Enhanced Arsenic Tolerance in Triticum aestivum Inoculated with Arsenic-Resistant and Plant Growth Promoter Microorganisms from a Heavy Metal-Polluted Soil

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Isolation of Bacteria and Fungi and Their Resistance to Arsenic
2.3. Molecular Characterization
2.4. Screening of Potential Plant Growth-Promoting Traits
2.5. Establishment of Plants and Inoculations
2.6. Soil Biochemical Properties
2.7. Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. Bacteria and Fungi Identification and Arsenic Resistance
3.2. Screening of Potential Plant Growth-Promoting Traits
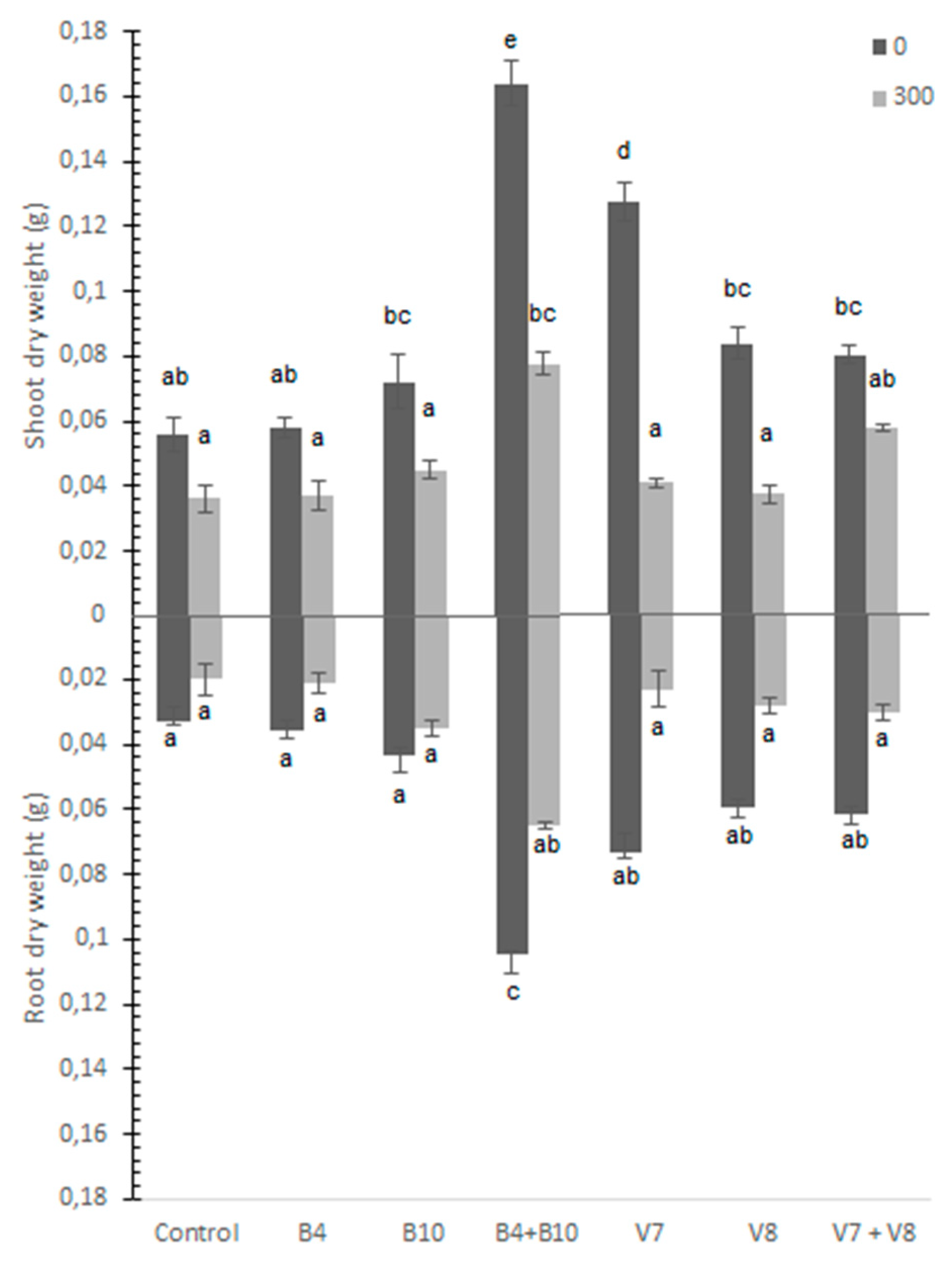
3.3. Shoot and Root Dry Biomass
3.4. Soil Enzyme Activities
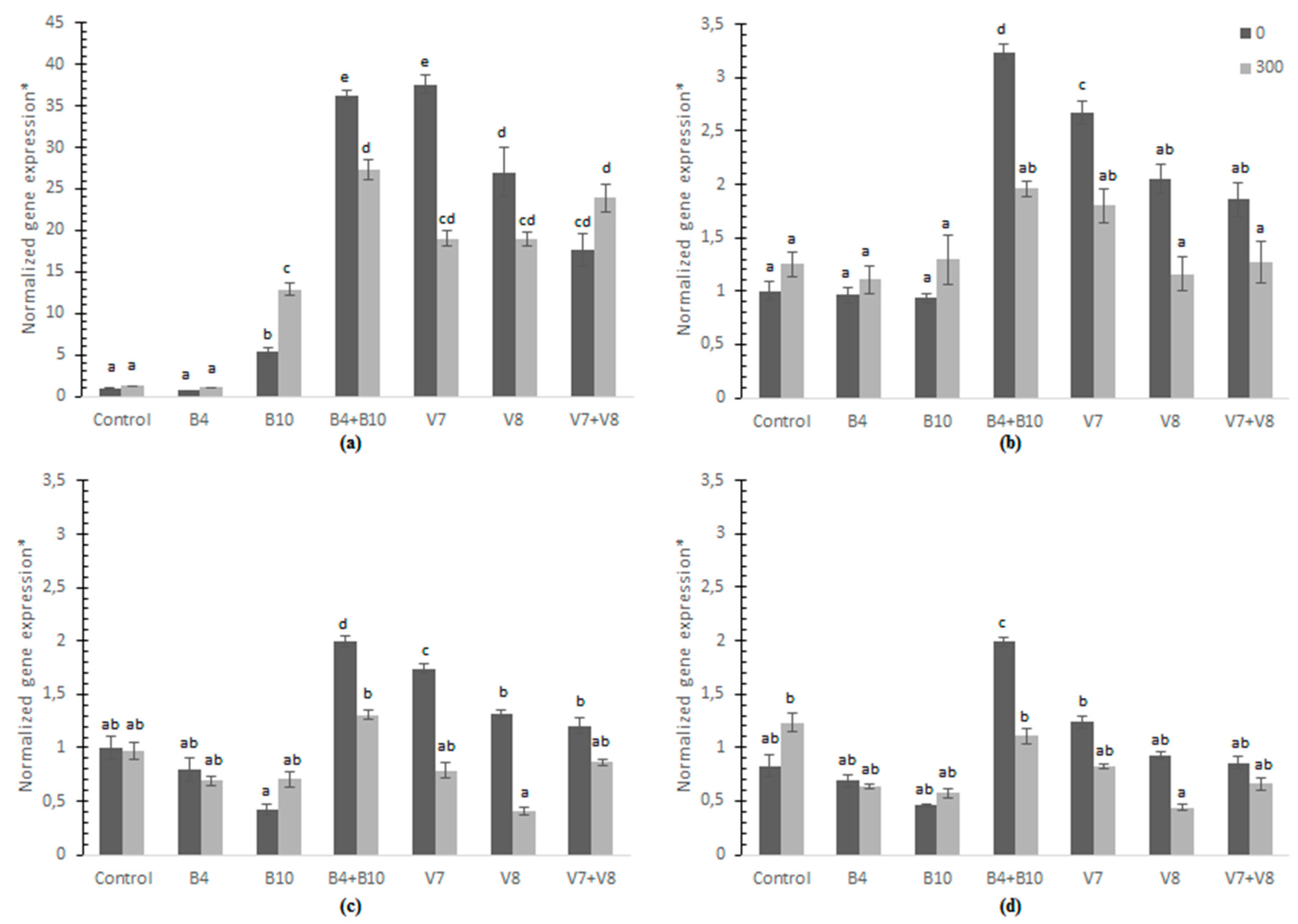
3.5. Relative Quantification of Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Arsenic Content (mg Kg−1) | Control | B4 | B10 | B4 + B10 | V7 | V8 | V7 + V8 |
|---|---|---|---|---|---|---|---|
| Shoot | |||||||
| Control | 0.37 | 0.51 | 0.41 | 0.6 | 0.52 | 0.45 | 0.42 |
| 300 mg Kg−1 | 8.83 | 6.79 | 6.13 | 6.11 | 7.13 | 6.07 | 6.17 |
| Root | |||||||
| Control | 0.407 | 0.5967 | 0.4879 | 0.72 | 0.5772 | 0.495 | 0.4998 |
| 300 mg Kg−1 | 9.0949 | 7.9443 | 7.7238 | 7.2709 | 8.2708 | 8.0124 | 8.2061 |
References
- Leist, M.; Casey, R.; Caridi, D. The management of arsenic wastes: Problems and prospects. J. Hazard. Mater. 2000, 76, 125–138. [Google Scholar] [CrossRef]
- Mandal, B.K.; Suzuki, K.T. Arsenic round the world: A review. Talanta 2002, 58, 201–235. [Google Scholar] [CrossRef]
- EPA U. Arsenic, inorganic (CASRN 7440-38-2). 2007. [Google Scholar]
- Smedley, P.; Kinniburgh, D. A review of the source, behaviour and distribution of arsenic in natural waters. Appl. Geochem. 2002, 17, 517–568. [Google Scholar] [CrossRef] [Green Version]
- Meharg, A. Integrated tolerance mechanisms: Constitutive and adaptive plant responses to elevated metal concentrations in the environment. Plant Cell Environ. 1994, 17, 989–993. [Google Scholar] [CrossRef]
- Kamiya, K.; Cruse, W.B.; Kennard, O. The arsonomethyl group as an analogue of phosphate. An X-ray investigation. Biochem. J. 1983, 213, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonell-Barrachina, A.; Burlo, F.; Burgos-Hernandez, A.; Lopez, E.; Mataix, J. The influence of arsenite concentration on arsenic accumulation in tomato and bean plants. Sci. Hortic. 1997, 71, 167–176. [Google Scholar] [CrossRef]
- Ahmed, F.S.; Killham, K.; Alexander, I. Influences of arbuscular mycorrhizal fungus Glomus mosseae on growth and nutrition of lentil irrigated with arsenic contaminated water. Plant Soil 2006, 283, 33. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K.; Suprasanna, P.; D’souza, S. Comparative biochemical and transcriptional profiling of two contrasting varieties of Brassica juncea L. in response to arsenic exposure reveals mechanisms of stress perception and tolerance. J. Exp. Bot. 2009, 60, 3419–3431. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Singla, P. Arsenic toxicity in crop plants: Physiological effects and tolerance mechanisms. Environ. Chem. Lett. 2011, 9, 303–321. [Google Scholar] [CrossRef]
- Finnegan, P.; Chen, W. Arsenic toxicity: The effects on plant metabolism. Front. Physiol. 2012, 3, 182. [Google Scholar] [CrossRef]
- Stoeva, N.; Bineva, T. Oxidative changes and photosynthesis in oat plants grown in As-contaminated soil. Bulg J. Plant Physiol 2003, 29, 87–95. [Google Scholar]
- Zhao, F.J.; McGrath, S.P.; Meharg, A.A. Arsenic as a food chain contaminant: Mechanisms of plant uptake and metabolism and mitigation strategies. Annu. Rev. Plant Biol. 2010, 61, 535–559. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Huang, H.; Yang, X.; Razafindrabe, B.; Inouhe, M. The detoxification of lead in Sedum alfredii H. is not related to phytochelatins but the glutathione. J. Hazard. Mater. 2010, 177, 437–444. [Google Scholar] [CrossRef]
- Wickes, W.; Wiskish, J. Arsenate uncoupling of oxidative phosphorylation in isolated plant mitochondria. Funct. Plant Biol. 1976, 3, 153–162. [Google Scholar] [CrossRef]
- Flora, S.J. Arsenic-induced oxidative stress and its reversibility. Free Radic. Biol. Med. 2011, 51, 257–281. [Google Scholar] [CrossRef]
- Sharma, I. Arsenic induced oxidative stress in plants. Biologia 2012, 67, 447–453. [Google Scholar] [CrossRef]
- Awasthi, S.; Chauhan, R.; Dwivedi, S.; Srivastava, S.; Srivastava, S.; Tripathi, R.D. A consortium of alga (Chlorella vulgaris) and bacterium (Pseudomonas putida) for amelioration of arsenic toxicity in rice: A promising and feasible approach. Environ. Exp. Bot. 2018, 150, 115–126. [Google Scholar] [CrossRef]
- Srivastava, S.; Singh, N. Mitigation approach of arsenic toxicity in chickpea grown in arsenic amended soil with arsenic tolerant plant growth promoting Acinetobacter sp. Ecol. Eng. 2014, 70, 146–153. [Google Scholar] [CrossRef]
- Pandey, N.; Bhatt, R. Role of soil associated Exiguobacterium in reducing arsenic toxicity and promoting plant growth in Vigna radiata. Eur. J. Soil Biol. 2016, 75, 142–150. [Google Scholar] [CrossRef]
- Hansda, A.; Kumar, V.; Anshumali, A.; Usmani, Z. Phytoremediation of heavy metals contaminated soil using plant growth promoting rhizobacteria (PGPR): A current perspective. Recent Res. Sci. Technol. 2014, 6. [Google Scholar]
- Glick, B.R. Using soil bacteria to facilitate phytoremediation. Biotechnol. Adv. 2010, 28, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Kuffner, M.; De Maria, S.; Puschenreiter, M.; Fallmann, K.; Wieshammer, G.; Gorfer, M.; Strauss, J.; Rivelli, A.; Sessitsch, A. Culturable bacteria from Zn-and Cd-accumulating Salix caprea with differential effects on plant growth and heavy metal availability. J. Appl. Microbiol. 2010, 108, 1471–1484. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yang, C.; Shi, S.; Shu, W. Effects of plant growth-promoting bacteria isolated from copper tailings on plants in sterilized and non-sterilized tailings. Chemosphere 2014, 97, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Ginocchio, R. Effects of a copper smelter on a grassland community in the Puchuncavı Valley, Chile. Chemosphere 2000, 41, 15–23. [Google Scholar] [CrossRef]
- Fuentes, A.; Almonacid, L.; Ocampo, J.A.; Arriagada, C. Synergistic interactions between a saprophytic fungal consortium and Rhizophagus irregularis alleviate oxidative stress in plants grown in heavy metal contaminated soil. Plant Soil 2016, 407, 355–366. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Olsen, S. Anion resin extractable phosphorus. Methods Soil Anal. 1982, 2, 423–424. [Google Scholar]
- Mingorance, M. Focused microwave-assisted digestion of vegetal materials for the determination of essential mineral nutrients. Anal. Bioanal. Chem. 2002, 373, 153–158. [Google Scholar] [CrossRef]
- Bissett, J.; Widden, P. An automatic, multichamber soil-washing apparatus for removing fungal spores from soil. Can. J. Microbiol. 1972, 18, 1399–1404. [Google Scholar] [CrossRef]
- Banerjee, S.; Datta, S.; Chattyopadhyay, D.; Sarkar, P. Arsenic accumulating and transforming bacteria isolated from contaminated soil for potential use in bioremediation. J. Environ. Sci. Health Part A 2011, 46, 1736–1747. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Herrera, H.; Valadares, R.; Contreras, D.; Bashan, Y.; Arriagada, C. Mycorrhizal compatibility and symbiotic seed germination of orchids from the Coastal Range and Andes in south central Chile. Mycorrhiza 2017, 27, 175–188. [Google Scholar] [CrossRef]
- Milagres, A.M.; Machuca, A.; Napoleao, D. Detection of siderophore production from several fungi and bacteria by a modification of chrome azurol S (CAS) agar plate assay. J. Microbiol. Methods 1999, 37, 1–6. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Khalid, A.; Arshad, M.; Zahir, Z. Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol. 2004, 96, 473–480. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Plant growth promoting activities of phosphate-solubilizing Enterobacter asburiae as influenced by fungicides. Eur. Asian. J. Bio. Sci. 2010, 4, 88–95. [Google Scholar] [CrossRef]
- Gianfreda, L.; Rao, M.A.; Piotrowska, A.; Palumbo, G.; Colombo, C. Soil enzyme activities as affected by anthropogenic alterations: Intensive agricultural practices and organic pollution. Sci. Total Environ. 2005, 341, 265–279. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Adam, G.; Duncan, H. Development of a sensitive and rapid method for the measurement of total microbial activity using fluorescein diacetate (FDA) in a range of soils. Soil Biol. Biochem. 2001, 33, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.; Fernández, F. O-CAS, a fast and universal method for siderophore detection. J. Microbiol. Methods 2007, 70, 127–131. [Google Scholar] [CrossRef]
- Rajkumar, M.; Ae, N.; Prasad, M.N.V.; Freitas, H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef]
- Yang, H.C.; Rosen, B.P. New mechanisms of bacterial arsenic resistance. Biomed. J. 2016, 39, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, P.K.; Vaish, A.; Dwivedi, S.; Chakrabarty, D.; Singh, N.; Tripathi, R.D. Biological removal of arsenic pollution by soil fungi. Sci. Total Environ. 2011, 409, 2430–2442. [Google Scholar] [CrossRef]
- Liao, V.H.C.; Chu, Y.J.; Su, Y.C.; Hsiao, S.Y.; Wei, C.C.; Liu, C.W.; Liao, C.M.; Shen, W.C.; Chang, F.J. Arsenite-oxidizing and arsenate-reducing bacteria associated with arsenic-rich groundwater in Taiwan. J. Contam. Hydrol. 2011, 123, 20–29. [Google Scholar] [CrossRef]
- Zhang, Z.; Yin, N.; Cai, X.; Wang, Z.; Cui, Y. Arsenic redox transformation by Pseudomonas sp. HN-2 isolated from arsenic-contaminated soil in Hunan, China. J. Environ. Sci. 2016, 47, 165–173. [Google Scholar] [CrossRef]
- Tsubouchi, T.; Kaneko, Y. Draft genome sequence of the arsenic-resistant bacterium Brevundimonas denitrificans TAR-002T. Genome Announc. 2017, 5, e01326-17. [Google Scholar] [CrossRef]
- Upadhyay, M.K.; Yadav, P.; Shukla, A.; Srivastava, S. Utilizing the potential of microorganisms for managing arsenic contamination: A feasible and sustainable approach. Front. Environ. Sci. 2018, 6, 24. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Jean, J.S.; Kar, S.; Chou, M.L.; Chen, C.Y. Screening of plant growth-promoting traits in arsenic-resistant bacteria isolated from agricultural soil and their potential implication for arsenic bioremediation. J. Hazard. Mater. 2014, 272, 112–120. [Google Scholar] [CrossRef]
- Xu, J.Y.; Han, Y.H.; Chen, Y.; Zhu, L.J.; Ma, L.Q. Arsenic transformation and plant growth promotion characteristics of As-resistant endophytic bacteria from As-hyperaccumulator Pteris vittata. Chemosphere 2016, 144, 1233–1240. [Google Scholar] [CrossRef]
- Carlos, M.H.J.; Stefani, P.V.Y.; Janette, A.M.; Melani, M.S.S.; Gabriela, P.O. Assessing the effects of heavy metals in ACC deaminase and IAA production on plant growth-promoting bacteria. Microbiol. Res. 2016, 188, 53–61. [Google Scholar] [CrossRef]
- Ma, Y.; Prasad, M.; Rajkumar, M.; Freitas, H. Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol. Adv. 2011, 29, 248–258. [Google Scholar] [CrossRef]
- Dale, S.E.; Doherty-Kirby, A.; Lajoie, G.; Heinrichs, D.E. Role of siderophore biosynthesis in virulence of Staphylococcus aureus: Identification and characterization of genes involved in production of a siderophore. Infect. Immun. 2004, 72, 29–37. [Google Scholar] [CrossRef]
- Wahyudi, A.T.; Astuti, R.P.; Widyawati, A.; Mery, A.; Nawangsih, A.A. Characterization of Bacillus sp. strains isolated from rhizosphere of soybean plants for their use as potential plant growth for promoting rhizobacteria. J. Microbiol. Antimicrob. 2011, 3, 34–40. [Google Scholar]
- Welbaum, G.E.; Sturz, A.V.; Dong, Z.; Nowak, J. Managing soil microorganisms to improve productivity of agro-ecosystems. Crit. Rev. Plant Sci. 2004, 23, 175–193. [Google Scholar] [CrossRef]
- Malik, J.A.; Goel, S.; Sandhir, R.; Nayyar, H. Uptake and distribution of arsenic in chickpea: Effects on seed yield and seed composition. Commun. Soil Sci. Plant Anal. 2011, 42, 1728–1738. [Google Scholar] [CrossRef]
- Shri, M.; Kumar, S.; Chakrabarty, D.; Trivedi, P.K.; Mallick, S.; Misra, P.; Shukla, D.; Mishra, S.; Srivastava, S.; Tripathi, R.D. Effect of arsenic on growth, oxidative stress, and antioxidant system in rice seedlings. Ecotoxicol. Environ. Saf. 2009, 72, 1102–1110. [Google Scholar] [CrossRef]
- Arriagada, C.; Aranda, E.; Sampedro, I.; Garcia-Romera, I.; Ocampo, J. Contribution of the saprobic fungi Trametes versicolor and Trichoderma harzianum and the arbuscular mycorrhizal fungi Glomus deserticola and G. claroideum to arsenic tolerance of Eucalyptus globulus. Bioresour. Technol. 2009, 100, 6250–6257. [Google Scholar] [CrossRef]
- Wang, S.; Fu, J. Insights into auxin signaling in plant–pathogen interactions. Front. Plant Sci. 2011, 2, 74. [Google Scholar]
- Ali, S.S.; Vidhale, N. Bacterial siderophore and their application: A review. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 303–312. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture—a review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Wang, Q.; Xiong, D.; Zhao, P.; Yu, X.; Tu, B.; Wang, G. Effect of applying an arsenic-resistant and plant growth–promoting rhizobacterium to enhance soil arsenic phytoremediation by Populus deltoides LH05-17. J. Appl. Microbiol. 2011, 111, 1065–1074. [Google Scholar] [CrossRef]
- Minamisawa, K.; Fukai, K. Production of indole-3-acetic acid by Bradyrhizobium japonicum: A correlation with genotype grouping and rhizobitoxine production. Plant Cell Physiol. 1991, 32, 1–9. [Google Scholar]
- Drewniak, L.; Styczek, A.; Majder-Lopatka, M.; Sklodowska, A. Bacteria, hypertolerant to arsenic in the rocks of an ancient gold mine, and their potential role in dissemination of arsenic pollution. Environ. Pollut. 2008, 156, 1069–1074. [Google Scholar] [CrossRef]
- Abercrombie, J.M.; Halfhill, M.D.; Ranjan, P.; Rao, M.R.; Saxton, A.M.; Yuan, J.S.; Stewart, C.N. Transcriptional responses of Arabidopsis thaliana plants to As (V) stress. BMC Plant Biol. 2008, 8, 87. [Google Scholar] [CrossRef]
- Leão, G.A.; de Oliveira, J.A.; Felipe, R.T.A.; Farnese, F.S.; Gusman, G.S. Anthocyanins, thiols, and antioxidant scavenging enzymes are involved in Lemna gibba tolerance to arsenic. J. Plant Interact. 2014, 9, 143–151. [Google Scholar] [CrossRef]


| pH (Water) | 5.54 ± 0.13 |
| Electrical conductivity (CE) (dS L−1) | 0.1 ± 0.09 |
| Organic matter (%) | 2.41 ± 0.16 |
| N total (%) | 0.1 ± 0.02 |
| N available (mg·kg−1) | 28.7 ± 1.78 |
| P available (mg·kg−1) | 40.3 ± 3.47 |
| K interchangeable (mg·kg−1) | 210 ± 8.21 |
| Cu total (mg·kg−1) | 385 ± 14.53 |
| Zn total (mg·kg−1) | 183 ± 7.93 |
| Cd total (mg·kg−1) | 1.1 ± 0.07 |
| Pb total (mg·kg−1) | 135 ± 6.91 |
| As total (mg·kg−1) | 52 ± 3.65 |
| Gene | Function | Primers 5′–3′ | Amplicon Length (pb) | |
|---|---|---|---|---|
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | F | CCGTGTTCCCACTGTTGATGTT | 192 |
| R | GCATCAAAGATGCTGGACCTGT | |||
| α-TUB | Alpha-tubulin | F | CTGACAGCTTCCCTGAGGTTTGAT | 179 |
| R | TCAAAGGCGCTGTTGGTGATCT | |||
| PCS | Phytochelatin synthase | F | GCTATGTGGTAGTTGCTCGTCTTC | 195 |
| R | ACCACGGTTCCTGAGATAACAGTC | |||
| EF1α | Elongation factor-1 alpha | F | AGGCTGTCCGCAGTGTTCAAAT | 178 |
| R | TCACACGACTGGACATACTCGTTG | |||
| APX | Ascorbate peroxidase | F | TCCAACCGTTGAGTTCATCCCT | 199 |
| R | ACCGTCAAACCCAGACCTTTCA | |||
| Cu/Zn SOD | Cu/Zn Superoxide dismutase | F | TTTCCAGTCGCTCCGAATTGTCTC | 186 |
| R | AGTCCAGTGATACGAACGTTCACC | |||
| MT | Metallothionein | F | CCAGTGCAGATCAGTATCAGACCA | 176 |
| R | CTCGTCCATCTCAGGGTACATCTT |
| Strains | Molecular Identification | Close Match Accession Number | MIC (mg·L−1) | IAA As (mg·L−1) | Siderophore Production † | Phosphate Solubilization ‡ | ||
|---|---|---|---|---|---|---|---|---|
| AsV | AsIII | 0 | 250 | |||||
| B4 | Pseudomonas gessardii | MH398505.1 | 7000 | 1500 | 0 | 0 | + | + |
| B10 | Brevundimonas intermedia | KR811205.1 | 6000 | 1500 | 52.32 ± 0.9 a | 52.45 ± 0.3 a | − | − |
| V7 | Fimetariella rabenhortii | HQ406808 | 2500 | 750 | 40.5 ± 0.74 b | 41.6 ± 0.42 b | − | − |
| V8 | Hormonema viticola | NR137620.1 | 2500 | 720 | 57.64 ±1.06 a | 56.62 ± 2.78 a | − | − |
| β-Gucosidase(µmoles p-Nitrophenol g−1 h−1) | FDA (µg Fluorescein g−1) | Acid Phosphatase (µmoles p-Nitrophenol g−1 h−1) | |
|---|---|---|---|
| Soil without As Added | |||
| Control | 96.01 ± 8.23 | 12.8 ± 1.17 | 82.09 ± 8.01 a,b,c |
| B4 | 84.39 ± 11.41 | 13.02 ± 1.05 | 90.06 ± 6.18 a,b,c,d |
| B10 | 106.21 ± 13.45 | 14.03 ± 3.17 | 128.37 ± 2.88 d,e,f |
| B4 + B0 | 121.35 ± 10.19 | 14.14 ± 1.65 | 123.78 ± 15.08 c,d,e,f |
| V7 | 134.32 ± 4.3 | 15.4 ± 1.3 | 181.95 ± 6.53 g,h |
| V8 | 97.7 ± 7.43 | 12.24 ± 0.93 | 113.37 ± 4.66 c,d,e,f |
| V7 + V8 | 123.1 ± 10.1 | 11.02 ± 0.97 | 201.48 ± 10.42 h |
| Soil with As Added | |||
| Control | 99.32 ± 21.27 | 12.15 ± 1.74 | 60.06 ± 5.57 a |
| B4 | 109.18 ± 21 | 12.3 ± 0.85 | 70.67 ± 5.4 a,b |
| B10 | 144.32 ± 18.54 | 13.45 ± 1.36 | 150 ± 4.89 e,f,g |
| B4 + B0 | 114.25 ± 15.63 | 13.91 ± 2.77 | 109.72 ± 6.28 b,c,d,e |
| V7 | 109.32 ± 1.4 | 11.68 ± 0.52 | 96.41 ± 7.38 a,b,c,d |
| V8 | 138.51 ± 8.41 | 16.8 ± 1.99 | 158.01 ± 17.62 f,g,h |
| V7 + V8 | 117.97 ± 6.75 | 15.75 ± 1.5 | 114.39 ± 8.33 c,d,e,f |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto, J.; Ortiz, J.; Herrera, H.; Fuentes, A.; Almonacid, L.; Charles, T.C.; Arriagada, C. Enhanced Arsenic Tolerance in Triticum aestivum Inoculated with Arsenic-Resistant and Plant Growth Promoter Microorganisms from a Heavy Metal-Polluted Soil. Microorganisms 2019, 7, 348. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090348
Soto J, Ortiz J, Herrera H, Fuentes A, Almonacid L, Charles TC, Arriagada C. Enhanced Arsenic Tolerance in Triticum aestivum Inoculated with Arsenic-Resistant and Plant Growth Promoter Microorganisms from a Heavy Metal-Polluted Soil. Microorganisms. 2019; 7(9):348. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090348
Chicago/Turabian StyleSoto, Javiera, Javier Ortiz, Hector Herrera, Alejandra Fuentes, Leonardo Almonacid, Trevor C. Charles, and César Arriagada. 2019. "Enhanced Arsenic Tolerance in Triticum aestivum Inoculated with Arsenic-Resistant and Plant Growth Promoter Microorganisms from a Heavy Metal-Polluted Soil" Microorganisms 7, no. 9: 348. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090348