Horizontal Gene Transfer and Its Association with Antibiotic Resistance in the Genus Aeromonas spp.

 ,
,  , ,
, ,  and
and 
{kind=link}
Abstract
:1. Introduction
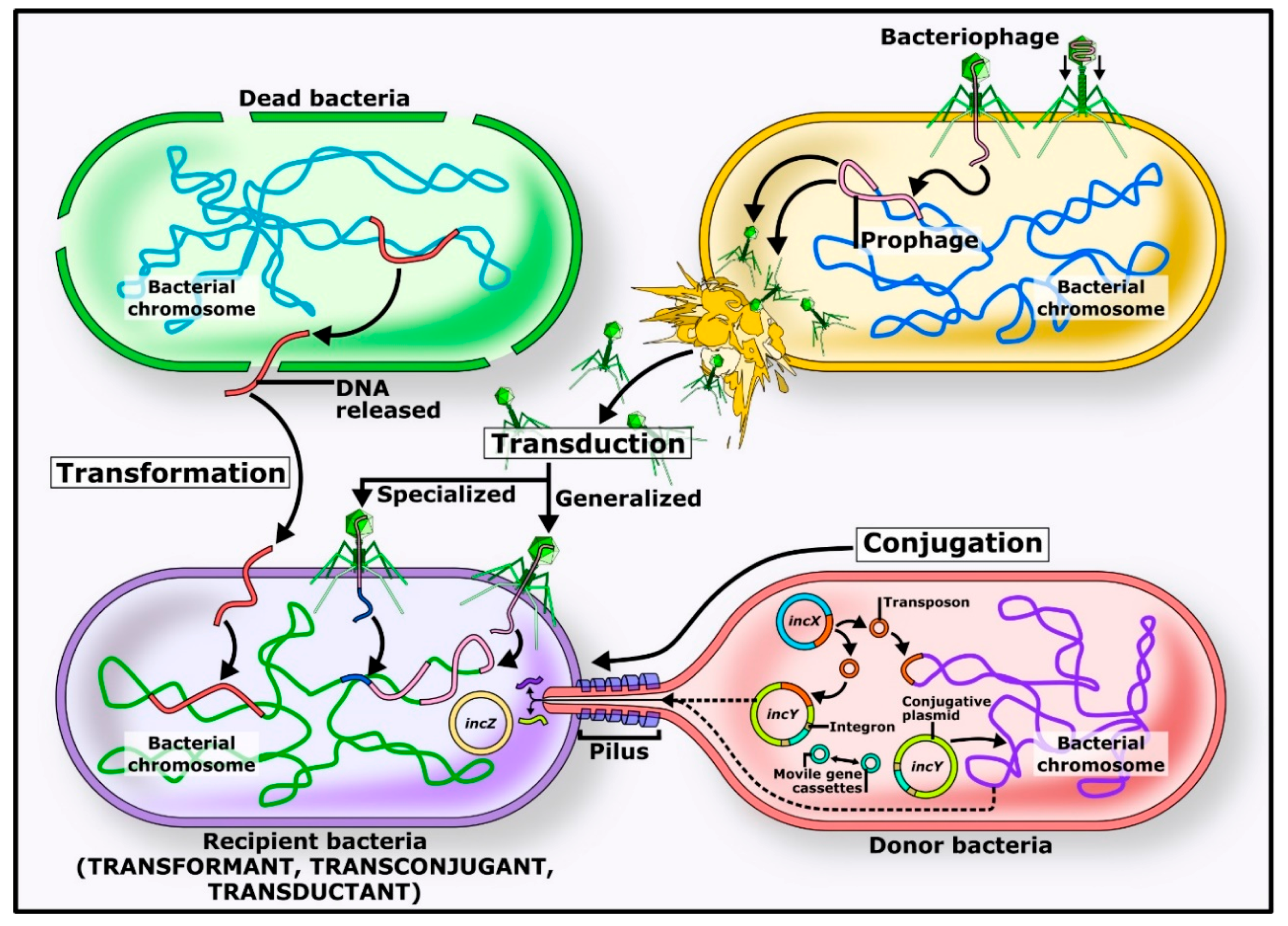
2. Horizontal Transfer Mechanisms
2.1. Transformation
2.2. Transduction
2.3. Conjugation
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Do, T.T.; Murphy, S.; Walsh, F. Antibiotic Resistance and Wastewater Treatment Process. Antimicrob. Resist. Wastewater Treat. Process. 2017, 263–291. [Google Scholar] [CrossRef]
- Figueras, M.J.; Alperi, A.; Beaz-Hidalgo, R.; Stackebrandt, E.; Brambilla, E.; Monera, A.; Martínez-Murcia, A.J. Aeromonas rivuli sp. nov., isolated from the upstream region of a karst water rivulet. Int. J. Syst. Evol. Microbiol. 2011, 61, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Aravena-Román, M.; Beaz-Hidalgo, R.; Inglis, T.J.J.; Riley, T.V.; Martínez-Murcia, A.J.; Chang, B.J.; Figueras, M.J. Aeromonas australiensis sp. nov., isolated from irrigation water. Int. J. Syst. Evol. Microbiol. 2013, 63, 2270–2276. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Murcia, A.; Beaz-Hidalgo, R.; Svec, P.; Saavedra, M.J.; Figueras, M.J.; Sedlacek, I. Aeromonas cavernicola sp. nov., isolated from fresh water of a brook in a cavern. Curr. Microbiol. 2013, 66, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Figueras, M.J.; Latif-Eugenín, F.; Ballester, F.; Pujol, I.; Tena, D.; Berg, K.; Hossain, M.J.; Beaz-Hidalgo, R.; Liles, M.R. ‘Aeromonas intestinalis’ and ‘Aeromonas enterica’ isolated from human faeces, ‘Aeromonas crassostreae’ from oyster and ‘Aeromonas aquatilis’ isolated from lake water represent novel species. New Microbes New Infect. 2017, 15, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Martínez-Murcia, A. Phylogenetic analyses of the genus Aeromonas based on housekeeping gene sequencing and its influence on systematics. J. Appl. Microbiol. 2018, 125, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves Pessoa, R.B.; de Oliveira, W.F.; Marques, D.S.C.; dos Santos Correia, M.T.; de Carvalho, E.V.M.M.; Coelho, L.C.B.B. The genus Aeromonas: A general approach. Microb. Pathog. 2019, 130, 81–94. [Google Scholar] [CrossRef]
- Dongyou, L. Molecular Medical Microbiology, 2nd ed.; Academic Press: New York, NY, USA, 2015. [Google Scholar]
- Percival, S.L.; Williams, D.W. Microbiology of Waterborne Diseases, 2nd ed.; Academic Press: New York, NY, USA, 2014. [Google Scholar]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. Sci. World J. 2012, 2012, 1–13. [Google Scholar]
- Lowry, R.; Balboa, S.; Parker, J.L.; Shaw, J.G. Aeromonas Flagella and Colonisation Mechanisms. Adv. Microb. Physiol. 2014, 65, 203–256. [Google Scholar]
- Martínez, J.L.; Coque, T.M.; Lanza, V.F.; de la Cruz, F.; Baquero, F. Genomic and metagenomic technologies to explore the antibiotic resistance mobilome. Ann. N. Y. Acad. Sci. 2017, 1388, 26–41. [Google Scholar] [CrossRef]
- Gillings, M.R. Evolutionary consequences of antibiotic use for the resistome, mobilome, and microbial pangenome. Front. Microbiol. 2013, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Gao, G.F.; Zhu, B. The antibiotic resistome: Gene flow in environments, animals and human beings. Front. Med. 2017, 11, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Wright, G.D. The antibiotic resistance ‘mobilome’: Searching for the link between environment and clinic. Front. Microbiol. 2013, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Torres Manrique, C. La Resistencia Bacteriana A Los Antibióticos, Siete Décadas Después De Fleming; Colegio Oficial de Farmacéuticos de Zaragoza: Zaragoza, Spain, 2012. [Google Scholar]
- Wright, G.D. The antibiotic resistome. Expert Opin. Drug Discov. 2010, 5, 779–788. [Google Scholar] [CrossRef]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Galán, J.C.; González-Candelas, F.; Rolain, J.M.; Cantón, R. Antibiotics as selectors and accelerators of diversity in the mechanisms of resistance: From the resistome to genetic plasticity in the β-lactamases world. Front. Microbiol. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, R.; Cantón, R.; Brown, D.F.J.; Giske, C.G.; Heisig, P.; MacGowan, A.P.; Mouton, J.W.; Nordmann, P.; Rodloff, A.C.; Rossolini, G.M.; et al. EUCAST expert rules in antimicrobial susceptibility testing. Clin. Microbiol. Infect. 2013, 19, 141–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekunberri, I.; Balcázar, J.L.; Borrego, C.M. Metagenomic exploration reveals a marked change in the river resistome and mobilome after treated wastewater discharges. Env. Pollut. 2018, 234, 538–542. [Google Scholar] [CrossRef]
- Norman, A.; Hansen, L.H.; Sørensen, S.J. Conjugative plasmids: Vessels of the communal gene pool. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2275–2289. [Google Scholar] [CrossRef]
- Thomas, C.M. Gene Recruiters and Transporters: The Modular Structure of Bacterial Mobile Elements. Horizontal Gene Pool; CRC Press: London, UK, 2000; pp. 388–443. [Google Scholar]
- Shintani, M. The behavior of mobile genetic elements (MGEs) in different environments. Biosci. Biotechnol. Biochem. 2017, 81, 854–862. [Google Scholar] [CrossRef] [Green Version]
- Sitaraman, R. Prokaryotic horizontal gene transfer within the human holobiont: Ecological-evolutionary inferences, implications and possibilities. Microbiome 2018, 6, 163. [Google Scholar] [CrossRef] [PubMed]
- Mandujano, A.G.; Lopez, V.E.G.; Longa, B.A.; Castro, E. Production of Outer Membrane Vesicles in a Clinical Strain of Aeromonas hydrophila. EC Microbiol. 2005, 1, 113–117. [Google Scholar]
- Hasegawa, Y.; Futamata, H.; Tashiro, Y. Complexities of cell-to-cell communication through membrane vesicles: Implications for selective interaction of membrane vesicles with microbial cells. Front. Microbiol. 2015, 6, 633. [Google Scholar] [CrossRef] [PubMed]
- Dubey, G.P.; Ben-Yehuda, S. Intercellular nanotubes mediate bacterial communication. Cell 2011, 144, 590–600. [Google Scholar] [CrossRef] [PubMed]
- García-Aljaro, C.; Ballesté, E.; Muniesa, M. Beyond the canonical strategies of horizontal gene transfer in prokaryotes. Curr. Opin. Microbiol. 2017, 38, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef]
- Skippington, E.; Ragan, M.A. Lateral genetic transfer and the construction of genetic exchange communities. Fems Microbiol. Rev. 2011, 35, 707–735. [Google Scholar] [CrossRef]
- Lorenz, M.G.; Wackernagel, W. Bacterial gene transfer by natural genetic transformation in the environment. Microbiol. Rev. 1994, 58, 563–602. [Google Scholar]
- Silver, A.C.; Williams, D.; Faucher, J.; Horneman, A.J.; Gogarten, J.P.; Graf, J. Complex Evolutionary History of the Aeromonas veronii Group Revealed by Host Interaction and DNA Sequence Data. PLoS ONE 2011, 6, e16751. [Google Scholar] [CrossRef]
- Huddleston, J.R.; Brokaw, J.M.; Zak, J.C.; Jeter, R.M. Natural transformation as a mechanism of horizontal gene transfer among environmental Aeromonas species. Syst. Appl. Microbiol. 2013, 36, 224–234. [Google Scholar] [CrossRef]
- Dallaire-Dufresne, S.; Emond-Rheault, J.G.; Attéré, S.A.; Tanaka, K.H.; Trudel, M.V.; Frenette, M.; Charette, S.J. Optimization of a plasmid electroporation protocol for Aeromonas salmonicida subsp. salmonicida. J. Microbiol. Methods 2014, 98, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Fengqing, H.; Song, Y. Electroporation-mediated transformation of Aeromonas hydrophila. Plasmid 2005, 54, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Nováková, J.; Izsáková, A.; Grivalský, T.; Ottmann, C.; Farkašovský, M. Improved method for high-efficiency electrotransformation of Escherichia coli with the large BAC plasmids. Folia Microbiol. 2014, 59, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.H.; Kumar, A.; Schweizer, H.P. A 10-min method for preparation of highly electrocompetent Pseudomonas aeruginosa cells: Application for DNA fragment transfer between chromosomes and plasmid transformation. J. Microbiol. Methods 2006, 64, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Arber, W. Horizontal Gene Transfer among Bacteria and Its Role in Biological Evolution. Life 2014, 4, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juhas, M. Horizontal gene transfer in human pathogens. Crit. Rev. Microbiol. 2015, 41, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Chen, L.; Liu, Q.; Zhou, Y.; Yang, J.; Deng, D.; Li, H.; Ma, Y. Characterization and genomic analyses of Aeromonas hydrophila phages AhSzq-1 and AhSzw-1, isolates representing new species within the T5 virus genus. Arch. Virol. 2018, 163, 1985–1988. [Google Scholar] [CrossRef] [PubMed]
- Le, T.S.; Nguyen, T.H.; Vo, H.P.; Doan, V.C.; Nguyen, H.L.; Tran, M.T.; Tran, T.T.; Southgate, P.C.; Kurtböke, D.İ. Protective Effects of Bacteriophages against Aeromonas hydrophila Causing Motile Aeromonas Septicemia (MAS) in Striped Catfish. Antibiotics 2018, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Paquet, V.E.; Bernatchez, A.; Tremblay, D.M.; Moineau, S.; Charette, S.J. Characterization and diversity of phages infecting Aeromonas salmonicida subsp. salmonicida. Sci. Rep. 2017, 7, 7054. [Google Scholar] [CrossRef]
- Jiang, S.C.; Paul, J.H. Gene transfer by transduction in the marine environment. Appl. Env. Microbiol. 1998, 64, 2780–2787. [Google Scholar]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Guglielmetti, E.; Korhonen, J.M.; Heikkinen, J.; Morelli, L.; Von Wright, A. Transfer of plasmid-mediated resistance to tetracycline in pathogenic bacteria from fish and aquaculture environments. Fems Microbiol. Lett. 2009, 293, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile genetic elements: The agents of open source evolution. Nat. Rev. Microbiol. 2005, 3, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P. Plasmid incompatibility. Microbiol. Rev. 1987, 51, 381–395. [Google Scholar] [CrossRef]
- Ilangovan, A.; Connery, S.; Waksman, G. Structural biology of the Gram-negative bacterial conjugation systems. Trends Microbiol. 2015, 23, 301–310. [Google Scholar] [CrossRef]
- Bello-López, J.M.; Vázquez-Ocampo, N.J.; Fernández-Rendón, E.; Curiel-Quesada, E. Inability of some Aeromonas hydrophila strains to act as recipients of plasmid pRAS1 in conjugal transfer experiments. Curr. Microbiol. 2012, 64, 332–337. [Google Scholar] [CrossRef]
- Bello-López, J.M.; Fernández-Rendón, E.; Curiel-Quesada, E. In vivo transfer of plasmid pRAS1 between Aeromonas salmonicida and Aeromonas hydrophila in artificially infected Cyprinus carpio L. J. Fish Dis. 2010, 33, 251–259. [Google Scholar] [CrossRef]
- Sandt, C.H.; Herson, D.S. Mobilization of the genetically engineered plasmid pHSV106 from Escherichia coli HB101(pHSV106) to Enterobacter cloacae in drinking water. Appl. Env. Microbiol. 1991, 57, 194–200. [Google Scholar]
- Hernández–Montañez, Z.; López–Ramírez, M.P.; Delgado–Balbuena, L.; Dendooven, L.; Bello–López, J.M. Mesophilic strains of Aeromonas spp. can acquire the multidrug resistance plasmid pRAS1 in horizontal transfer experiments at low temperatures. Ann. Microbiol. 2015, 65, 827–831. [Google Scholar] [CrossRef]
- Matsushita, M.; Okubo, T.; Hasegawa, T.; Matsuo, J.; Watanabe, T.; Iwasaki, S.; Fukumoto, T.; Hayasaka, K.; Akizawa, K.; Shimizu, C.; et al. Tetrahymena promotes interactive transfer of carbapenemase gene encoded in plasmid between fecal Escherichia coli and environmental Aeromonas caviae. Microbiol. Immunol. 2018, 62, 720–728. [Google Scholar] [CrossRef]
- Cabrero-Martínez, O.A.; Palma Nicolas, J.P.; Martínez-Vázquez, A.V.; Rivera Sánchez, G.; Bocanegra-García, V. Free-living amoebae detection by culture in superficial water sources in the city of Reynosa, Tamaulipas. Mex. J. Biotechnol. 2018, 3, 16–22. [Google Scholar] [CrossRef]
- Sørum, H.; L’Abée-Lund, T.M. Antibiotic resistance in food-related bacteria—A result of interfering with the global web of bacterial genetics. Int. J. Food Microbiol. 2002, 78, 43–56. [Google Scholar] [CrossRef]
- Rhodes, G.; Huys, G.; Swings, J.; McGann, P.; Hiney, M.; Smith, P.; Pickup, R.W. Distribution of oxytetracycline resistance plasmids between aeromonads in hospital and aquaculture environments: Implication of Tn1721 in dissemination of the tetracycline resistance determinant Tet, A. Appl. Env. Microbiol. 2000, 66, 3883–3890. [Google Scholar] [CrossRef] [PubMed]
- Scarano, C.; Piras, F.; Virdis, S.; Ziino, G.; Nuvoloni, R.; Dalmasso, A.; De Santis, E.P.L.; Spanu, C. Antibiotic resistance of Aeromonas ssp. strains isolated from Sparus aurata reared in Italian mariculture farms. Int. J. Food Microbiol. 2018, 284, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Popowska, M. Insight into the mobilome of Aeromonas strains. Front. Microbiol. 2015, 6, 494. [Google Scholar] [CrossRef]
- Jones, B.L.; Wilcox, M.H. Aeromonas infections and their treatment. J. Antimicrob. Chemother. 1995, 35, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Batra, P.; Mathur, P.; Misra, M. Aeromonas spp.: An emerging nosocomial pathogen. J. Lab. Phys. 2016, 8, 1. [Google Scholar]
- Sen, K.; Rodgers, M. Distribution of six virulence factors in Aeromonas species isolated from US drinking water utilities: A PCR identification. J. Appl. Microbiol. 2004, 97, 1077–1086. [Google Scholar] [CrossRef]
- Ghatak, S.; Blom, J.; Das, S.; Sanjukta, R.; Puro, K.; Mawlong, M.; Shakuntala, I.; Sen, A.; Goesmann, A.; Kumar, A.; et al. Pan-genome analysis of Aeromonas hydrophila, Aeromonas veronii and Aeromonas caviae indicates phylogenomic diversity and greater pathogenic potential for Aeromonas hydrophila. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2016, 109, 945–956. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef]
- Wu, C.J.; Chuang, Y.C.; Lee, M.F.; Lee, C.C.; Lee, H.C.; Lee, N.Y.; Chang, C.M.; Chen, P.L.; Lin, Y.T.; Yan, J.J.; et al. Bacteremia due to extended-spectrum-β-lactamase-producing Aeromonas spp. at a medical center in southern Taiwan. Antimicrob. Agents Chemother. 2011, 55, 5813–5818. [Google Scholar] [CrossRef] [PubMed]
- Agersø, Y.; Bruun, M.S.; Dalsgaard, I.; Larsen, J.L. The tetracycline resistance gene tet(E) is frequently occurring and present on large horizontally transferable plasmids in Aeromonas spp. from fish farms. Aquaculture 2007, 266, 47–52. [Google Scholar] [CrossRef]
- Kadlec, K.; von Czapiewski, E.; Kaspar, H.; Wallmann, J.; Michael, G.B.; Steinacker, U.; Schwarz, S. Molecular basis of sulfonamide and trimethoprim resistance in fish-pathogenic Aeromonas isolates. Appl. Env. Microbiol. 2011, 77, 7147–7150. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Shih, D.Y.C.; Wang, J.Y.; Yang, S.S. Molecular characterization of class 1 integrons and antimicrobial resistance in Aeromonas strains from foodborne outbreak-suspect samples and environmental sources in Taiwan. Diagn. Microbiol. Infect. Dis. 2007, 59, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, U.D.; Bhattacharjee, S. Bacteriological, clinical and virulence aspects of Aeromonas-associated diseases in humans. Pol. J. Microbiol. 2018, 67, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Antunes, P.; Campos, J.; Mourão, J.; Pereira, J.; Novais, C.; Peixe, L. Inflow water is a major source of trout farming contamination with Salmonella and multidrug resistant bacteria. Sci. Total Env. 2018, 642, 1163–1171. [Google Scholar] [CrossRef]
- Wen, Y.; Pu, X.; Zheng, W.; Hu, G. High prevalence of plasmid-mediated quinolone resistance and IncQ plasmids carrying qnrS2 gene in bacteria from rivers near hospitals and aquaculture in China. PLoS ONE 2016, 11, e0159418. [Google Scholar] [CrossRef]
- Jacoby, G.A.; Strahilevitz, J.; Hooper, D.C. Plasmid-Mediated Quinolone Resistance. Microbiol. Spectr. 2014. [Google Scholar] [CrossRef]
- Ishida, Y.; Ahmed, A.M.; Mahfouz, N.B.; Kimura, T.; El-Khodery, S.A.; Moawad, A.A.; Shimamoto, T. Molecular Analysis of Antimicrobial Resistance in Gram-Negative Bacteria Isolated from Fish Farms in Egypt. J. Vet. Med. Sci. 2010, 72, 727–734. [Google Scholar] [CrossRef] [Green Version]
- Aravena-Román, M.; Inglis, T.J.J.; Henderson, B.; Riley, T.V.; Chang, B.J. Antimicrobial susceptibilities of Aeromonas strains isolated from clinical and environmental sources to 26 antimicrobial agents. Antimicrob. Agents Chemother. 2012, 56, 1110–1112. [Google Scholar] [CrossRef]
- Lamy, B.; Kodjo, A.; Laurent, F.; Laurent, F. Prospective Nationwide Study of Aeromonas Infections in France. J. Clin. Microbiol. 2009, 47, 1234–1237. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Xu, C.; Sun, Q.; Schwarz, S.; Ou, Y.; Yang, L.; Huang, Z.; Eichhorn, I.; Walsh, T.R.; Wang, Y.; et al. Prevalence and Genetic Analysis of mcr-3 -Positive Aeromonas Species from Humans, Retail Meat, and Environmental Water Samples. Antimicrob. Agents Chemother. 2018. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bello-López, J.M.; Cabrero-Martínez, O.A.; Ibáñez-Cervantes, G.; Hernández-Cortez, C.; Pelcastre-Rodríguez, L.I.; Gonzalez-Avila, L.U.; Castro-Escarpulli, G. Horizontal Gene Transfer and Its Association with Antibiotic Resistance in the Genus Aeromonas spp. Microorganisms 2019, 7, 363. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090363
Bello-López JM, Cabrero-Martínez OA, Ibáñez-Cervantes G, Hernández-Cortez C, Pelcastre-Rodríguez LI, Gonzalez-Avila LU, Castro-Escarpulli G. Horizontal Gene Transfer and Its Association with Antibiotic Resistance in the Genus Aeromonas spp. Microorganisms. 2019; 7(9):363. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090363
Chicago/Turabian StyleBello-López, J. Manuel, Omar A. Cabrero-Martínez, Gabriela Ibáñez-Cervantes, Cecilia Hernández-Cortez, Leda I. Pelcastre-Rodríguez, Luis U. Gonzalez-Avila, and Graciela Castro-Escarpulli. 2019. "Horizontal Gene Transfer and Its Association with Antibiotic Resistance in the Genus Aeromonas spp." Microorganisms 7, no. 9: 363. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090363