; Vazquez, M.P. Integrated analysis of root microbiomes of soybean and wheat from agricultural fields. Sci. Rep. 2016, 6, 28084. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Kruijt, M.; De Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.M.; Piceno, Y.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the Rhizosphere Microbiome for Disease-Suppressive Bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.S.C.; Okura, V.K.; Armanhi, J.S.L.; Jorrín, B.; Lozano, N.; Da Silva, M.J.; González-Guerrero, M.; De Araújo, L.M.; Verza, N.C.; Bagheri, H.C.; et al. Unlocking the bacterial and fungal communities assemblages of sugarcane microbiome. Sci. Rep. 2016, 6, 28774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottesen, A.R.; Peña, A.G.; White, J.R.; Pettengill, J.B.; Li, C.; Allard, S.; Rideout, S.; Allard, M.; Hill, T.; Evans, P.; et al. Baseline survey of the anatomical microbial ecology of an important food plant: Solanum lycopersicum (tomato). BMC Microbiol. 2013, 13, 114. [Google Scholar] [CrossRef]
- Kwak, M.-J.; Kong, H.G.; Choi, K.; Kwon, S.-K.; Song, J.Y.; Lee, J.; Lee, P.A.; Choi, S.Y.; Seo, M.; Lee, H.J.; et al. Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat. Biotechnol. 2018, 36, 1100–1109. [Google Scholar] [CrossRef]
- Ofek-Lalzar, M.; Sela, N.; Goldman-Voronov, M.; Green, S.J.; Hadar, Y.; Minz, D. Niche and host-associated functional signatures of the root surface microbiome. Nat. Commun. 2014, 5, 4950. [Google Scholar] [CrossRef]
- Wagner, M.R.; Lundberg, D.S.; Del Rio, T.G.; Tringe, S.G.; Dangl, J.L.; Mitchell-Olds, T. Host genotype and age shape the leaf and root microbiomes of a wild perennial plant. Nat. Commun. 2016, 7, 12151. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Genet. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- Knott, G.J.; Doudna, J.A. CRISPR-Cas guides the future of genetic engineering. Science 2018, 361, 866–869. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, S.S.-E.-A.; Mukhtar, M.S.; Mansoor, S. Genome Editing: Targeting Susceptibility Genes for Plant Disease Resistance. Trends Biotechnol. 2018, 36, 898–906. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.C.; Guzmán, J.P.S.; Shay, J.E. Transmission of Bacterial Endophytes. Microorganisms 2017, 5, 70. [Google Scholar] [CrossRef]
- Cardinale, M.; Grube, M.; Erlacher, A.; Quehenberger, J.; Berg, G. Bacterial networks and co-occurrence relationships in the lettuce root microbiota. Environ. Microbiol. 2015, 17, 239–252. [Google Scholar] [CrossRef]
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The Plant Microbiota: Systems-Level Insights and Perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Genet. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Stringlis, I.A.; De Jonge, R.; Pieterse, C.M.J.; Pieterse, C.M.J. The Age of Coumarins in Plant-Microbe Interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef]
- Chapelle, E.; Mendes, R.; Bakker, P.A.H.; Raaijmakers, J.M. Fungal invasion of the rhizosphere microbiome. ISME J. 2016, 10, 265–268. [Google Scholar] [CrossRef]
- Ikeda, S.; Anda, M.; Inaba, S.; Eda, S.; Sato, S.; Sasaki, K.; Tabata, S.; Mitsui, H.; Sato, T.; Shinano, T.; et al. Autoregulation of Nodulation Interferes with Impacts of Nitrogen Fertilization Levels on the Leaf-Associated Bacterial Community in Soybeans. Appl. Environ. Microbiol. 2011, 77, 1973–1980. [Google Scholar] [CrossRef] [Green Version]
- Manching, H.C.; Balint-Kurti, P.J.; Stapleton, A.E. Southern leaf blight disease severity is correlated with decreased maize leaf epiphytic bacterial species richness and the phyllosphere bacterial diversity decline is enhanced by nitrogen fertilization. Front. Plant Sci. 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushio, M.; Nakaoka, S.; Onoda, Y.; et al. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef]
- Shelake, R.M.; Waghunde, R.R.; Verma, P.P.; Singh, C.; Kim, J.-Y. Carbon Sequestration for Soil Fertility Management: Microbiological Perspective. In Soil Fertility Management for Sustainable Development; Springer: Singapore, 2019; pp. 25–42. [Google Scholar]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef]
- Berg, M.; Koskella, B. Nutrient- and Dose-Dependent Microbiome-Mediated Protection against a Plant Pathogen. Curr. Boil. 2018, 28, 2487–2492. [Google Scholar] [CrossRef]
- Cao, Y.; Halane, M.K.; Gassmann, W.; Stacey, G. The Role of Plant Innate Immunity in the Legume-Rhizobium Symbiosis. Annu. Rev. Plant Boil. 2017, 68, 535–561. [Google Scholar] [CrossRef]
- Udvardi, M.; Poole, P.S. Transport and Metabolism in Legume-Rhizobia Symbioses. Annu. Rev. Plant Boil. 2013, 64, 781–805. [Google Scholar] [CrossRef] [Green Version]
- Shelake, R.M.; Waghunde, R.R.; Morita, E.H.; Hayashi, H. Plant-Microbe-Metal Interactions: Basics, Recent Advances, and Future Trends. In Plant Microbiome: Stress Response; Springer: Singapore, 2018; pp. 283–305. ISBN 9789811055140. [Google Scholar]
- Zoledowska, S.; Presta, L.; Fondi, M.; Decorosi, F.; Giovannetti, L.; Mengoni, A.; Lojkowska, E. Metabolic Modeling of Pectobacterium parmentieri SCC3193 Provides Insights into Metabolic Pathways of Plant Pathogenic Bacteria. Microorganisms 2019, 7, 101. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Vescio, K.; Mitter, B.; Trognitz, F.; Ma, L.-J.; Sessitsch, A. Ecology and Genomic Insights into Plant-Pathogenic and Plant-Nonpathogenic Endophytes. Annu. Rev. Phytopathol. 2017, 55, 61–83. [Google Scholar] [CrossRef]
- Berg, G.; Zachow, C.; Müller, H.; Philipps, J.; Tilcher, R. Next-Generation Bio-Products Sowing the Seeds of Success for Sustainable Agriculture. Agronomy 2013, 3, 648–656. [Google Scholar] [CrossRef]
- Basu, S.; Rabara, R.C.; Negi, S.; Shukla, P. Engineering PGPMOs through Gene Editing and Systems Biology: A Solution for Phytoremediation? Trends Biotechnol. 2018, 36, 499–510. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. The soil microbiome—From metagenomics to metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef]
- Vorholt, J.A.; Vogel, C.; Carlström, C.I.; Müller, D.B. Establishing Causality: Opportunities of Synthetic Communities for Plant Microbiome Research. Cell Host Microbe 2017, 22, 142–155. [Google Scholar] [CrossRef]
- Fürnkranz, M.; Lukesch, B.; Müller, H.; Huss, H.; Grube, M.; Berg, G. Microbial Diversity Inside Pumpkins: Microhabitat-Specific Communities Display a High Antagonistic Potential Against Phytopathogens. Microb. Ecol. 2012, 63, 418–428. [Google Scholar] [CrossRef]
- Muñoz, I.V.; Sarrocco, S.; Malfatti, L.; Baroncelli, R.; Vannacci, G. CRISPR-Cas for Fungal Genome Editing: A New Tool for the Management of Plant Diseases. Front. Plant Sci. 2019, 10, 1–5. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef]
- Brandt, K.; Barrangou, R. Applications of CRISPR Technologies across the Food Supply Chain. Annu. Rev. Food Sci. Technol. 2019, 10, 133–150. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat. Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Güell, M.; Dicarlo, J.E.; Norville, J.E.; Church, G.M. RNA-Guided Human Genome Engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.V.; Nuñez, J.K.; Doudna, J.A. Biology and Applications of CRISPR Systems: Harnessing Nature’s Toolbox for Genome Engineering. Cell 2016, 164, 29–44. [Google Scholar] [CrossRef]
- Sakuma, T.; Yamamoto, T. Magic wands of CRISPR—Lots of choices for gene knock-in. Cell Boil. Toxicol. 2017, 33, 501–505. [Google Scholar] [CrossRef]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakatura, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isoenzyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small CRISPR RNAs Guide Antiviral Defense in Prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [Green Version]
- Karvelis, T.; Gasiunas, G.; Miksys, A.; Barrangou, R.; Horvath, P.; Siksnys, V. crRNA and tracrRNA guide Cas9-mediated DNA interference in Streptococcus thermophilus. RNA Boil. 2013, 10, 841–851. [Google Scholar] [CrossRef]
- Garneau, J.E.; Dupuis, M.-È.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Fremaux, C.; Horvath, P.; Magadán, A.H.; Moineau, S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef]
- Hou, Z.; Zhang, Y.; Propson, N.E.; Howden, S.E.; Chu, L.-F.; Sontheimer, E.J.; Thomson, J.A. Efficient genome engineering in human pluripotent stem cells using Cas9 from Neisseria meningitidis. Proc. Natl. Acad. Sci. USA 2013, 110, 15644–15649. [Google Scholar] [CrossRef]
- Ran, F.A.; Cong, L.; Yan, W.X.; Scott, D.A.; Gootenberg, J.S.; Kriz, A.J.; Zetsche, B.; Shalem, O.; Wu, X.; Makarova, K.S.; et al. In vivo genome editing using Staphylococcus aureus Cas9. Nature 2015, 520, 186–191. [Google Scholar] [CrossRef]
- Price, A.A.; Sampson, T.R.; Ratner, H.K.; Grakoui, A.; Weiss, D.S. Cas9-mediated targeting of viral RNA in eukaryotic cells. Proc. Natl. Acad. Sci. USA 2015, 112, 6164–6169. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Koo, T.; Park, S.W.; Kim, D.; Kim, K.; Cho, H.-Y.; Song, D.W.; Lee, K.J.; Jung, M.H.; Kim, S.; et al. In vivo genome editing with a small Cas9 orthologue derived from Campylobacter jejuni. Nat. Commun. 2017, 8, 14500. [Google Scholar] [CrossRef]
- Chatterjee, P.; Jakimo, N.; Jacobson, J.M. Minimal PAM specificity of a highly similar SpCas9 ortholog. Sci. Adv. 2018, 4, eaau0766. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-J.; Orlova, N.; Oakes, B.L.; Ma, E.; Spinner, H.B.; Baney, K.L.M.; Chuck, J.; Tan, D.; Knott, G.J.; Harrington, L.B.; et al. CasX enzymes comprise a distinct family of RNA-guided genome editors. Nature 2019, 566, 218–223. [Google Scholar] [CrossRef]
- Yin, K.; Gao, C.; Qiu, J.-L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 17107. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A.; et al. Cpf1 is a single RNA-guided endonuclease of a Class 2 CRISPR-Cas system. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef]
- Tóth, E.; Czene, B.C.; Kulcsár, P.I.; Krausz, S.L.; Tálas, A.; Nyeste, A.; Varga, É.; Huszár, K.; Weinhardt, N.; Ligeti, Z.; et al. Mb- and FnCpf1 nucleases are active in mammalian cells: Activities and PAM preferences of four wild-type Cpf1 nucleases and of their altered PAM specificity variants. Nucleic Acids Res. 2018, 46, 10272–10285. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Lin, C.-Y.; Gootenberg, J.S.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Kleinstiver, B.P.; Prew, M.S.; Tsai, S.Q.; Topkar, V.V.; Nguyen, N.T.; Zheng, Z.; Gonzales, A.P.W.; Li, Z.; Peterson, R.T.; Yeh, J.-R.J.; et al. Engineered CRISPR-Cas9 nucleases with altered PAM specificities. Nature 2015, 523, 481–485. [Google Scholar] [CrossRef] [Green Version]
- Anders, C.; Bargsten, K.; Jinek, M. Structural plasticity of PAM recognition by engineered variants of the RNA-guided endonuclease Cas9. Mol. Cell 2016, 61, 895–902. [Google Scholar] [CrossRef]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Rationally engineered Cas9 nucleases with improved specificity. Science 2016, 351, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Kleinstiver, B.P.; Pattanayak, V.; Prew, M.S.; Tsai, S.Q.; Nguyen, N.T.; Joung, J.K. 731. High-Fidelity CRISPR-Cas9 Nucleases with No Detectable Genome-Wide Off-Target Effects. Mol. Ther. 2016, 24, S288. [Google Scholar] [CrossRef]
- Kulcsár, P.I.; Tálas, A.; Huszár, K.; Ligeti, Z.; Tóth, E.; Weinhardt, N.; Fodor, E.; Welker, E. Crossing enhanced and high fidelity SpCas9 nucleases to optimize specificity and cleavage. Genome Boil. 2017, 18, 190. [Google Scholar] [CrossRef]
- Chen, J.S.; Dagdas, Y.S.; Kleinstiver, B.P.; Welch, M.M.; Sousa, A.A.; Harrington, L.B.; Sternberg, S.H.; Joung, J.K.; Yildiz, A.; Doudna, J.A. Enhanced proofreading governs CRISPR-Cas9 targeting accuracy. Nature 2017, 550, 407–410. [Google Scholar] [CrossRef]
- Casini, A.; Olivieri, M.; Petris, G.; Montagna, C.; Reginato, G.; Maule, G.; Lorenzin, F.; Prandi, D.; Romanel, A.; Demichelis, F.; et al. A highly specific SpCas9 variant is identified by in vivo screening in yeast. Nat. Biotechnol. 2018, 36, 265–271. [Google Scholar] [CrossRef]
- Hu, J.H.; Miller, S.M.; Geurts, M.H.; Tang, W.; Chen, L.; Sun, N.; Zeina, C.M.; Gao, X.; Rees, H.A.; Lin, Z.; et al. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 2018, 556, 57–63. [Google Scholar] [CrossRef]
- Nishimasu, H.; Shi, X.; Ishiguro, S.; Gao, L.; Hirano, S.; Okazaki, S.; Noda, T.; Abudayyeh, O.O.; Gootenberg, J.S.; Mori, H.; et al. Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science 2018, 361, 1259–1262. [Google Scholar] [CrossRef]
- Vakulskas, C.A.; Dever, D.P.; Rettig, G.R.; Turk, R.; Jacobi, A.M.; Collingwood, M.A.; Bode, N.M.; McNeill, M.S.; Yan, S.; Camarena, J.; et al. A high-fidelity Cas9 mutant delivered as a ribonucleoprotein complex enables efficient gene editing in human hematopoietic stem and progenitor cells. Nat. Med. 2018, 24, 1216–1224. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Jeong, E.; Lee, J.; Jung, M.; Shin, E.; Kim, Y.-H.; Lee, K.; Jung, I.; Kim, D.; Kim, S.; et al. Directed evolution of CRISPR-Cas9 to increase its specificity. Nat. Commun. 2018, 9, 3048. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Yan, Y.; Zhang, Y.; Gao, C. Genotyping genome-edited mutations in plants using CRISPR ribonucleoprotein complexes. Plant Biotechnol. J. 2018, 16, 2053–2062. [Google Scholar] [CrossRef]
- Gao, L.; Cox, D.B.T.; Yan, W.X.; Manteiga, J.C.; Schneider, M.W.; Yamano, T.; Nishimasu, H.; Nureki, O.; Crosetto, N.; Zhang, F. Engineered Cpf1 variants with altered PAM specificities. Nat. Biotechnol. 2017, 35, 789–792. [Google Scholar] [CrossRef] [Green Version]
- Kleinstiver, B.P.; Sousa, A.A.; Walton, R.T.; Tak, Y.E.; Hsu, J.Y.; Clement, K.; Welch, M.M.; Horng, J.E.; Malagon-Lopez, J.; Scarfò, I.; et al. Engineered CRISPR-Cas12a variants with increased activities and improved targeting ranges for gene, epigenetic and base editing. Nat. Biotechnol. 2019, 37, 276–282. [Google Scholar] [CrossRef]
- Kadam, U.S.; Shelake, R.M.; Chavhan, R.L.; Suprasanna, P. Concerns regarding ’off-target’ activity of genome editing endonucleases. Plant Physiol. Biochem. 2018, 131, 22–30. [Google Scholar] [CrossRef]
- Smargon, A.A.; Cox, D.B.; Pyzocha, N.K.; Zheng, K.; Slaymaker, I.M.; Gootenberg, J.S.; Abudayyeh, O.A.; Essletzbichler, P.; Shmakov, S.; Makarova, K.S.; et al. Cas13b is a Type VI-B CRISPR-associated RNA-Guided RNAse differentially regulated by accessory proteins Csx27 and Csx28. Mol. Cell 2017, 65, 618–630. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA editing with CRISPR-Cas13. Science 2017, 358, 1019–1027. [Google Scholar] [CrossRef] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Franklin, B.; Koob, J.; Kellner, M.J.; Ladha, A.; Joung, J.; Kirchgatterer, P.; Cox, D.B.T.; Zhang, F. A cytosine deaminase for programmable single-base RNA editing. Science 2019, 538, eaax7063. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR-Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Gootenberg, J.S.; Abudayyeh, O.O.; Kellner, M.J.; Joung, J.; Collins, J.J.; Zhang, F. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science 2018, 360, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Kellner, M.J.; Zhang, F. Nucleic Acid Detection of Plant Genes Using CRISPR-Cas13. CRISPR J. 2019, 2, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Qi, L.S. A CRISPR–dCas Toolbox for Genetic Engineering and Synthetic Biology. J. Mol. Boil. 2019, 431, 34–47. [Google Scholar] [CrossRef]
- Simeonov, D.R.; Marson, A. CRISPR-Based Tools in Immunity. Annu. Rev. Immunol. 2019, 37, 571–597. [Google Scholar] [CrossRef]
- Adli, M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018, 9, 1911. [Google Scholar] [CrossRef]
- Bassett, A.R.; Tibbit, C.; Ponting, C.P.; Liu, J.-L. Mutagenesis and homologous recombination in Drosophila cell lines using CRISPR/Cas9. Biol. Open 2014, 3, 42–49. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene Editing in the Escherichia coli Genome via the CRISPR-Cas9 System. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef]
- Halperin, S.O.; Tou, C.J.; Wong, E.B.; Modavi, C.; Schaffer, D.V.; Dueber, J.E. CRISPR-guided DNA polymerases enable diversification of all nucleotides in a tunable window. Nature 2018, 560, 248–252. [Google Scholar] [CrossRef]
- Li, J.; Sun, Y.; Du, J.; Zhao, Y.; Xia, L. Generation of Targeted Point Mutations in Rice by a Modified CRISPR/Cas9 System. Mol. Plant 2017, 10, 526–529. [Google Scholar] [CrossRef] [Green Version]
- Zheng, K.; Wang, Y.; Li, N.; Jiang, F.-F.; Wu, C.-X.; Liu, F.; Chen, H.-C.; Liu, Z.-F. Highly efficient base editing in bacteria using a Cas9-cytidine deaminase fusion. Commun. Boil. 2018, 1, 32. [Google Scholar] [CrossRef]
- Nelles, D.A.; Fang, M.Y.; O’Connell, M.R.; Xu, J.L.; Markmiller, S.J.; Doudna, J.A.; Yeo, G.W. Programmable RNA tracking in Live Cells with CRISPR/Cas9. Cell 2016, 165, 488–496. [Google Scholar] [CrossRef]
- Roberts, B.; Haupt, A.; Tucker, A.; Grancharova, T.; Arakaki, J.; Fuqua, M.A.; Nelson, A.; Hookway, C.; Ludmann, S.A.; Mueller, I.A.; et al. Systematic gene tagging using CRISPR/Cas9 in human stem cells to illuminate cell organization. Mol. Boil. Cell 2017, 28, 2854–2874. [Google Scholar] [CrossRef]
- Kalhor, R.; Kalhor, K.; Mejia, L.; Leeper, K.; Graveline, A.; Mali, P.; Church, G.M. Developmental barcoding of whole mouse via homing CRISPR. Science 2018, 361, eaat9804. [Google Scholar] [CrossRef]
- McKenna, A.; Findlay, G.M.; Gagnon, J.A.; Horwitz, M.S.; Schier, A.F.; Shendure, J. Whole organism lineage tracing by combinatorial and cumulative genome editing. Science 2016, 353, aaf7907. [Google Scholar] [CrossRef]
- Miles, L.A.; Garippa, R.J.; Poirier, J.T. Design, execution, and analysis of pooled in vitro CRISPR/Cas9 screens. FEBS J. 2016, 283, 3170–3180. [Google Scholar] [CrossRef]
- Qin, P.; Parlak, M.; Kuscu, C.; Bandaria, J.; Mir, M.; Szlachta, K.; Singh, R.; Darzacq, X.; Yildiz, A.; Adli, M. Live cell imaging of low- and non-repetitive chromosome loci using CRISPR-Cas9. Nat. Commun. 2017, 8, 14725. [Google Scholar] [CrossRef]
- Dreissig, S.; Schiml, S.; Schindele, P.; Weiss, O.; Rutten, T.; Schubert, V.; Gladilin, E.; Mette, M.F.; Puchta, H.; Houben, A. Live-cell CRISPR imaging in plants reveals dynamic telomere movements. Plant J. 2017, 91, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Acar, M. Live-Cell Imaging of Chromatin Condensation Dynamics by CRISPR. iScience 2018, 4, 216–235. [Google Scholar] [CrossRef]
- Morgan, S.L.; Mariano, N.C.; Bermudez, A.; Arruda, N.L.; Wu, F.; Luo, Y.; Shankar, G.; Jia, L.; Chen, H.; Hu, J.-F.; et al. Manipulation of nuclear architecture through CRISPR-mediated chromosomal looping. Nat. Commun. 2017, 8, 15993. [Google Scholar] [CrossRef] [Green Version]
- Piatek, A.; Ali, Z.; Baazim, H.; Li, L.; Abulfaraj, A.; Al-Shareef, S.; Aouida, M.; Mahfouz, M.M. RNA-guided transcriptional regulation in planta via synthetic dCas9-based transcription factors. Plant Biotechnol. J. 2015, 13, 578–589. [Google Scholar] [CrossRef]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef]
- Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell 2013, 154, 442. [Google Scholar] [CrossRef]
- Hilton, I.B.; D’Ippolito, A.M.; Vockley, C.M.; Thakore, P.I.; Crawford, G.E.; Reddy, T.E.; Gersbach, C.A. Epigenome editing by a CRISPR/Cas9-based acetyltransferase activates genes from promoters and enhancers. Nat. Biotechnol. 2015, 33, 510–517. [Google Scholar] [CrossRef]
- Gallego-Bartolomé, J.; Gardiner, J.; Liu, W.; Papikian, A.; Ghoshal, B.; Kuo, H.Y.; Zhao, J.M.-C.; Segal, D.J.; Jacobsen, S.E. Targeted DNA demethylation of the Arabidopsis genome using the human TET1 catalytic domain. Proc. Natl. Acad. Sci. USA 2018, 115, E2125–E2134. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Boil. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Langner, T.; Kamoun, S.; Belhaj, K. CRISPR Crops: Plant Genome Editing Toward Disease Resistance. Annu. Rev. Phytopathol. 2018, 56, 479–512. [Google Scholar] [CrossRef]
- Levy, A.; Conway, J.M.; Dangl, J.L.; Woyke, T. Elucidating Bacterial Gene Functions in the Plant Microbiome. Cell Host Microbe 2018, 24, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Van Der Meer, J.W. Trained Immunity: An Ancient Way of Remembering. Cell Host Microbe 2017, 21, 297–300. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Han, G.-Z. Origin and evolution of the plant immune system. New Phytol. 2019, 222, 70–83. [Google Scholar] [CrossRef]
- Pinski, A.; Betekhtin, A.; Hupert-Kocurek, K.; Mur, L.A.J.; Hasterok, R. Defining the Genetic Basis of Plant–Endophytic Bacteria Interactions. Int. J. Mol. Sci. 2019, 20, 1947. [Google Scholar] [CrossRef]
- Bisht, D.S.; Bhatia, V.; Bhattacharya, R. Improving plant-resistance to insect-pests and pathogens: The new opportunities through targeted genome editing. Semin. Cell Dev. Boil. 2019. [Google Scholar] [CrossRef]
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Genet. 2018, 16, 316–328. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Sun, J.; Wang, Q.; Jiang, Y.; Wen, Z.; Yang, L.; Wu, J.; Yang, S. Genome editing and transcriptional repression in Pseudomonas putida KT2440 via the type II CRISPR system. Microb. Cell Fact. 2018, 17, 41. [Google Scholar] [CrossRef]
- Aparicio, T.; de Lorenzo, V.; Martínez-García, E. CRISPR/Cas9-Based Counterselection Boosts Recombineering Efficiency in Pseudomonas putida. Biotechnol. J. 2018, 13, 1–10. [Google Scholar] [CrossRef]
- Eoh, J.; Gu, L. Biomaterials as vectors for the delivery of CRISPR–Cas9. Biomater. Sci. 2019, 7, 1240–1261. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Van Esse, H.P.; Reuber, L.; van der Does, D. GM approaches to improve disease resistance in crops. New Phytol. 2019. [Google Scholar] [CrossRef]
- Berger, S.; El Chazli, Y.; Babu, A.F.; Coste, A.T. Azole Resistance in Aspergillus fumigatus: A Consequence of Antifungal Use in Agriculture? Front. Microbiol. 2017, 8, 1024. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Vance, R.E.; Dangl, J.L. Intracellular innate immune surveillance devices in plants and animals. Science 2016, 354, aaf6395. [Google Scholar] [CrossRef] [Green Version]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant Boil. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Upson, J.L.; Zess, E.K.; Białas, A.; Wu, C.-H.; Kamoun, S. The coming of age of EvoMPMI: Evolutionary molecular plant–microbe interactions across multiple timescales. Curr. Opin. Plant Boil. 2018, 44, 108–116. [Google Scholar] [CrossRef]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-efficiency TALEN-based gene editing produces disease-resistant rice. Nat. Biotechnol. 2012, 30, 390–392. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, Z.; Long, J.; Sosso, D.; Liu, B.; Eom, J.-S.; Huang, S.; Liu, S.; Cruz, C.V.; Frommer, W.B.; et al. Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice. Plant J. 2015, 82, 632–643. [Google Scholar] [CrossRef]
- Thomazella, D.P.D.T.; Brail, Q.; Dahlbeck, D.; Staskawicz, B.J. CRISPR-Cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. bioRxiv 2016. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Kanchiswamy, C.N. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1473. [Google Scholar] [CrossRef]
- Ortigosa, A.; Gimenez-Ibanez, S.; Leonhardt, N.; Solano, R. Design of a bacterial speck resistant tomato by CRISPR/Cas9-mediated editing of SlJAZ2. Plant Biotechnol. J. 2019, 17, 665–673. [Google Scholar] [CrossRef]
- Xie, K.; Yang, Y. RNA-Guided Genome Editing in Plants Using a CRISPR-Cas System. Mol. Plant 2013, 6, 1975–1983. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.-L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Foster, A.J.; Martin-Urdiroz, M.; Yan, X.; Wright, H.S.; Soanes, D.M.; Talbot, N.J. CRISPR-Cas9 ribonucleoprotein-mediated co-editing and counterselection in the rice blast fungus. Sci. Rep. 2018, 8, 14355. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Zhang, Y.; Yu, P.-L.; Pan, H.; Rollins, J.A. Introduction of Large Sequence Inserts by CRISPR-Cas9 To Create Pathogenicity Mutants in the Multinucleate Filamentous Pathogen Sclerotinia sclerotiorum. mBio 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Liang, Y.; Han, Y.; Wang, C.; Jiang, C.; Xu, J.-R. Targeted Deletion of the USTA and UvSLT2 Genes Efficiently in Ustilaginoidea virens with the CRISPR-Cas9 System. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Ma, J.; Chen, J.; Wang, M.; Ren, Y.; Wang, S.; Lei, C.; Cheng, Z. Sodmergen Disruption of OsSEC3A increases the content of salicylic acid and induces plant defense responses in rice. J. Exp. Bot. 2018, 69, 1051–1064. [Google Scholar] [CrossRef]
- Wang, X.; Tu, M.; Wang, D.; Liu, J.; Li, Y.; Li, Z.; Wang, Y.; Wang, X. CRISPR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation. Plant Biotechnol. J. 2018, 16, 844–855. [Google Scholar] [CrossRef]
- Wang, Q.; Cobine, P.A.; Coleman, J.J. Efficient genome editing in Fusarium oxysporum based on CRISPR/Cas9 ribonucleoprotein complexes. Fungal Genet. Boil. 2018, 117, 21–29. [Google Scholar] [CrossRef]
- Miao, J.; Chi, Y.; Lin, D.; Tyler, B.M.; Liu, X.L. Mutations in ORP1 Conferring Oxathiapiprolin Resistance Confirmed by Genome Editing using CRISPR/Cas9 in Phytophthora capsici and P. sojae. Phytopathology 2018, 108, 1412–1419. [Google Scholar] [CrossRef]
- Wang, Q.; Coleman, J.J. CRISPR/Cas9-mediated endogenous gene tagging in Fusarium oxysporum. Fungal Genet. Boil. 2019, 126, 17–24. [Google Scholar] [CrossRef]
- Kong, G.; Wan, L.; Deng, Y.Z.; Yang, W.; Li, W.; Jiang, L.; Situ, J.; Xi, P.; Li, M.; Jiang, Z. Pectin acetylesterase PAE5 is associated with the virulence of plant pathogenic oomycete Peronophythora litchii. Physiol. Mol. Plant Pathol. 2019, 106, 16–22. [Google Scholar] [CrossRef]
- Schuster, M.; Schweizer, G.; Reissmann, S.; Kahmann, R. Genome editing in Ustilago maydis using the CRISPR-Cas system. Fungal Genet. Boil. 2016, 89, 3–9. [Google Scholar] [CrossRef]
- Fister, A.S.; Landherr, L.; Maximova, S.N.; Guiltinan, M.J. Transient Expression of CRISPR/Cas9 Machinery Targeting TcNPR3 Enhances Defense Response in Theobroma cacao. Front. Plant Sci. 2018, 9, 268. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, Y.; Wu, G.; Zou, S.; Chen, Y.; Gao, C.; Tang, D. Simultaneous modification of three homoeologs of TaEDR1 by genome editing enhances powdery mildew resistance in wheat. Plant J. 2017, 91, 714–724. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef]
- Fang, Y.; Tyler, B.M. Efficient disruption and replacement of an effector gene in the oomycete Phytophthora sojae using CRISPR/Cas9. Mol. Plant Pathol. 2016, 17, 127–139. [Google Scholar] [CrossRef]
- Wang, F.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Zhao, K. Enhanced Rice Blast Resistance by CRISPR/Cas9-Targeted Mutagenesis of the ERF Transcription Factor Gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef]
- Idnurm, A.; Urquhart, A.S.; Vummadi, D.R.; Chang, S.; Van De Wouw, A.P.; López-Ruiz, F.J. Spontaneous and CRISPR/Cas9-induced mutation of the osmosensor histidine kinase of the canola pathogen Leptosphaeria maculans. Fungal Boil. Biotechnol. 2017, 4, 12. [Google Scholar] [CrossRef]
- Wenderoth, M.; Pinecker, C.; Voß, B.; Fischer, R. Establishment of CRISPR/Cas9 in Alternaria alternata. Fungal Genet. Boil. 2017, 101, 55–60. [Google Scholar] [CrossRef]
- Sera, T. Inhibition of Virus DNA Replication by Artificial Zinc Finger Proteins. J. Virol. 2005, 79, 2614–2619. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Qian, Y.; Wu, X.; Sun, Y.; Wu, X.; Cheng, X. Inhibiting replication of begomoviruses using artificial zinc finger nucleases that target viral-conserved nucleotide motif. Virus Genes 2014, 48, 494–501. [Google Scholar] [CrossRef]
- Zhang, T.; Zheng, Q.; Yi, X.; An, H.; Zhao, Y.; Ma, S.; Zhou, G. Establishing RNA virus resistance in plants by harnessing CRISPR immune system. Plant Biotechnol. J. 2018, 16, 1415–1423. [Google Scholar] [CrossRef] [Green Version]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Boil. 2018, 19, 1. [Google Scholar] [CrossRef]
- Kis, A.; Hamar, É.; Tholt, G.; Bán, R.; Havelda, Z. Creating highly efficient resistance against wheat dwarf virus in barley by employing CRISPR /Cas9 system. Plant Biotechnol. J. 2019, 17, 1004–1006. [Google Scholar] [CrossRef]
- Bastet, A.; Zafirov, D.; Giovinazzo, N.; Guyon-Debast, A.; Nogué, F.; Robaglia, C.; Gallois, J.-L. Mimicking natural polymorphism in eIF4E by CRISPR-Cas9 base editing is associated with resistance to potyviruses. Plant Biotechnol. J. 2019, 17, 1736–1750. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musa spp. overcomes a major challenge in banana breeding. Commun. Boil. 2019, 2, 46. [Google Scholar] [CrossRef]
- Ali, Z.; Ali, S.; Tashkandi, M.; Zaidi, S.S.-E.-A.; Mahfouz, M.M. CRISPR/Cas9-Mediated Immunity to Geminiviruses: Differential Interference and Evasion. Sci. Rep. 2016, 6, 26912. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Li, F.; Cai, J.; Chen, W.; Zhao, N.; Sun, Y.; Guo, Y.; Yang, X.; Wu, X. Artificial TALE as a Convenient Protein Platform for Engineering Broad-Spectrum Resistance to Begomoviruses. Viruses 2015, 7, 4772–4782. [Google Scholar] [CrossRef]
- Pyott, D.E.; Sheehan, E.; Molnár, A. Engineering of CRISPR/Cas9-mediated potyvirus resistance in transgene-free Arabidopsis plants. Mol. Plant Pathol. 2016, 17, 1276–1288. [Google Scholar] [CrossRef]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef] [Green Version]
- Macovei, A.; Sevilla, N.R.; Cantos, C.; Jonson, G.B.; Slamet-Loedin, I.; Čermák, T.; Voytas, D.F.; Choi, I.-R.; Chadha-Mohanty, P. Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant Biotechnol. J. 2018, 16, 1918–1927. [Google Scholar] [CrossRef]
- Ali, Z.; Abulfaraj, A.; Idris, A.; Ali, S.; Tashkandi, M.; Mahfouz, M.M. CRISPR/Cas9-mediated viral interference in plants. Genome Boil. 2015, 16, 267. [Google Scholar] [CrossRef]
- Baltes, N.J.; Hummel, A.W.; Konečná, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR–Cas prokaryotic immune system. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPR–Cas-like immune system conferring DNA virus resistance in plants. Nat. Plants 2015, 1, 15144. [Google Scholar] [CrossRef]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J.; et al. Simultaneous CRISPR/Cas9-mediated editing of cassava eIF4E isoforms nCBP-1 and nCBP-2 reduces cassava brown streak disease symptom severity and incidence. Plant Biotechnol. J. 2019, 17, 421–434. [Google Scholar] [CrossRef]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 2015, 1, 15007. [Google Scholar] [CrossRef]
- Jiao, Y.; Li, Y.; Li, Y.; Cao, H.; Mao, Z.; Ling, J.; Yang, Y.; Xie, B. Functional genetic analysis of the leucinostatin biosynthesis transcription regulator lcsL in Purpureocillium lilacinum using CRISPR-Cas9 technology. Appl. Microbiol. Biotechnol. 2019, 103, 6187–6194. [Google Scholar] [CrossRef]
- Chen, J.; Lai, Y.; Wang, L.; Zhai, S.; Zou, G.; Zhou, Z.; Cui, C.; Wang, S. CRISPR/Cas9-mediated efficient genome editing via blastospore-based transformation in entomopathogenic fungus Beauveria bassiana. Sci. Rep. 2017, 7, 45763. [Google Scholar] [CrossRef]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant–fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1, 1–11. [Google Scholar] [CrossRef]
- Hiruma, K.; Gerlach, N.; Sacristán, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramírez, D.; Bucher, M.; O’Connell, R.J.; et al. Root Endophyte Colletotrichum tofieldiae Confers Plant Fitness Benefits that Are Phosphate Status Dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef]
- Geurts, R.; Xiao, T.T.; Reinhold-Hurek, B.; Information, P.E.K.F.C. What Does It Take to Evolve A Nitrogen-Fixing Endosymbiosis? Trends Plant Sci. 2016, 21, 199–208. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Tan, Q.; Fan, Q.; Zhu, H.; Hong, Z.; Zhang, Z.; Duanmu, D. Efficient Inactivation of Symbiotic Nitrogen Fixation Related Genes in Lotus japonicus Using CRISPR-Cas9. Front. Plant Sci. 2016, 7, 76. [Google Scholar] [CrossRef]
- Curtin, S.J. Editing the Medicago truncatula Genome: Targeted Mutagenesis Using the CRISPR-Cas9 Reagent. Adv. Struct. Saf. Stud. 2018, 161–174. [Google Scholar]
- Li, Z.; Liu, Z.-B.; Xing, A.; Moon, B.P.; Koellhoffer, J.P.; Huang, L.; Ward, R.T.; Clifton, E.; Falco, S.C.; Cigan, A.M. Cas9-Guide RNA Directed Genome Editing in Soybean. Plant Physiol. 2015, 169, 960–970. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Zhang, C.; Sun, Z.; Wang, L.; Duanmu, D.; Fan, Q. Genome Editing in Cowpea Vigna unguiculata Using CRISPR-Cas9. Int. J. Mol. Sci. 2019, 20, 2471. [Google Scholar] [CrossRef]
- Yi, Y.; Li, Z.; Song, C.; Kuipers, O.P. Exploring plant-microbe interactions of the rhizobacteria Bacillus subtilis and Bacillus mycoides by use of the CRISPR-Cas9 system. Environ. Microbiol. 2018, 20, 4245–4260. [Google Scholar] [CrossRef]
- Avellan, A.; Yun, J.; Zhang, Y.; Spielman-Sun, E.; Unrine, J.M.; Thieme, J.; Li, J.; Lombi, E.; Bland, G.; Lowry, G.V. Nanoparticle Size and Coating Chemistry Control Foliar Uptake Pathways, Translocation, and Leaf-to-Rhizosphere Transport in Wheat. ACS Nano 2019, 13, 5291–5305. [Google Scholar] [CrossRef]
- Castro, M.J.L.; Ojeda, C.; Cirelli, A.F. Advances in surfactants for agrochemicals. Environ. Chem. Lett. 2014, 12, 85–95. [Google Scholar] [CrossRef]
- Monreal, C.M.; DeRosa, M.; Mallubhotla, S.C.; Bindraban, P.S.; Dimkpa, C. Nanotechnologies for increasing the crop use efficiency of fertilizer-micronutrients. Biol. Fertil. Soils 2016, 52, 423–437. [Google Scholar] [CrossRef]
- Qiu, Z.; Egidi, E.; Liu, H.; Kaur, S.; Singh, B.K. New frontiers in agriculture productivity: Optimised microbial inoculants and in situ microbiome engineering. Biotechnol. Adv. 2019. [Google Scholar] [CrossRef]
- Pyne, M.E.; Narcross, L.; Martin, V.J.J. Engineering Plant Secondary Metabolism in Microbial Systems. Plant Physiol. 2019, 179, 844–861. [Google Scholar] [CrossRef] [Green Version]
- Rybakova, D.; Mancinelli, R.; Wikström, M.; Birch-Jensen, A.-S.; Postma, J.; Ehlers, R.-U.; Goertz, S.; Berg, G. The structure of the Brassica napus seed microbiome is cultivar-dependent and affects the interactions of symbionts and pathogens. Microbiome 2017, 5, 104. [Google Scholar] [CrossRef]
- Lv, H.; Li, J.; Wu, Y.; Garyali, S.; Wang, Y. Transporter and its engineering for secondary metabolites. Appl. Microbiol. Biotechnol. 2016, 100, 6119–6130. [Google Scholar] [CrossRef]
- Podolsky, I.A.; Seppälä, S.; Lankiewicz, T.S.; Brown, J.L.; Swift, C.L.; O’Malley, M.A. Harnessing Nature’s Anaerobes for Biotechnology and Bioprocessing. Annu. Rev. Chem. Biomol. Eng. 2019, 10, 105–128. [Google Scholar] [CrossRef]
- Chen, H.; Wu, H.; Yan, B.; Zhao, H.; Liu, F.; Zhang, H.; Sheng, Q.; Miao, F.; Liang, Z. Core Microbiome of Medicinal Plant Salvia miltiorrhiza Seed: A Rich Reservoir of Beneficial Microbes for Secondary Metabolism? Int. J. Mol. Sci. 2018, 19, 672. [Google Scholar] [CrossRef]
- Liu, Y.; Zuo, S.; Zou, Y.; Wang, J.; Song, W. Investigation on diversity and population succession dynamics of endophytic bacteria from seeds of maize (Zea mays L., Nongda108) at different growth stages. Ann. Microbiol. 2013, 63, 71–79. [Google Scholar] [CrossRef]
- Klaedtke, S.; Jacques, M.-A.; Raggi, L.; Préveaux, A.; Bonneau, S.; Negri, V.; Chable, V.; Barret, M. Terroir is a key driver of seed-associated microbial assemblages. Environ. Microbiol. 2016, 18, 1792–1804. [Google Scholar] [CrossRef]
- Midha, S.; Bansal, K.; Sharma, S.; Kumar, N.; Patil, P.P.; Chaudhry, V.; Patil, P.B. Genomic Resource of Rice Seed Associated Bacteria. Front. Microbiol. 2016, 6, 282. [Google Scholar] [CrossRef]
- Köberl, M.; Schmidt, R.; Ramadan, E.M.; Bauer, R.; Berg, G. The microbiome of medicinal plants: Diversity and importance for plant growth, quality and health. Front. Microbiol. 2013, 4, 1–9. [Google Scholar] [CrossRef]
- Shi, T.-Q.; Liu, G.-N.; Shi, K.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. CRISPR/Cas9-based genome editing of the filamentous fungi: The state of the art. Appl. Microbiol. Biotechnol. 2017, 101, 7435–7443. [Google Scholar] [CrossRef]
- Shanmugam, K.; Ramalingam, S.; Venkataraman, G.; Hariharan, G.N. The CRISPR/Cas9 System for Targeted Genome Engineering in Free-Living Fungi: Advances and Opportunities for Lichenized Fungi. Front. Microbiol. 2019, 10, 62. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Du, G.; Chen, J.; Zhou, J. Enhancing flavonoid production by systematically tuning the central metabolic pathways based on a CRISPR interference system in Escherichia coli. Sci. Rep. 2015, 5, 13477. [Google Scholar] [CrossRef]
- Liu, Q.; Gao, R.; Li, J.; Lin, L.; Zhao, J.; Sun, W.; Tian, C. Development of a genome-editing CRISPR/Cas9 system in thermophilic fungal Myceliophthora species and its application to hyper-cellulase production strain engineering. Biotechnol. Biofuels 2017, 10, 165. [Google Scholar] [CrossRef]
- Kuivanen, J.; Wang, Y.M.J.; Richard, P. Engineering Aspergillus niger for galactaric acid production: Elimination of galactaric acid catabolism by using RNA sequencing and CRISPR/Cas9. Microb. Cell Fact. 2016, 15, 210. [Google Scholar] [CrossRef]
- Katayama, T.; Tanaka, Y.; Okabe, T.; Nakamura, H.; Fujii, W.; Kitamoto, K.; Maruyama, J.-I. Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 2016, 38, 637–642. [Google Scholar] [CrossRef]
- Deng, H.; Gao, R.; Liao, X.; Cai, Y. Genome editing in Shiraia bambusicola using CRISPR-Cas9 system. J. Biotechnol. 2017, 259, 228–234. [Google Scholar] [CrossRef]
- Weber, J.; Valiante, V.; Nødvig, C.S.; Mattern, D.J.; Slotkowski, R.A.; Mortensen, U.H.; Brakhage, A.A. Functional Reconstitution of a Fungal Natural Product Gene Cluster by Advanced Genome Editing. ACS Synth. Biol. 2017, 6, 62–68. [Google Scholar] [CrossRef]
- Alagoz, Y.; Gürkök, T.; Zhang, B.; Unver, T. Manipulating the Biosynthesis of Bioactive Compound Alkaloids for Next-Generation Metabolic Engineering in Opium Poppy Using CRISPR-Cas 9 Genome Editing Technology. Sci. Rep. 2016, 6, 30910. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Li, R.; Li, X.; Fu, D.; Zhu, B.; Tian, H.; Luo, Y.; Zhu, H. Multiplexed CRISPR/Cas9-mediated metabolic engineering of γ-aminobutyric acid levels in Solanum lycopersicum. Plant Biotechnol. J. 2018, 16, 415–427. [Google Scholar] [CrossRef]
- Li, B.; Cui, G.; Shen, G.; Zhan, Z.; Huang, L.; Chen, J.; Qi, X. Targeted mutagenesis in the medicinal plant Salvia miltiorrhiza. Sci. Rep. 2017, 7, 43320. [Google Scholar] [CrossRef]
- Kui, L.; Chen, H.; Zhang, W.; He, S.; Xiong, Z.; Zhang, Y.; Yan, L.; Zhong, C.; He, F.; Chen, J.; et al. Building a Genetic Manipulation Tool Box for Orchid Biology: Identification of Constitutive Promoters and Application of CRISPR/Cas9 in the Orchid, Dendrobium officinale. Front. Plant Sci. 2016, 7, 2036. [Google Scholar] [CrossRef]
- Aznar-Moreno, J.A.; Durrett, T.P. Simultaneous Targeting of Multiple Gene Homeologs to Alter Seed Oil Production in Camelina sativa. Plant Cell Physiol. 2017, 58, 1260–1267. [Google Scholar] [CrossRef]
- Mercx, S.; Smargiasso, N.; Chaumont, F.; De Pauw, E.; Boutry, M.; Navarre, C. Inactivation of the β(1,2)-xylosyltransferase and the α(1,3)-fucosyltransferase genes in Nicotiana tabacum BY-2 Cells by a Multiplex CRISPR/Cas9 Strategy Results in Glycoproteins without Plant-Specific Glycans. Front. Plant Sci. 2017, 8, 99. [Google Scholar] [CrossRef]
- Zhou, Z.; Tan, H.; Li, Q.; Chen, J.; Gao, S.; Wang, Y.; Chen, W.; Zhang, L. CRISPR/Cas9-mediated efficient targeted mutagenesis of RAS in Salvia miltiorrhiza. Phytochemistry 2018, 148, 63–70. [Google Scholar] [CrossRef]
- Barrangou, R.; Doudna, J.A. Applications of CRISPR technologies in research and beyond. Nat. Biotechnol. 2016, 34, 933–941. [Google Scholar] [CrossRef]
- Armanhi, J.S.L.; De Souza, R.S.C.; Damasceno, N.D.B.; De Araújo, L.M.; Imperial, J.; Arruda, P. A Community-Based Culture Collection for Targeting Novel Plant Growth-Promoting Bacteria from the Sugarcane Microbiome. Front. Plant Sci. 2018, 8, 1–17. [Google Scholar] [CrossRef]
- Sarhan, M.S.; Hamza, M.A.; Youssef, H.H.; Patz, S.; Becker, M.; Elsawey, H.; Nemr, R.; Daanaa, H.-S.A.; Mourad, E.F.; Morsi, A.T.; et al. Culturomics of the plant prokaryotic microbiome and the dawn of plant-based culture media—A review. J. Adv. Res. 2019, 19, 15–27. [Google Scholar] [CrossRef]
- Wan, T.; Niu, D.; Wu, C.; Xu, F.-J.; Church, G.; Ping, Y. Material solutions for delivery of CRISPR/Cas-based genome editing tools: Current status and future outlook. Mater. Today 2019, 26, 40–66. [Google Scholar] [CrossRef]
- Zetche, B.; Volz, S.E.; Zhang, F. A Split Cas9 Architecture for Inducible Genome Editing and Transcription Modulation. Nat. Biotechnol. 2015, 33, 139–142. [Google Scholar] [CrossRef]
- Kaya, H.; Ishibashi, K.; Toki, S. A Split Staphylococcus aureus Cas9 as a Compact Genome-Editing Tool in Plants. Plant Cell Physiol. 2017, 58, 643–649. [Google Scholar] [CrossRef]
- Klompe, S.E.; Vo, P.L.H.; Halpin-Healy, T.S.; Sternberg, S.H. Transposon-encoded CRISPR–Cas systems direct RNA-guided DNA integration. Nature 2019, 571, 219–225. [Google Scholar] [CrossRef]
- Strecker, J.; Ladha, A.; Gardner, Z.; Schmid-Burgk, J.L.; Makarova, K.S.; Koonin, E.V.; Zhang, F. RNA-guided DNA insertion with CRISPR-associated transposases. Science 2019, 365, 48–53. [Google Scholar] [CrossRef]
- Baskaran, P.; Soós, V.; Balázs, E.; Van Staden, J. Shoot apical meristem injection: A novel and efficient method to obtain transformed cucumber plants. S. Afr. J. Bot. 2016, 103, 210–215. [Google Scholar] [CrossRef]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Mao, Y.; Lu, Y.; Wang, Z.; Tao, X.; Zhu, J.-K. Multiplex gene editing in rice with simplified CRISPR-Cpf1 and CRISPR-Cas9 systems. J. Integr. Plant Boil. 2018, 60, 626–631. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalan, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef]
- Van Vu, T.; Sivankalyani, V.; Kim, E.-J.; Tran, M.T.; Kim, J.; Sung, Y.W.; Doan, D.T.H.; Kim, J.-Y. Homology-directed repair using next-generation CRISPR/Cpf1-geminiviral replicons in tomato. bioRxiv 2019, 521419. [Google Scholar] [CrossRef]
- Bin Moon, S.; Kim, D.Y.; Ko, J.-H.; Kim, J.-S.; Kim, Y.-S. Improving CRISPR Genome Editing by Engineering Guide RNAs. Trends Biotechnol. 2019, 37, 870–881. [Google Scholar] [CrossRef]
- Eckerstorfer, M.F.; Engelhard, M.; Heissenberger, A.; Simon, S.; Teichmann, H. Plants Developed by New Genetic Modification Techniques—Comparison of Existing Regulatory Frameworks in the EU and Non-EU Countries. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef]
Figure 1.
Microbiome in plant ecosystem. Schematic plant and plant-associated microbiota colonizing different niches on and inside the plant tissue. All the above-ground plant parts together, called the phyllosphere, are a continuously evolving habitat due to ultraviolet (UV) radiation and altering climatic conditions. It is primarily composed of leaves. Below-ground plant parts, mainly roots, are generally influenced by soil properties. Harmful interactions affect the plant growth through pathogenic activities of some microbiota members (left side). On the other hand, beneficial microbial interactions promote plant growth (right side).
Figure 1.
Microbiome in plant ecosystem. Schematic plant and plant-associated microbiota colonizing different niches on and inside the plant tissue. All the above-ground plant parts together, called the phyllosphere, are a continuously evolving habitat due to ultraviolet (UV) radiation and altering climatic conditions. It is primarily composed of leaves. Below-ground plant parts, mainly roots, are generally influenced by soil properties. Harmful interactions affect the plant growth through pathogenic activities of some microbiota members (left side). On the other hand, beneficial microbial interactions promote plant growth (right side).

Figure 2.
Driving factors of plant–microbe interactions. Environment-, soil- and plant-mediated factors determine the composition and structure of host microbiota. Furthermore, plant–plant, microbe–microbe, and plant–microbe interactions also impact the plant and soil microbiome.
Figure 2.
Driving factors of plant–microbe interactions. Environment-, soil- and plant-mediated factors determine the composition and structure of host microbiota. Furthermore, plant–plant, microbe–microbe, and plant–microbe interactions also impact the plant and soil microbiome.

Figure 3.
Biology and components of site-specific nucleases (SSNs) modified for genome editing applications. (A) Repair of double-strand breaks (DSBs) in damaged DNA strands occurs through two main pathways that consist of non-homologous end-joining (NHEJ) and homology-directed repair (HDR). NHEJ is most common in cells. It is an error-prone pathway that introduces indel mutations (small insertions or deletions). HDR is more precise compared to NHEJ, but it requires a donor template that results in either insertion or replacement. (B) Zinc finger nuclease (ZNF) is designed using an array of DNA-binding domains from zinc-finger proteins. Each ZFN comprises DNA-binding domain at N-terminus and FokI nuclease at C-terminus. The linker and spacers are shown in black and pink, respectively. (C) Design of transcription activator-like effectors (TALE) protein-based nuclease (TALEN) bound to DNA. (D) Illustration of clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9 with a sgRNA (red) encoding gRNA (green) bound to a target DNA (blue) adjacent to PAM, i.e., protospacer adjacent motif (magenta). Cleavage sites on both strands shown with scissors and dotted line depict the blunt ends produced by Cas9. (E) CRISPR/Cpf1 (Cas12a) system shown with a crRNA (red), a gRNA (green), target DNA (blue), PAM (magenta). Cleavage sites of both strands (scissors) produce staggered ends (dotted line) with Cpf1.
Figure 3.
Biology and components of site-specific nucleases (SSNs) modified for genome editing applications. (A) Repair of double-strand breaks (DSBs) in damaged DNA strands occurs through two main pathways that consist of non-homologous end-joining (NHEJ) and homology-directed repair (HDR). NHEJ is most common in cells. It is an error-prone pathway that introduces indel mutations (small insertions or deletions). HDR is more precise compared to NHEJ, but it requires a donor template that results in either insertion or replacement. (B) Zinc finger nuclease (ZNF) is designed using an array of DNA-binding domains from zinc-finger proteins. Each ZFN comprises DNA-binding domain at N-terminus and FokI nuclease at C-terminus. The linker and spacers are shown in black and pink, respectively. (C) Design of transcription activator-like effectors (TALE) protein-based nuclease (TALEN) bound to DNA. (D) Illustration of clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9 with a sgRNA (red) encoding gRNA (green) bound to a target DNA (blue) adjacent to PAM, i.e., protospacer adjacent motif (magenta). Cleavage sites on both strands shown with scissors and dotted line depict the blunt ends produced by Cas9. (E) CRISPR/Cpf1 (Cas12a) system shown with a crRNA (red), a gRNA (green), target DNA (blue), PAM (magenta). Cleavage sites of both strands (scissors) produce staggered ends (dotted line) with Cpf1.

Figure 4.
Schematic representation of Cas9 and Cpf1 variants including domain organization. Color scheme and nomenclature to represent domains followed from published structures for each enzyme. MbCpf1 structure is not yet available, hence protein domains are not drawn. Abbreviations: RuvCI-III, RuvC nuclease domain; Arg, arginine-rich bridge helix; REC1-3, recognition lobe; HNH, HNH-like nuclease domain; CTD, C-terminal domain; WEDI-III, wedge domain; PI, PAM-interacting domain; BH, bridge helix; Nuc, novel nuclease domain; OBD, oligonucleotide-binding domain; LHD, looped-out helical domain; HLH, helix-loop-helix; UK, domain with unknown functions.
Figure 4.
Schematic representation of Cas9 and Cpf1 variants including domain organization. Color scheme and nomenclature to represent domains followed from published structures for each enzyme. MbCpf1 structure is not yet available, hence protein domains are not drawn. Abbreviations: RuvCI-III, RuvC nuclease domain; Arg, arginine-rich bridge helix; REC1-3, recognition lobe; HNH, HNH-like nuclease domain; CTD, C-terminal domain; WEDI-III, wedge domain; PI, PAM-interacting domain; BH, bridge helix; Nuc, novel nuclease domain; OBD, oligonucleotide-binding domain; LHD, looped-out helical domain; HLH, helix-loop-helix; UK, domain with unknown functions.

Figure 5.
Expanding CRISPR toolkit, including applications beyond genome editing, are shown. The CRISPR-based tools have been designed for various applications in diverse fields. The SpCas9-mediated knock-out or knock-in strategy is most widely implemented in prokaryotes and eukaryotes. Catalytically dead Cas9 (dCas9) or nickase Cas9 (nCas9) and Cas9 orthologs fused with specific modulators have been reprogrammed to perform base editing (ABE, adenine base editor; CBE, cytidine base editor) RNA editing, screening libraries, chromatin imaging, transcription regulation, allele generation using the EvolvR system, and epigenome editing. Further details of each technique discussed in the main text.
Figure 5.
Expanding CRISPR toolkit, including applications beyond genome editing, are shown. The CRISPR-based tools have been designed for various applications in diverse fields. The SpCas9-mediated knock-out or knock-in strategy is most widely implemented in prokaryotes and eukaryotes. Catalytically dead Cas9 (dCas9) or nickase Cas9 (nCas9) and Cas9 orthologs fused with specific modulators have been reprogrammed to perform base editing (ABE, adenine base editor; CBE, cytidine base editor) RNA editing, screening libraries, chromatin imaging, transcription regulation, allele generation using the EvolvR system, and epigenome editing. Further details of each technique discussed in the main text.


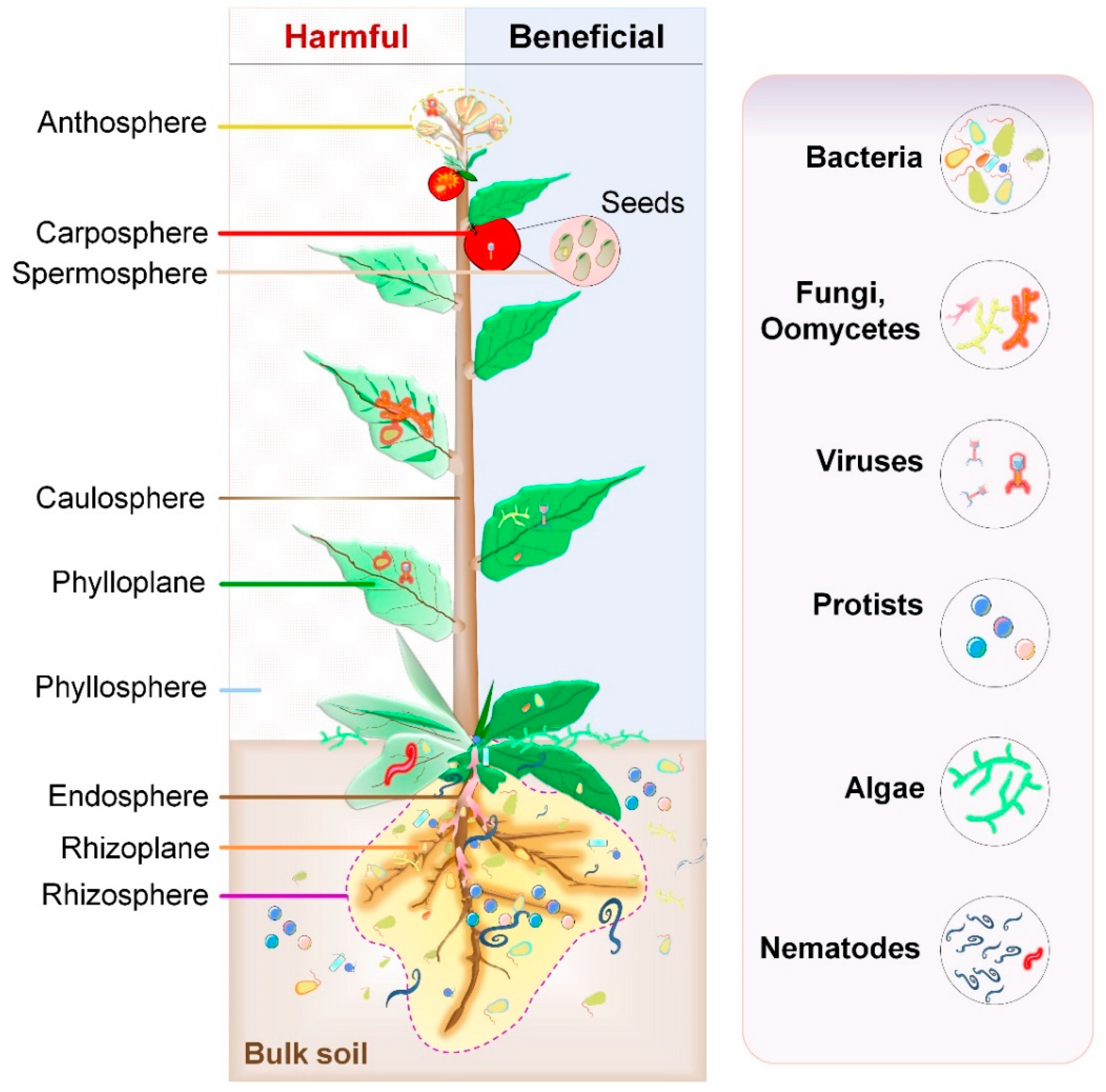{kind=link}
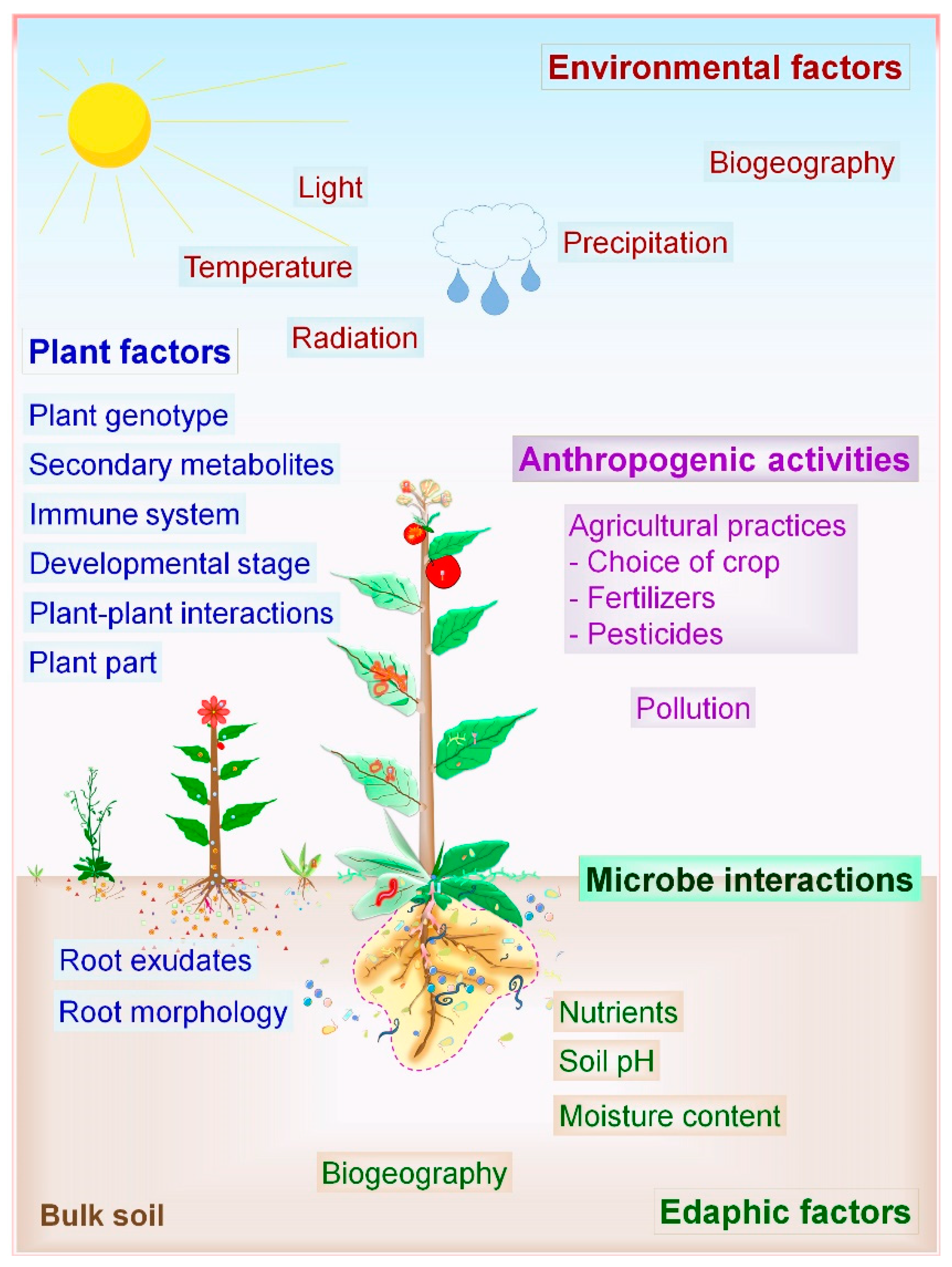{kind=link}
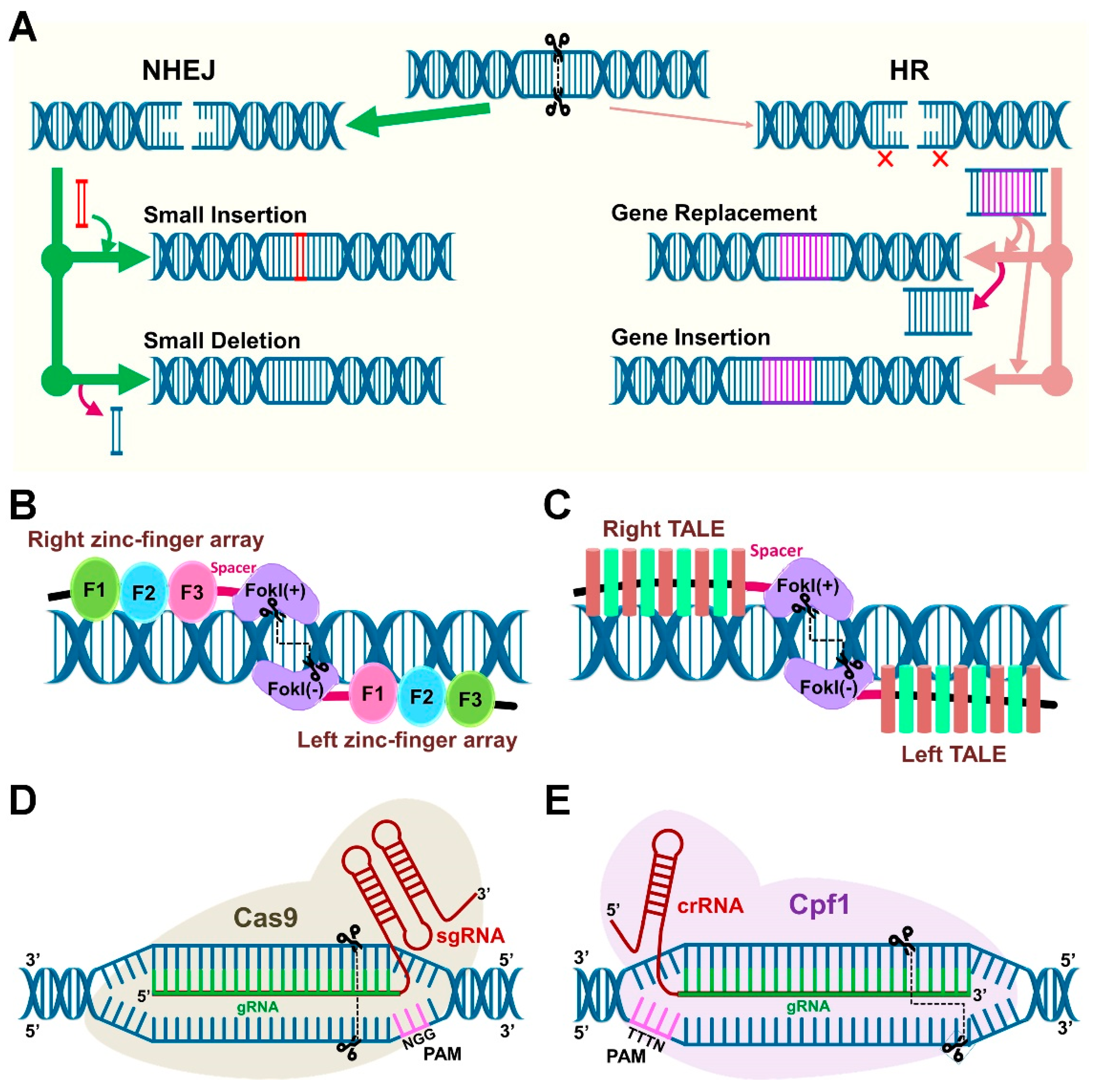{kind=link}
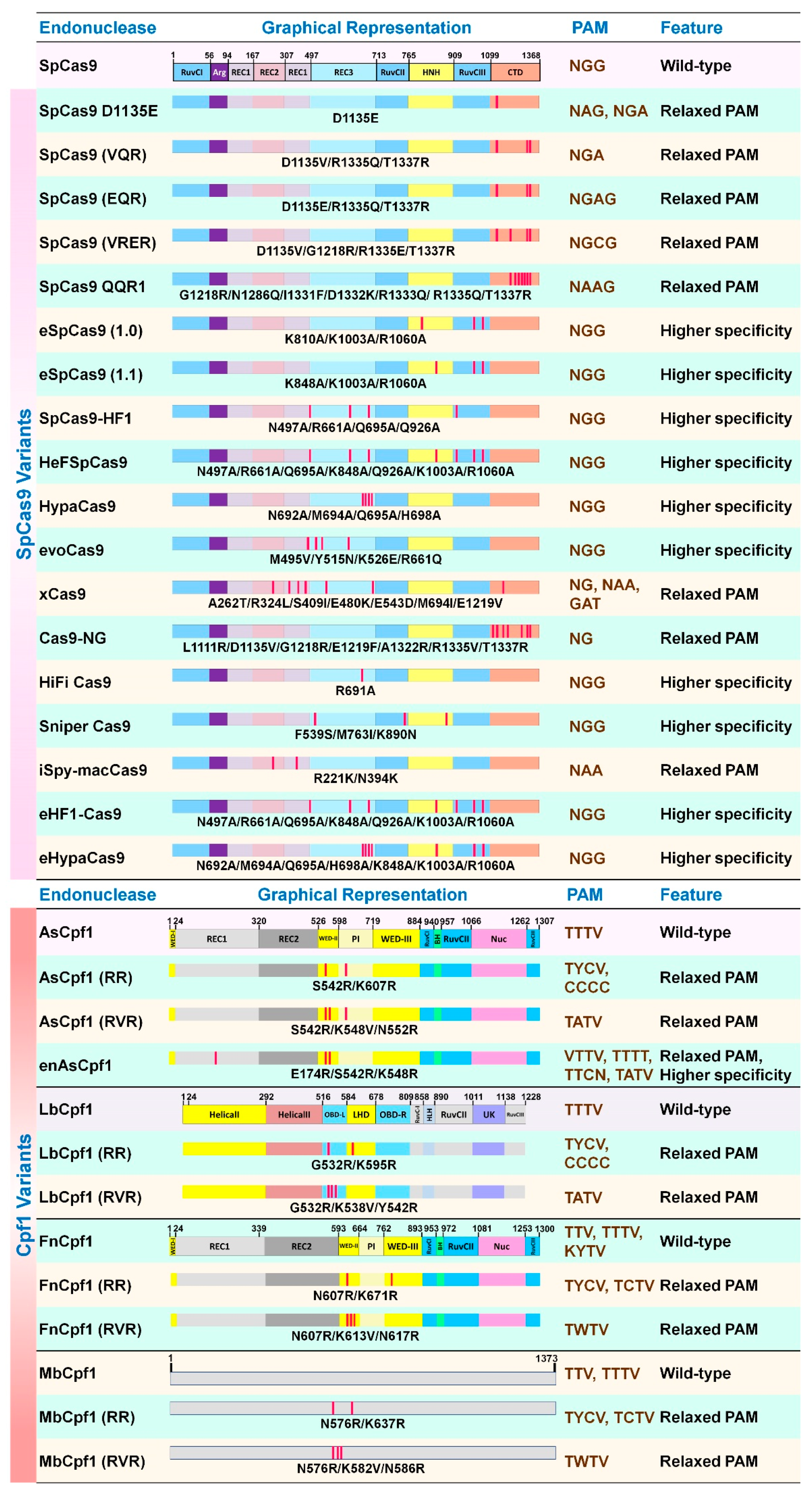{kind=link}
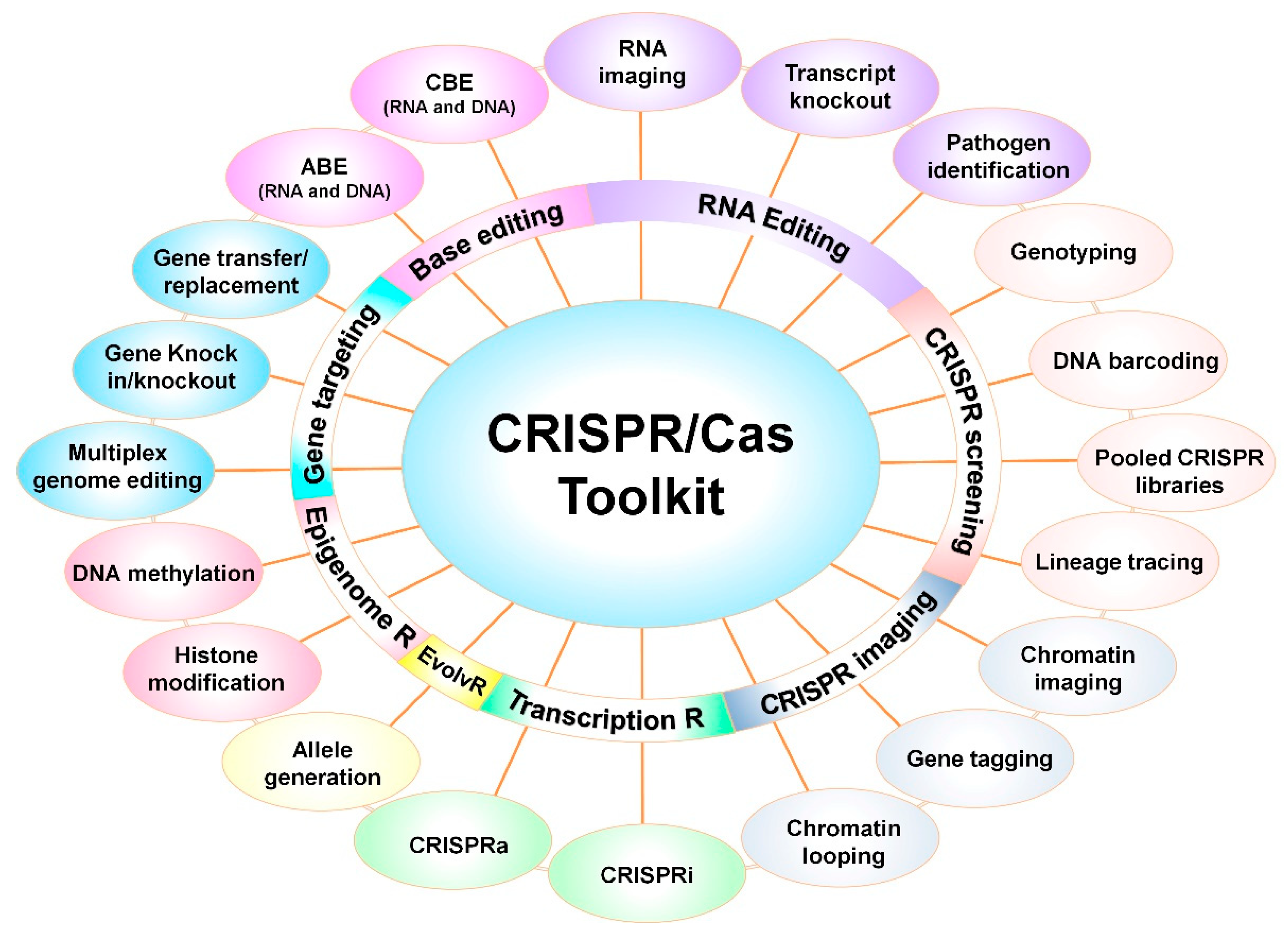{kind=link}
Table 1.
Metagenomic-based studies of plant microbiota are summarized.
| Host | Sampling | Key Findings | Ref. |
|---|---|---|---|
| Agave | Rhizosphere, whole plant | Microbial composition was mainly regulated by the plant compartment, while the fungal community composition was primarily determined by the plant host biogeography. | [9] |
| Arabidopsis | Root, rhizosphere | The composition of rhizospheric microbiota was found reliant on the environment rather than host species. | [10] |
| Arabidopsis | Leaf, root | Genome drafts of 400 isolates revealed a substantial overlap of genome-encoded functional capabilities between leaf- and root-derived bacteria with few significant differences. | [11] |
| Arabidopsis | Root, rhizosphere | Explored genetic network controlling the phosphate stress response influences the structure of the root microbiome community, even under non-stress phosphate conditions. | [12] |
| Arabidopsis | Roots, rhizosphere | Bacterial microbiota is indispensable for plant survival and protection against root-filamentous fungi. | [13] |
| Barley | Root, rhizosphere | Rhizospheric and root microbiota affect plant growth. The interactions between microbe–microbe and plant–microbe drive distinct microbiota. | [14] |
| Citrus | Root, rhizosphere | The core rhizosphere microbiome comprises several potential beneficial plant microbial species and detected over-represented microbial functional traits. | [15] |
| Grapevine | Grape must | Environmental factors, variety, and regional origins determine the unique grapevine-associated microbiota. These factors are the key to the unique taste and wine quality. | [16] |
| Grapevine | Rhizosphere, whole plant | Microbial composition of soil and root is primarily influenced by plant-selective pressure, soil C:N ratio, and pH. Leaf and fruit microbiota alterations correlated with soil carbon, cultivation practices, and geography. | [17] |
| Maize | Roots, rhizosphere | Associated microbiota showed heritable variation in community composition of rhizosphere and significant field-specific heritable variation. | [18] |
| Maize | Roots, rhizosphere | Assembled a simplified and representative synthetic bacterial model community containing seven dominant strains to study the community assembly dynamics that interfered with the growth of a plant-pathogenic fungus. | [19] |
| Maize | Root, rhizosphere | Microbiome composition varies with plant genotype, plant age, and climate events. | [20] |
| Petunia, Arabidopsis | Root, rhizosphere | Root microbiota composition and responses vary substantially in response to the varying phosphorus (P) application. | [21] |
| Potato | Roots, rhizosphere | Early stages of the plant showed the cultivar-dependent composition of bacterial communities, but in the flowering and senescence stages, this was not the case. Furthermore, the population of some species flourished under different ecological conditions more than the other species. | [22] |
| Rice | Root, rhizosphere | Endosphere, rhizoplane, and rhizosphere consist of a diverse microbiome. Cultivation practices influence the diversity of microbiome compositions at each compartment. | [23] |
| Rice | Root, rhizosphere | Type of soil environment (i.e., rhizosphere versus bulk soil) is a driving factor of the structure of the microbial community than the plant age. | [24] |
| Soybean, Wheat | Rhizosphere, root | Soil properties such as pH and nitrate content may influence the composition of root microbiome in agricultural fields. | [25] |
| Sugar beet | Soil after harvesting | Identified crucial bacterial taxa and genes suppressing a fungal root pathogen and showed that plant protection depends on the rhizospheric microbial community. | [26] |
| Sugarcane | Rhizosphere, whole plant | Microbial communities enter primarily from native rhizospheric soil and colonize plant organs in distinct patterns. | [27] |
| Tomato | Rhizosphere, whole plant | Distinct microbial communities found associated with different plant organs. | [28] |
| Tomato | Rhizosphere, whole plant | The study explored the protection role of rhizosphere microbiota against soil-borne pathogen causing wilt disease. | [29] |
| Wheat, Cucumber | Roots from pots | Genus or species level differences observed between the rhizospheric microbiome from diverse plant species related to environmental factors. | [30] |
| Wild mustard | Leaf and root | Leaf microbiome genetically controlled by the host and several bacterial species of leaf microbiomes shared with root microbiomes, suggesting acquisition from the soil. | [31] |
Table 2.
List of genome-edited plant–pathogen interactions is summarized.
| Pathogen | Disease | Host | Target Gene (plant or pathogen), Interaction | GE Tool | Ref. | |
|---|---|---|---|---|---|---|
| Bacteria | ||||||
| 1. | Xanthomonas oryzae pv. oryzae | Bacterial blight | Rice | OsSWEET14 (plant); Pathogen interacts with the promoter of gene and hijacks plant sugars | TALEN | [151] |
| 2. | Xanthomonas oryzae pv. oryzae | Bacterial blight | Rice | OsSWEET14 and OsSWEET11 (plant); Pathogen interacts with the promoter of gene and hijacks plant sugars | CRISPR/Cas9 | [152] |
| 3. | Xanthomonas oryzae pv. oryzae | Bacterial blight | Rice | OsSWEET13 (plant); Pathogen hijacks sucrose from plant cells | TALEN | [153] |
| 4. | Pseudomonas syringae pv. tomato, Xanthomonas spp., Phytophthora capsici | Bacterial speck, Blight, and spot | Tomato | SlDMR6-1 (plant); Knock-out of DMR6 increases salicylic acid levels that induces production of secondary metabolites and PR genes | CRISPR/Cas9 | [154] |
| 5. | Xanthomonas citri subsp. citri | Citrus canker | Citrus | CsLOB1 (plant); Susceptibility gene induced by pathogen | CRISPR/Cas9 | [155] |
| 6. | Xanthomonas citri subsp. citri | Citrus canker | Citrus | CsLOB1 (plant); Susceptibility gene induced by pathogen | CRISPR/Cas9 | [156] |
| 7. | Erwinia amylovora | Fire blight | Apple | DIPM-1, 2 and 4 (plant); Directly interact with the disease-specific gene of bacterial pathogen | CRISPR/Cas9 | [157] |
| 8. | Pseudomonas syringae pv. tomato (Pto) DC3000 | Bacterial speck | Tomato | SlJAZ2 (plant); Directly interact with coronatine produced by bacteria that helps in leaf colonization | CRISPR/Cas9 | [158] |
| Fungi and Oomycetes | ||||||
| 9. | Magnaporthe grisea, Burkholderia glumae | Fungal blast, bacterial blight | Rice | OsMPK5 (plant); A negative regulator of rice defense response | CRISPR/Cas9 | [159] |
| 10. | Blumeria graminis f. sp. tritici | Powdery mildew | Wheat | MLO-A1, B1, and D1 (plant); Confer susceptibility to fungi | CRISPR/Cas9 | [160] |
| 11. | Uncinula necator | Powdery mildew | Grape | MLO-7 (plant); Confer susceptibility to a fungal pathogen | CRISPR/Cas9 | [157] |
| 12. | Ustilago maydis | Corn smut | Maize | bW2 and bE1 (microbe); To evaluate the CRISPR system. | CRISPR/Cas9 | [170] |
| 13. | Phytophthora tropicalis | Black pod disease | Cacao | Non-Expressor of Pathogenesis-Related3 (TcNPR3) gene (plant) | CRISPR/Cas9 | [171] |
| 14. | Blumeria graminis f. sp. tritici | Powdery mildew | Wheat | Three homologs of TaEDR1 (plant); Plays a negative role in plant immunity | CRISPR/Cas9 | [172] |
| 15. | Oidium neolycopersici | Powdery mildew | Tomato | SlMlo1 (plant); Confer susceptibility to fungi | CRISPR/Cas9 | [173] |
| 16. | Phytophthora sojae | Damping off | Soybean | Avr4/6 (microbe); Virulence proteins enter host cells and promote host susceptibility. | CRISPR/Cas9 | [174] |
| 17. | Magnaporthe oryzae | Rice blast | Rice | OsERF922 (plant); Negative regulator of blast fungus | CRISPR/Cas9 | [175] |
| 18. | Leptosphaeria maculans | Blackleg disease | Canola | Histidine kinase (microbe); To study resistance mechanism against a pesticide (Iprodione) | CRISPR/Cas9 | [176] |
| 19. | Alternaria alternata | Black molds | Sunflower | Phosphate decarboxylase pyrG, polyketide-synthase, pksA, and 1,3,8-THN reductase, brm2 (microbe); To establish a CRISPR system | CRISPR/Cas9 | [177] |
| 20. | Magnaporthe oryzae | Rice blast | Rice | Melanin biosynthetic polyketide synthase genes ALB1 and RSY1, succinate dehydrogenase enzyme SDI1 (microbe); Mutations to study the pathogenicity | CRISPR/Cas9 (RNP) | [161] |
| 21. | Sclerotinia sclerotiorum | White mold | Flowers, Vegetables | Oxalate biosynthesis gene Ssoah1 (microbe); Mutations to study the pathogenicity | CRISPR/Cas9 | [162] |
| 22. | Ustilaginoidea virens | False smut | Rice | USTA ustiloxin and UvSLT2 MAP kinase (microbe); To study the gene function | CRISPR/Cas9 | [163] |
| 23. | Magnaporthe oryzae | Rice blast | Rice | OsSEC3A (plant); participate in the exocyst complex and interact with defense proteins | CRISPR/Cas9 | [164] |
| 24. | Botrytis cinerea | Gray mold | Grape | WRKY52 (plant); Transcription factor involved in response to biotic stress | CRISPR/Cas9 | [165] |
| 25. | Fusarium oxysporum | Wilt | Tomato, legumes, cotton | Polyketide synthase PKS4 (microbe); To study gene function | CRISPR/Cas9 (RNP) | [166] |
| 26. | Phytophthora capsici and P. sojae | Powdery mildew, Damping-off | Vegetables, soybean | Oxysterol binding protein-related protein 1 (microbe); To study resistance mechanism against a pesticide (Oxathiapiprolin) | CRISPR/Cas9 | [167] |
| 27. | Fusarium oxysporum | Wilt | Tomato, legumes, cotton | FoSso1 and FoSso2 (microbe); For endogenous tagging of target genes | CRISPR/Cas9 | [168] |
| 28. | Peronophythora litchii | Downy blight | Lychee | Pectin acetylesterase, PAE4, and PAE5 (microbe); to study the pathogenicity | CRISPR/Cas9 | [169] |
| Viruses | ||||||
| 29. | BSCTV | Viral (DNA) | Arabidopsis | Replication origin (microbe) | ZNF | [178] |
| 30. | TYLCCNV, TbCSV | Viral (DNA) | Tobacco | AC1 replication-associated (Rep) protein (microbe) | ZNF | [179] |
| 31. | TYCCNV, TbCSV, TLCYnV | Viral (DNA) | Tobacco | AC1 replication-associated (Rep) protein (microbe) | TALE | [186] |
| 32. | TuMV | Viral (RNA) | Arabidopsis | eIF4E/exon (plant); Directly interact with viral protein and helps viral replication | CRISPR/Cas9 | [187] |
| 33. | CVYV, ZYMV, PRSV-W | Viral (RNA) | Cucumber | eIF4E/exon (plant); Directly interact with viral protein and helps viral replication | CRISPR/Cas9 | [188] |
| 34. | RTSV | Tungro (RNA) | Rice | eIF4G (plant); Directly interact with viral protein and helps viral RNA replication | CRISPR/Cas9 | [189] |
| 35. | TYLCV, BCTV, MeMV | Viral (DNA) | Tobacco | Intergenic region of origin of replication (IR), capsid protein (CP), RCRII motif of Rep protein (microbe) | CRISPR/Cas9 | [190] |
| 36. | BeYDV | Viral (DNA) | Tobacco | Long intergenic region (LIR), Rep protein encoding gene (microbe) | CRISPR/Cas9 | [191] |
| 37. | BSCTV | Viral (DNA) | Arabidopsis, Tobacco | IR, CP and Rep (microbe) | CRISPR/Cas9 | [192] |
| 38. | CBSV | Brown streak (RNA) | Cassava | nCBP-1 & nCBP-2/exon (plant); Directly interact with viral protein and helps viral replication | CRISPR/Cas9 | [193] |
| 39. | TMV | Viral (RNA) | Arabidopsis, Tobacco | ORF1a, ORFCP, 3’- UTR (microbe) | CRISPR/Cas9 | [180] |
| 40. | TuMV | Viral (RNA) | Tobacco | TuMV-GFP, Helper component proteinase silencing suppressor (HC-Pro), coat protein genes (microbe) | CRISPR/Cas13a | [181] |
| 41. | WDV | Viral (DNA) | Barley | Rep, MP, LIR (microbe) | CRISPR/Cas9 | [182] |
| 42. | CYVV | Viral (DNA) | Arabidopsis | eIF4E1 gene (plant); Directly interact with viral protein and helps viral replication | Cas9- PmCDA1 | [183] |
| 43. | eBSV | Viral (DNA) | Banana | Three target sites in viral genome (microbe) | CRISPR/Cas9 | [184] |
| 44. | CLCuKoV, TYLCV, TYLCSV, MeMV, BCTV | Viral (DNA) | Tobacco | IR, coat protein and Rep (microbe) | CRISPR/Cas9 | [185] |
BCTV, Beet curly top virus; BeYDV, Bean yellow dwarf virus; BSCTV, Beet severe curly top virus; CBSV, Cassava brown streak virus; CLCuKoV, Cotton Leaf Curl Kokhran Virus; CRISPR/Cas9, clustered regularly interspaced palindromic repeat-CRISPR-associated protein 9; CVYV, Cucumber vein yellowing virus; CYVV, Clover yellow vein virus; eBSV, Endogenous Banana streak virus; eIF4E, eukaryotic translation initiation factor 4E; MeMV, Merremia mosaic virus; ORF, open reading frame, PmCDA1- Petromyzon marinus cytidine deaminase 1 base editor; PR genes, Pathogenesis-related genes; PRSV-W, Papaya ring spot mosaic virus-W; RTSV, Rice tungro spherical virus; SWEET, sugar will eventually be exported transporter; TALEN, transcription-activator-like effector nuclease; TbCSV, Tobacco curly shoot virus; TLCYnV, Tomato leaf curl Yunnan virus; TMV, Tobacco mosaic virus; TuMV, Turnip mosaic virus; TYLCCNV, Tomato yellow leaf curl China virus; TYLCSV, Tomato yellow leaf curl Sardinia virus; TYLCV, Tomato yellow leaf curl virus; UTR, untranslated terminal repeat; WDV, Wheat dwarf virus; ZNF, Zinc finger protein; ZYMV, Zucchini yellow mosaic virus.
Table 3.
Potential applications of genome-editing in plant–microbe interaction are summarized.
| Trait | Present and Future Applications | Potential CRISPR Tools |
|---|---|---|
| Understanding the fundamentals of the PM interactions | Identification of genes involved in PM interactions | Genotyping, DNA barcoding, lineage tracing |
| Study of gene function in microbe and plant | Cas9, Cpf1 (gene knock-in/knock-out, gene replacement) | |
| Regulation of gene expression, promoter engineering | CRISPRa, CRISPRi (transcription regulation); DNA methylation, histone modification (epigenome editing) | |
| Novel allele generation | EvolvR (diversification of target genomic locus) | |
| Plant disease resistance | Functional characterization of pathogenesis-related factors | Cas9, Cpf1 (gene knock-in/knock-out) |
| Phytopathogen identification | Cas13 (RNA editing tool), RNA base editors | |
| Development of disease-resistant plant varieties | Cas9, Cpf1 (gene knock-in/knock-out, gene replacement), ABE/CBE (base editing) | |
| Pyramiding of multiple disease-resistant traits | Multiplex GE | |
| Pesticide resistance in crops | Cas9, Cpf1(gene knock-in/knock-out, gene replacement), ABE/CBE (base editing) | |
| Plant growth promotion and nutrient uptake | Improvement of nutrient accessibility (biological nitrogen fixation, phosphate solubilization) | Cas9, Cpf1 (gene knock-in/knock-out, gene replacement) |
| Application of nodulation in non-leguminous crops through pathway engineering | Cas9, Cpf1 (gene replacement, multiplex GE) | |
| Improved stress resistance by signaling molecules | Cas9, Cpf1 (gene knock-in/knock-out, gene transfer/replacement) | |
| Engineered microbes to reduce cost and chemical use | Cas9, Cpf1 (gene knock-in/knock-out, gene replacement, multiplex GE) | |
| Metabolic engineering | Exploration of the novel plant metabolome pathways | Cas9, Cpf1 (gene knock-in/knock-out, gene replacement, multiplex GE) |
| Secondary metabolites | Cas9, Cpf1 (gene knock-in/knock-out, gene replacement, multiplex GE) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shelake, R.M.; Pramanik, D.; Kim, J.-Y. Exploration of Plant-Microbe Interactions for Sustainable Agriculture in CRISPR Era. Microorganisms 2019, 7, 269. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080269
AMA Style
Shelake RM, Pramanik D, Kim J-Y. Exploration of Plant-Microbe Interactions for Sustainable Agriculture in CRISPR Era. Microorganisms. 2019; 7(8):269. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080269
Chicago/Turabian StyleShelake, Rahul Mahadev, Dibyajyoti Pramanik, and Jae-Yean Kim. 2019. "Exploration of Plant-Microbe Interactions for Sustainable Agriculture in CRISPR Era" Microorganisms 7, no. 8: 269. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080269
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.