Effects on Gene Transcription Profile and Fatty Acid Composition by Genetic Modification of Mevalonate Diphosphate Decarboxylase MVD/Erg19 in Aspergillus Oryzae

 ,
,
Abstract
:1. Introduction
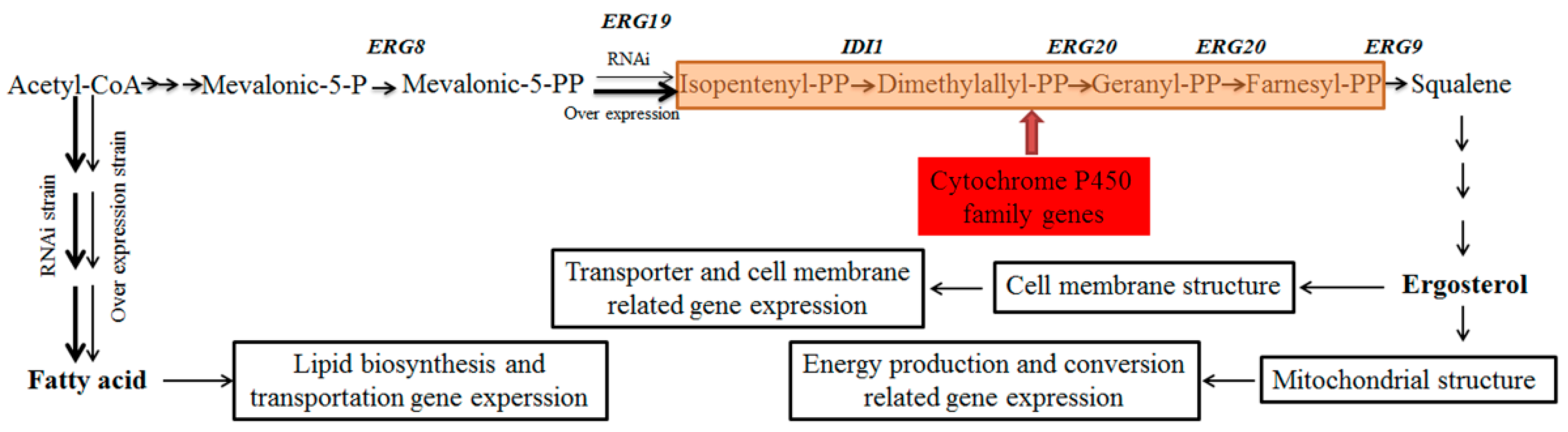
2. Results
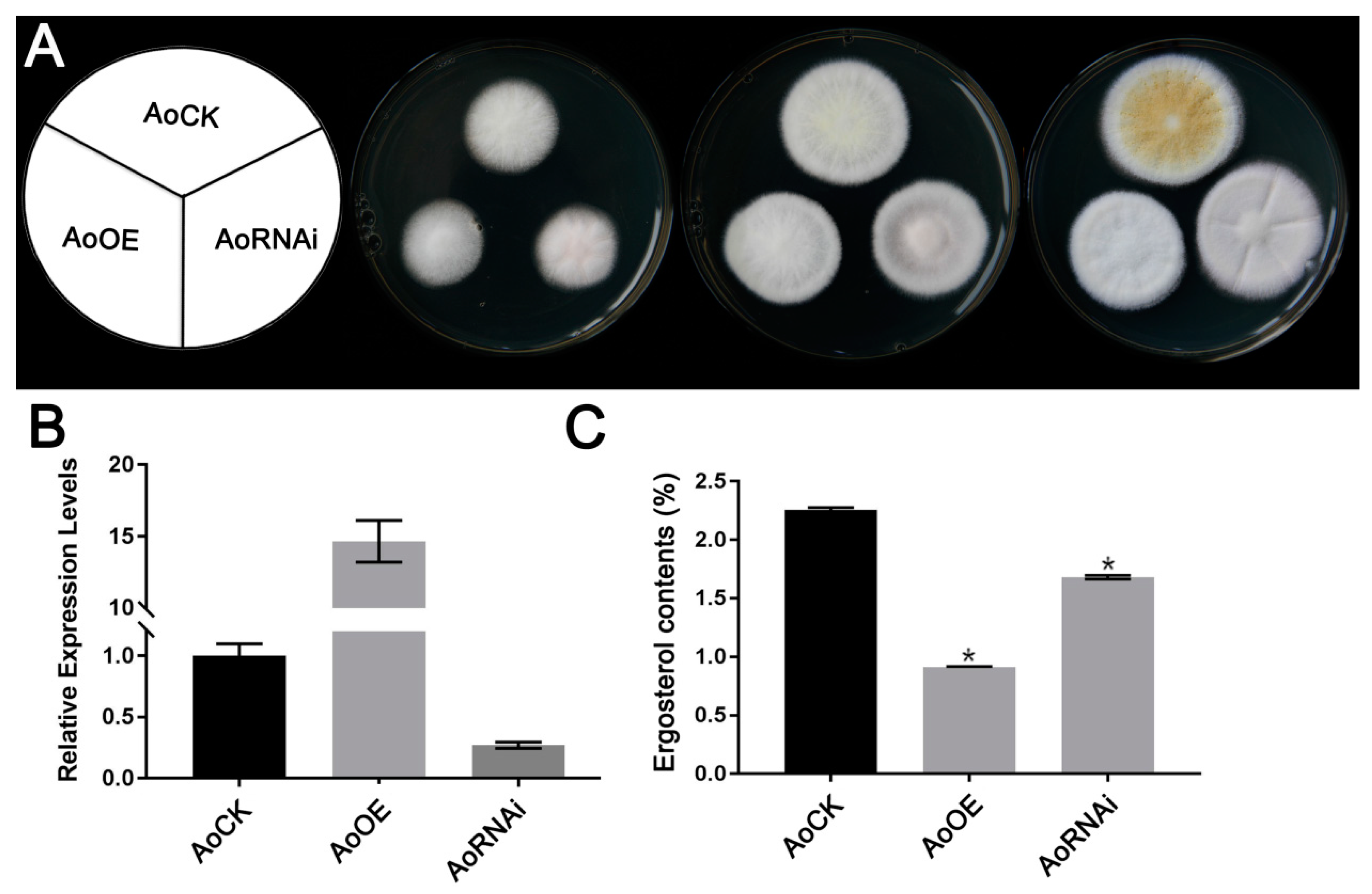
2.1. Phenotypic Characterization of A. oryzae AoErg19 Overexpression and RNAi Strains
2.2. Transcriptome Overview
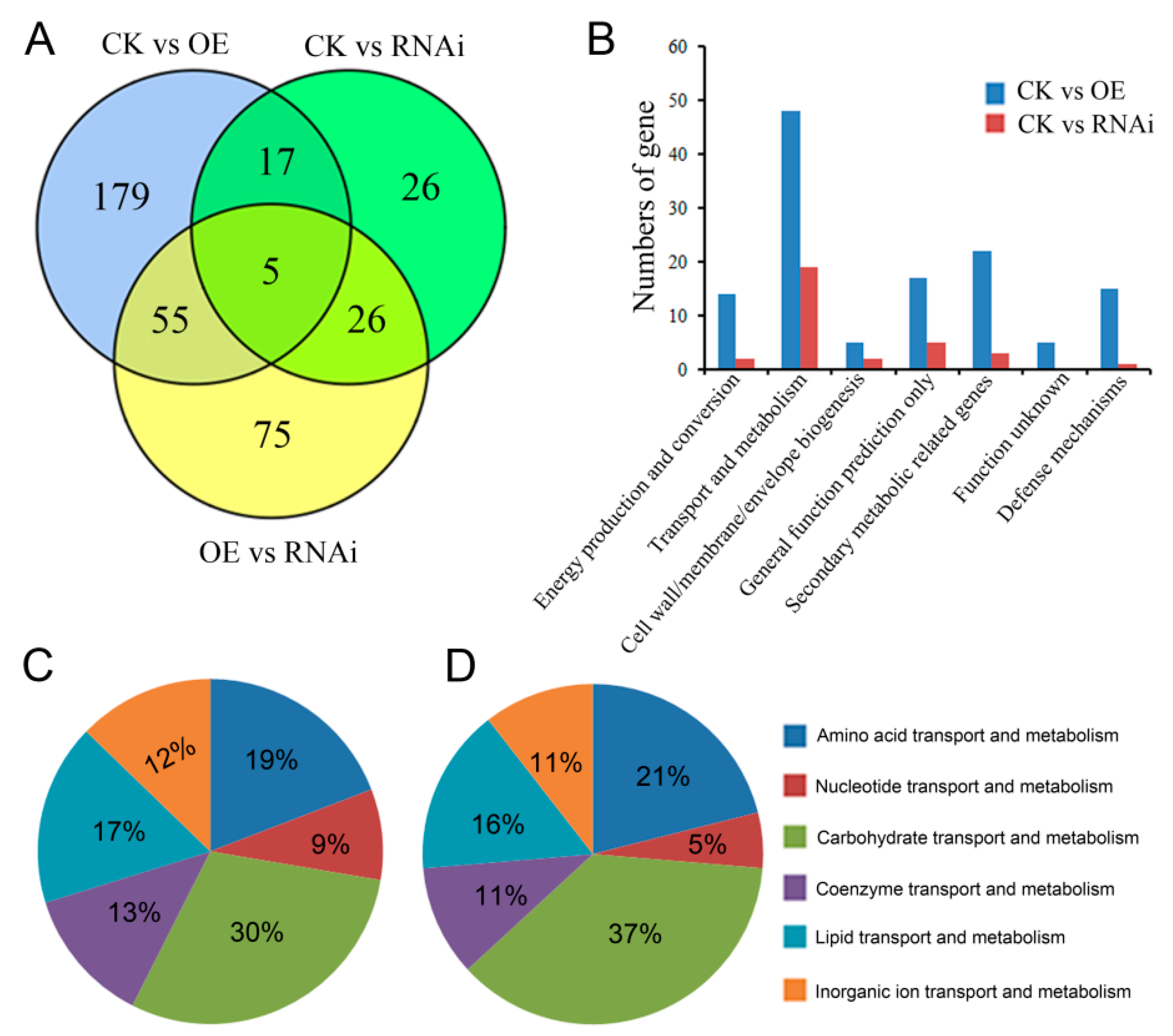
2.3. Differentially Expressed Genes in OE and RNAi Strains
2.4. Comparison of Gene Expression Profiles in OE and RNAi Strains
2.5. Expression of Cytochrome P450 Family Genes in the Over-Expression Strain
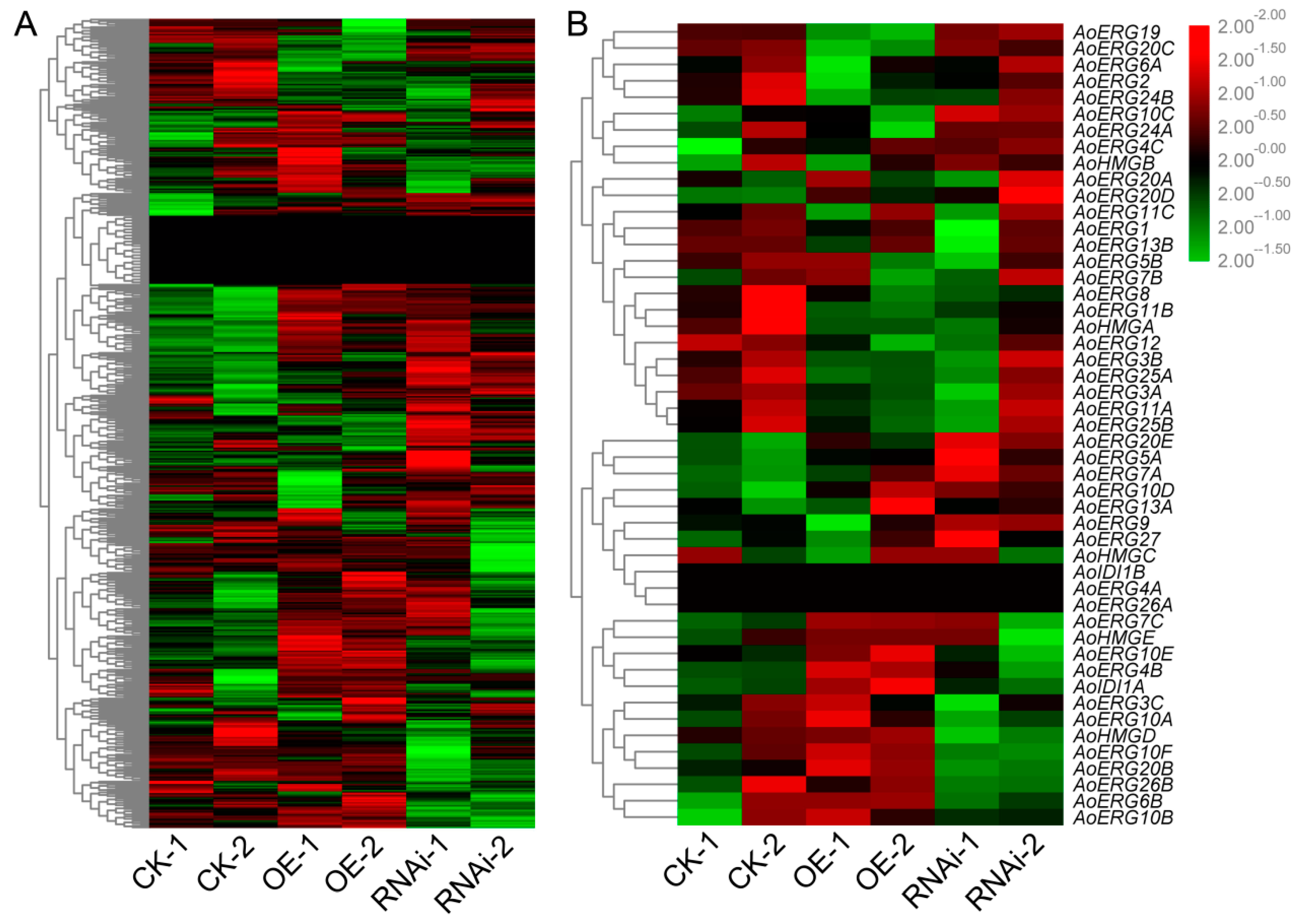
2.6. Expression Pattern of Genes Involved Lipid Biosynthesis and Metabolism
2.7. Fatty Acid Composition in Over Expression and RNAi A. oryzae Strains
3. Discussion
3.1. Effects of AoErg19 on Gene Transcription in A. oryzae
3.2. Comparison of Gene Expression Profiles in OE and RNAi Strains
3.3. Expression Pattern of Lipid Biosynthesis and Metabolism Genes, and Fatty Acid Content in OE and RNAi Strains
4. Method and Materials
4.1. Strain and Culture Conditions
4.2. Preparation of cDNA Libraries and RNA Sequencing
4.3. Mapping Reads to the A. oryzae Reference Genome and Normalized Gene Expressions
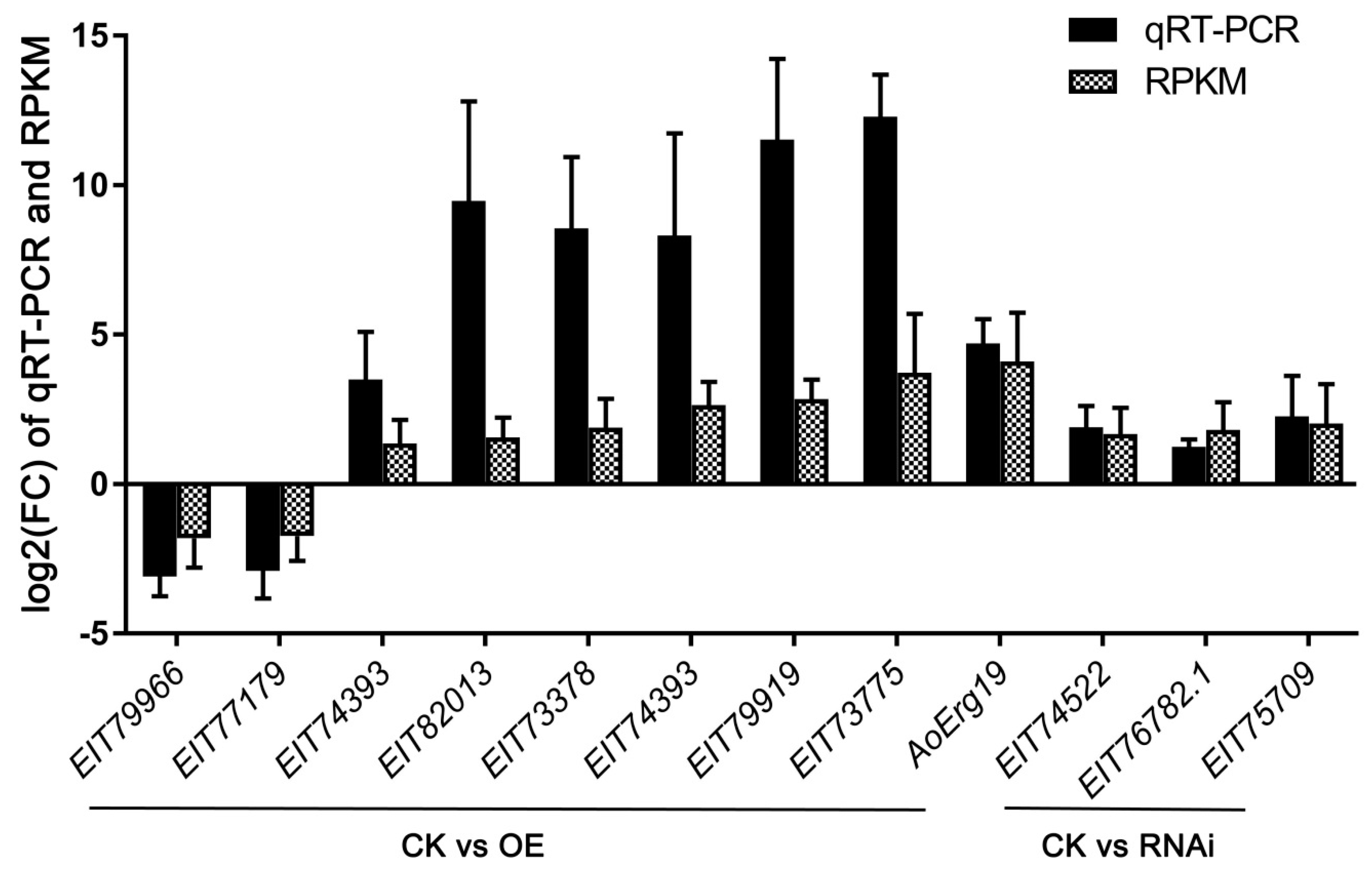
4.4. Quantitative Real-time Reverse Transcription–PCR
4.5. Measurement of Fatty Acid Contents
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hayakawa, H.; Sobue, F.; Motoyama, K.; Yoshimura, T.; Hemmi, H. Identification of enzymes involved in the mevalonate pathway of Flavobacterium johnsoniae. Biochem. Biophys. Res. Commun. 2017, 487, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.J.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.L. The potential of the mevalonate pathway for enhanced isoprenoid production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem.Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.M.; Garcia, D.E.; Reddingjohanson, A.M.; Friedland, G.D.; Chan, R.; Batth, T.S.; Haliburton, J.R.; Chivian, D.; Keasling, J.D.; Petzold, C.J. Optimization of a heterologous mevalonate pathway through the use of variant HMG-CoA reductases. Metab. Eng. 2011, 13, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; He, B.; Ma, L.; Sun, Y.; Niu, Y.; Zeng, B. Recent Advances in Ergosterol Biosynthesis and Regulation Mechanisms in Saccharomyces cerevisiae. Indian J. Microbiol. 2017, 57, 270–277. [Google Scholar] [CrossRef]
- Klug, L.; Daum, G. Yeast lipid metabolism at a glance. FEMS yeast Res. 2014, 14, 369–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krepkiy, D.; Miziorko, H.M. Identification of active site residues in mevalonate diphosphate decarboxylase: Implications for a family of phosphotransferases. Protein Sci. A Publ. Protein Soc. 2004, 13, 1875–1881. [Google Scholar] [CrossRef]
- Bergès, T.; Guyonnet, D.; Karst, F. The Saccharomyces cerevisiae mevalonate diphosphate decarboxylase is essential for viability, and a single Leu-to-Pro mutation in a conserved sequence leads to thermosensitivity. J. Bacteriol. 1997, 179, 4664–4670. [Google Scholar] [CrossRef] [Green Version]
- Kodedova, M.; Sychrova, H. Changes in the Sterol Composition of the Plasma Membrane Affect Membrane Potential, Salt Tolerance and the Activity of Multidrug Resistance Pumps in Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0139306. [Google Scholar] [CrossRef]
- Zhen, S.; Gary, R. The mevalonate pathway regulates microRNA activity in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2012, 109, 4568–4573. [Google Scholar]
- Abbassi, S.; Patel, K.; Khan, B.; Bhosale, S.; Gaikwad, S. Functional and conformational transitions of mevalonate diphosphate decarboxylase from Bacopa monniera. Int. J. Biol. Macromol. 2016, 83, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Brown, M.S. Regulation of the mevalonate pathway. Nature 1990, 343, 425. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, S.J.; Vishwakarma, R.K.; Patel, P.; Kumari, U.; Khan, B.M. Bacopa monniera recombinant mevalonate diphosphate decarboxylase: Biochemical characterization. Int. J. Biol. Macromol. 2015, 79, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aleem, S.; Nada, M.A.; Sayed-Ahmed, M.; Hendrickson, S.C.; Louis, J.S.; Walthall, H.P.; Lowe, J.E. Regulation of Fatty Acid Oxidation by Acetyl-CoA Generated from Glucose Utilization in Isolated Myocytes. J. Mol. Cell. Cardiol. 1996, 28, 825. [Google Scholar] [CrossRef] [PubMed]
- Sara, T.; Sidney, B.; Ute, S. De novo fatty acid biosynthesis and elongation in very long-chain acyl-CoA dehydrogenase-deficient mice supplemented with odd or even medium-chain fatty acids. Febs J. 2015, 282, 4242–4253. [Google Scholar]
- Raychaudhuri, S.; Young, B.P.; Espenshade, P.J.; Loewen, C., Jr. Regulation of lipid metabolism: A tale of two yeasts. Curr. Opin. Cell Biol. 2012, 24, 502–508. [Google Scholar] [CrossRef]
- Williams, M.L.; Menon, G.K.; Hanley, K.P. HMG-CoA reductase inhibitors perturb fatty acid metabolism and induce peroxisomes in keratinocytes. J. Lipid Res. 1992, 33, 193–208. [Google Scholar]
- Brandi, J.; Dando, I.; Pozza, E.D.; Biondani, G.; Jenkins, R.; Elliott, V.; Park, K.; Fanelli, G.; Zolla, L.; Costello, E.; et al. Proteomic analysis of pancreatic cancer stem cells: Functional role of fatty acid synthesis and mevalonate pathways. J. Proteom. 2017, 150, 310–322. [Google Scholar] [CrossRef]
- Yang, J.; Nie, Q.; Liu, H.; Xian, M.; Liu, H. A novel MVA-mediated pathway for isoprene production in engineered E. coli. BMC Biotechnol. 2016, 16, 5. [Google Scholar] [CrossRef]
- He, B.; Tu, Y.; Jiang, C.; Zhang, Z.; Li, Y.; Zeng, B. Functional Genomics of Aspergillus oryzae: Strategies and Progress. Microorganisms 2019, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Niu, Y.; Huang, H.; He, B.; Ma, L.; Tu, Y.; Tran, V.-T.; Zeng, B.; Hu, Z. Mevalonate Diphosphate Decarboxylase MVD/Erg19 Is Required for Ergosterol Biosynthesis, Growth, Sporulation and Stress Tolerance in Aspergillus oryzae. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Li, G.; Sun, Y.; Niu, Y.; Ma, L.; He, B.; Ai, M.; Han, J.; Zeng, B. Gene transcription profiling of Aspergillus oryzae 3.042 treated with ergosterol biosynthesis inhibitors. Braz. J. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jiang, J.; Shao, J.; Yin, Y.; Ma, Z. Gene transcription profiling of Fusarium graminearum treated with an azole fungicide tebuconazole. Appl. Microbiol. Biotechnol. 2010, 85, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Cirigliano, A.; Macone, A.; Bianchi, M.M.; Oliaro-Bosso, S.; Balliano, G.; Negri, R.; Rinaldi, T. Ergosterol reduction impairs mitochondrial DNA maintenance in S. cerevisiae. BBA-Mol. Cell Biol. Lipids 2019, 1864, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Ward, D.M.; Chen, O.S.; Li, L.; Kaplan, J.; Bhuiyan, S.A.; Natarajan, S.K.; Bard, M.; Cox, J.E. Altered sterol metabolism in budding yeast affects mitochondrial iron-sulfur (Fe-S) cluster synthesis. J. Biol. Chem. 2018, 293, 10782–10795. [Google Scholar] [CrossRef] [PubMed]
- Dimmer, K.S.; Fritz, S.; Fuchs, F.; Messerschmitt, M.; Weinbach, N.; Neupert, W.; Westermann, B. Genetic basis of mitochondrial function and morphology in Saccharomyces cerevisiae. Mol. Biol. Cell 2002, 13, 847. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Niu, Y.; He, B.; Ma, L.; Li, G.; Tran, V.T.; Zeng, B.; Hu, Z. A dual selection marker transformation system using Agrobacterium tumefaciens for the industrial Aspergillus oryzae 3.042. J. Microbiol. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Li, Z.Q.; Lei, B.; Cai, X.M.; Luo, Z.X.; Zhang, Y.J.; Chen, Z.M. Identification and Comparative Study of Chemosensory Genes Related to Host Selection by Legs Transcriptome Analysis in the Tea GeometridEctropis obliqua. PLoS ONE 2016, 11, e0149591. [Google Scholar]
- Audic, S.; Claverie, J.M. The Significance of Digital Gene Expression Profiles. Genome Res. 1997, 7, 986. [Google Scholar] [CrossRef] [PubMed]
- Blanc, G.; Gallot-Lavallée, L.; Maumus, F. Provirophages in the Bigelowiella genome bear testimony to past encounters with giant viruses. Proc. Natl. Acad. Sci. USA 2015, 112, 5318–5326. [Google Scholar] [CrossRef] [PubMed]
- Mannazzu, I.; Angelozzi, D.; Belviso, S.; Budroni, M.; Farris, G.A.; Goffrini, P.; Lodi, T.; Marzona, M.; Bardi, L. Behaviour of Saccharomyces cerevisiae wine strains during adaptation to unfavourable conditions of fermentation on synthetic medium: Cell lipid composition, membrane integrity, viability and fermentative activity. Int. J. Food Microbiol. 2008, 121, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Fu, L.; Hu, Z.; Li, Y.; Zheng, X.; Zhang, Z.; Jiang, C.; Zeng, B. Modulation of Fatty Acid Composition of Aspergillus oryzae in Response to Ethanol Stress. Microorganisms 2019, 7, 158. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tag Classification | WT | Over Expression | RNAi | |||
|---|---|---|---|---|---|---|
| 1 | 2 | 1 | 2 | 1 | 2 | |
| Total reads | 55,002,642 | 42,717,586 | 41,217,752 | 47,194,454 | 38,391,610 | 41,080,464 |
| Percentage of mapped reads | 89.33% | 91.69% | 88.29% | 87.58% | 86.95% | 87.83% |
| Percentage of unique reads | 89.04% | 91.38% | 88.02% | 87.27% | 87.27% | 87.51% |
| %≥Q30 | 91.75% | 92.19% | 92.17% | 91.96% | 91.61% | 92.59% |
| GC content | 52.58% | 52.58% | 52.86% | 52.75% | 52.78% | 52.75% |
| Pearson’s correlation coefficient | 0.919 | 0.98 | 0.767 | |||
| DEG Set | DEG Number | Up-Regulated | Down-Regulated |
|---|---|---|---|
| CK vs. OE | 256 | 148 | 108 |
| CK vs. RNAi | 74 | 41 | 33 |
| OE vs. RNAi | 161 | 87 | 74 |
| Accession Number | Putative Production Encoded by the Gene | Fold Changes | |
|---|---|---|---|
| OE | RNAi | ||
| Cell Membrane Protein or Transporter | |||
| EIT81161 | Integral component of membrane, function unknown | −3.14 | −2.95 |
| EIT72587 | Ca2+ transporting ATPase | −1.63 | −1.45 |
| EIT83492 | Transmembrane amino acid transporter protein | 4.48 | 4.67 |
| EIT76067 | Inorganic ion transport and metabolism | 4.48 | 1.87 |
| EIT82403 | Integral component of membrane | 2.55 | −4.15 |
| EIT83199 | Cellular membrane protein with unknown function | 2.19 | 2.79 |
| EIT79933 | Acyl-CoA hydrolase activity | −7.71 | −7.57 |
| EIT81552 | Phosphoribosylglycinamide synthetase | 1.59 | 1.53 |
| Energy production and conversion | |||
| EIT72870 | Mitochondrial protein FMP32 | 2.39 | 1.93 |
| EIT80752 | FAD-linked oxidoreductase | −4.30 | −4.44 |
| EIT83112 | FAD-dependent oxidoreductase | -Inf | -Inf |
| Transcription | |||
| EIT75959 | Demethylsterigmatocystin 6-O-methyltransferase | 3.48 | 5.32 |
| EIT77884 | RNA polymerase II transcription factor activity | −7.44 | −7.00 |
| EIT77860.1 | Transcriptional adapter 2 | −5.52 | −8.46 |
| Defense mechanisms | |||
| EIT76073 | Beta-lactamase class C | 5.30 | 1.81 |
| EIT77689 | Fungal fucose-specific lectin | −1.47 | 2.13 |
| Unknown function and others | |||
| EIT81216 | Hypothetical protein | −2.98 | −2.13 |
| EIT81230 | Hypothetical protein | −2.19 | −1.98 |
| EIT74353.1 | Hypothetical protein | 2.98 | 7.14 |
| EIT72406 | Hypothetical protein | −1.64 | −1.32 |
| EIT77940 | Tetratricopeptide repeat protein | −1.54 | −1.58 |
| EIT79995.1 | Alkyl sulfatase | 4.59 | 2.88 |
| Accession Number | CK vs. OE | CK vs. RNAi | OE vs. RNAi |
|---|---|---|---|
| EIT79521 | −3.76 | ND | ND |
| EIT79561 | −3.35 | ND | ND |
| EIT79542 | −3.25 | ND | ND |
| EIT79575 | −2.29 | ND | ND |
| EIT81599 | 1.09 | ND | ND |
| EIT80587 | 1.40 | ND | −2.48 |
| EIT73378 | 1.89 | ND | ND |
| EIT82715 | 2.32 | ND | ND |
| EIT83385 | 2.46 | ND | −2.35 |
| EIT82718 | 2.51 | ND | −1.53 |
| EIT81977 | 2.64 | ND | −3.28 |
| EIT81654 | 3.91 | ND | −2.91 |
| EIT80982 | ND | ND | 1.45 |
| EIT73302 | ND | ND | 6.47 |
| EIT81011 | ND | ND | 1.39 |
| EIT75636 | ND | ND | −2.27 |
| EIT81129 | ND | 1.58 | 1.18 |
| EIT80973 | ND | ND | 1.45 |
| EIT81014 | ND | ND | 1.43 |
| EIT80470 | ND | ND | 1.58 |
| EIT82011 | ND | ND | 3.48 |
| Accession Number | Putative Production Encoded by the Gene | Fold Changes |
|---|---|---|
| CK vs. OE | ||
| EIT79966 | Lanosterol synthase (AoErg7C) | −1.81 |
| EIT77179 | Phosphatidylserine decarboxylase activity | −1.73 |
| EIT74393 | Enoyl reductase domain of FAS1 | 1.36 |
| EIT82013 | Lipid transport and metabolism | 1.56 |
| EIT73378 | Sterol 14-demethylase (AoErg11B) | 1.89 |
| EIT74393 | Fatty acid synthase complex (AoFAT1) | 2.64 |
| EIT79919 | Sterol C-5 desaturases (AoErg3A) | 2.84 |
| EIT73775 | Oxidoreductase activity | 3.72 |
| EIT78501 | Diphosphomevalonate decarboxylase (AoErg19) | 4.10 |
| CK vs. RNAi | ||
| EIT74522 | Enoyl-(Acyl carrier protein) reductase | 1.67 |
| EIT76782.1 | Alpha-glucosidase | 1.81 |
| EIT75709 | Phosphogluconate dehydrogenase | 2.03 |
| Fatty Acid | CK | OE | RNAi |
|---|---|---|---|
| C14:0 | 0.014 ± 0.0031 | 0.047 ± 0.007* | 0.019 ± 0.003 |
| C15:0 | 0.242 ± 0.012 | 0.238 ± 0.013 | 0.249 ± 0.038 |
| C16:0 | 3.448 ± 0.810 | 3.523 ± 0.810 | 4.010 ± 0.910* |
| C16:1 | 0.061 ± 0.011 | 0.104 ± 0.012* | 0.059 ± 0.009 |
| C17:0 | 0.112 ± 0.023 | 0.120 ± 0.009 | 0.189 ± 0.031* |
| C18:0 | 0.183 ± 0.025 | 0.276 ± 0.0130* | 0.385 ± 0.016* |
| C18:1n9c | 2.102 ± 0.360 | 2.427 ± 0.190* | 2.054 ± 0.540 |
| C18:2n6c | 18.834 ± 1.890 | 17.773 ± 1.490 | 22.431 ± 1.970* |
| C20:0 | 0.006 ± 0.001 | 0.010 ± 0.001 | 0.012 ± 0.021* |
| C20:1 | 0.016 ± 0.003 | 0.022 ± 0.002 | 0.018 ± 0.003 |
| C18:3n3 | 1.214 ± 0.230 | 1.291 ± 0.190 | 2.015 ± 0.350* |
| C20:2 | 0.060 ± 0.011 | 0.059 ± 0.003 | 0.074 ± 0.019 |
| C22:0 | 0.008 ± 0.009 | 0.011 ± 0.003 | 0.021 ± 0.002 |
| C22:1n9 | 0.211 ± 0.013 | 0.291 ± 0.013 | 0.369 ± 0.018* |
| C20:3n3 | 0.019 ± 0.001 | 0.026 ± 0.019 | 0.040 ± 0.005* |
| C23:0 | 0.008 ± 0.001 | 0.011 ± 0.025 | 0.011 ± 0.004 |
| C22:2 | 0.016 ± 0.003 | 0.017 ± 0.003 | 0.020 ± 0.002 |
| C24:0 | 0.259 ± 0.013 | 0.324 ± 0.018* | 0.397 ± 0.071 |
| C24:1 | 0.008 ± 0.001 | 0.008 ± 0.001 | 0.009 ± 0.001 |
| UFA | 22.541 ± 2.523 | 22.018 ± 1.924 | 27.089 ± 2.916* |
| FA | 4.280 ± 0.887 | 4.560 ± 0.899 | 5.293 ± 1.096* |
| Total | 26.821 ± 3.410 | 26.578 ± 2.823 | 32.382 ± 4.012* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Huang, H.; Sun, Y.; Niu, Y.; Xu, W.; Liu, Q.; Zhang, Z.; Jiang, C.; Li, Y.; Zeng, B. Effects on Gene Transcription Profile and Fatty Acid Composition by Genetic Modification of Mevalonate Diphosphate Decarboxylase MVD/Erg19 in Aspergillus Oryzae. Microorganisms 2019, 7, 342. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090342
Hu Z, Huang H, Sun Y, Niu Y, Xu W, Liu Q, Zhang Z, Jiang C, Li Y, Zeng B. Effects on Gene Transcription Profile and Fatty Acid Composition by Genetic Modification of Mevalonate Diphosphate Decarboxylase MVD/Erg19 in Aspergillus Oryzae. Microorganisms. 2019; 7(9):342. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090342
Chicago/Turabian StyleHu, Zhihong, Hui Huang, Yunlong Sun, Yali Niu, Wangzishuai Xu, Qicong Liu, Zhe Zhang, Chunmiao Jiang, Yongkai Li, and Bin Zeng. 2019. "Effects on Gene Transcription Profile and Fatty Acid Composition by Genetic Modification of Mevalonate Diphosphate Decarboxylase MVD/Erg19 in Aspergillus Oryzae" Microorganisms 7, no. 9: 342. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090342