Development of Salmonellosis as Affected by Bioactive Food Compounds
by
Ajay Kumar
1,*,
Abimbola Allison
2, 
Monica Henry
2,
Anita Scales
2 and
Aliyar Cyrus Fouladkhah
2,3,* 1
Division of Gastroenterology, Hepatology and Nutrition, Department of Pediatrics, University of Virginia School of Medicine, Charlottesville, VA 22908, USA
2
Public Health Microbiology Laboratory, Tennessee State University, Nashville, TN 37209, USA
3
Cooperative Extension Program, Tennessee State University, Nashville, TN 37209, USA
*
Authors to whom correspondence should be addressed.
Microorganisms 2019, 7(9), 364; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090364
Submission received: 30 August 2019
/
Revised: 15 September 2019
/
Accepted: 17 September 2019
/
Published: 18 September 2019
(This article belongs to the Special Issue Advances in Prevention of Foodborne Pathogens of Public Health Concern during Manufacturing)
Abstract
:Infections caused by Salmonella serovars are the leading cause of foodborne hospitalizations and deaths in Americans, extensively prevalent worldwide, and pose a considerable financial burden on public health infrastructure and private manufacturing. While a comprehensive review is lacking for delineating the role of dietary components on prevention of Salmonellosis, evidence for the role of diet for preventing the infection and management of Salmonellosis symptoms is increasing. The current study is an evaluation of preclinical and clinical studies and their underlying mechanisms to elaborate the efficacy of bioactive dietary components for augmenting the prevention of Salmonella infection. Studies investigating dietary components such as fibers, fatty acids, amino acids, vitamins, minerals, phenolic compounds, and probiotics exhibited efficacy of dietary compounds against Salmonellosis through manipulation of host bile acids, mucin, epithelial barrier, innate and adaptive immunity and gut microbiota as well as impacting the cellular signaling cascades of the pathogen. Pre-clinical studies investigating synergism and/or antagonistic activities of various bioactive compounds, additional randomized clinical trials, if not curtailed by lack of equipoise and ethical concerns, and well-planned epidemiological studies could augment the development of a validated and evidence-based guideline for mitigating the public health burden of human Salmonellosis through dietary compounds.
1. Introduction
Despite increased awareness and development of treatments such as antimicrobial interventions in manufacturing and antibiotic therapies in healthcare facilities for over a hundred years, Salmonella serovars are still a major concern in infectious diseases related premature morbidity and mortality [1,2,3,4]. Various serovars of Salmonella are the leading cause of foodborne hospitalizations and deaths in Americans causing over one million, about 20,000, and 378 annual illness, hospitalization, and deaths episodes, respectively [5]. Non-typhoidal Salmonella serovars are also the leading agent among most common foodborne infectious diseases, responsible for highest number (32,900 years) of disability adjusted life year (DALY), annually [6]. National Antimicrobial Resistance Monitoring System (NARMS) and other epidemiological sampling also reveal a widespread presence of multiple drug resistance (MDR) phenotypes of the pathogen in various facilities—as an example, 0.6% of ground meat samples may harbor MDR Salmonella [7] with approximately 7% of them displaying MDR-AmpC phenotype [8]. As such, the U.S. Department of Health and Human Services had categorized non-typhoidal Salmonella as a “serious threat” to the public health [9].
Data from world population also indicate that the pathogen is one of the leading causes of deaths associated with diarrheal diseases globally, with estimated 3.4 million cases (invasive non-typhoidal Salmonella serovars) and over 600,000 deaths annually [10,11]. The bacterium is a Gram-negative organism with a complicated and evolving nomenclature, currently consist of two species, at least six sub-species, and over 2500 serovars [12].
Changes in production and manufacturing practices, increased international commerce and travel, increased proportion of at-risk populations for infectious diseases, and changes in population’s eating habits during last few decades had contributed to increased incidences of Salmonella infections [13,14]. Salmonella serovars induce acute inflammation in the intestinal track after infection and utilizes the environment to further proliferate and colonize [15,16,17]. Colonization resistance against Salmonella is modulated by gut microflora, intestinal immunity, epithelium, and quality and quantity of digestive fluids. Various food components have been shown to modulate these factors and could be a potential intervention for reducing the likelihood of enteric infections.
Over the past 20 years, role of the dietary agents in shaping immunity against enteric infections has becoming increasingly evident [18,19,20,21,22] and piqued the interest in nutritional interventions for enteric infections. Several dietary components ranging from polyphenolic compounds, fibers, micronutrients, fatty acids, peptides, and carbohydrates of plant and animal origin had been shown efficacious against Salmonella serovars in various experimental models [22,23,24,25,26,27,28,29,30,31,32]. These associations are the result of an array of potential biochemical pathways, very complex and dynamic in nature, including interactions among dietary components, gut epithelium, digestive system, immune system and gut microbiota as affected by various seasons [33,34,35,36,37]. Better understanding of these underlying mechanisms could reduce Salmonella prevalence in the food chain though modifications in food animal diets. It could further reduce the public health burden of non-typhoidal Salmonella serovars by mitigating severe symptoms and reducing the pathogen DALY and mortality rate in healthcare facilities for Salmonellosis patients. Hence, the current work is a review of Salmonella infection studies as affected by various dietary components with discussions of the mechanisms of action and types of preclinical, animal models, and clinical studies employed.
2. Current Status of Knowledge
2.1. Effect of Dietary Components against Salmonella: In-Vitro Models
Dietary components may prevent infection outcome by directly affecting the pathogen multiplication and virulence [7,38,39] or by modulating host response to the pathogens [26,37]. To test the direct effects of dietary components on pathogens, researchers have used food extracts or dietary bioactive components on Salmonella cultures. Summary of potential relationships between dietary components and Salmonella infections are presented in Figure 1. Following treatment of dietary components, multiplication, and gene expression for virulence and motility could be measured. These models are comparatively less expensive and less cumbersome to assess the efficacy of dietary components for Salmonella infection. For instance, several essential oils were added to Salmonella growth media at various doses and Salmonella multiplication was compared with untreated controls [38]. Among 28 tested essential oils, Origanum heracleoticum, Cinnamomum cassia, Corydothymus capitatus, Satureja montana, and Cinnamomum verum were particularly effective against Salmonella Typhimurium [38]. Citrus flavonoids were similarly evaluated on Salmonella virulence gene expression [39]. In the study, Naringenin, a flavanone present in grapefruit, repressed 24 genes in pathogenicity island of Salmonella Typhimurium LT2 and further down-regulated 17 genes associated with the pathogen motility [39]. Most recent studies also reveal similar trends, for example, various essential oils extracted from Aloysia triphylla, Cinnamomum zeylanicum, Cymbopogon citratus, Litsea cubeba, Mentha piperita, and Syzygium aromaticum had been shown to be efficacious against Salmonella serovars during in vitro challenge studies [40].
It is noteworthy that aforementioned studies are conducted without host interaction and interpretation and generalization of the results should be drawn with caution and after further investigations in presence of host cells.
Orally infected Salmonella can enter circulation through various routes. It can invade several phagocytic and non-phagocytic cells depending upon serotype. In the murine model, Salmonella invades both phagocytic and epithelial non-phagocytic cell types. Hence, in vitro models of Salmonella entry have been developed to assess the effect of a test compound on a host. The Salmonella entry model could reveal the mechanism of action of a test compound on an organism. Several human and mouse cell lines such as Caco-2 [41] and RAW264.7 have been used in the literature to test efficacy of the compounds against Salmonella entry. For example, secretory immunoglobulin A (SIgA) was demonstrated to be a potent inhibitor for Salmonella Typhimurium entry into polarized monolayers of HeLa cells [42].
Salmonella contains the pathogenicity islands for the secretion of effector molecules to infect the target cells [43]. The molecules released by these secretory systems change the host cell cytoskeleton to facilitate Salmonella entry. The in vitro Salmonella entry models are impactful in studying the effects of dietary components on Salmonella as well as on host cells. However, these studies do not represent involvement of all host cell types that are simultaneously present in gastrointestinal area of humans. Dietary components could affect Salmonella virulence by affecting secretory systems or by competing with Salmonella for the receptors on host cells [39]. Host cells can also release the cytokines in response to the dietary components that can affect Salmonella virulence or motility. Therefore, in vitro models of Salmonella infection can have great implications for assessing mechanisms of actions by the dietary components.
2.2. Summary of Effect of Dietary Components on Salmonella Infection in Rodent Models
The fecal shedding of Salmonella, tissue colonization, local and systemic inflammatory changes, survival and weight reduction are the major observable changes associated with Salmonella infections in rodents. Bovee-Oudenhoven et al. showed reduced Salmonella fecal shedding when fructooligosaccharides were fed to the male Wistar rats as compared to the cellulose-fed group after 2 weeks of dietary intervention [44]. Furthermore, dietary fructooligosaccharides increased fecal Lactobacilli count and increased the translocation of Salmonella to the liver and spleen with an increase in fecal mucin as compared to cellulose fed rats [19]. The author concluded that dietary fructooligosaccharides decreased Salmonella colonization but increased the translocation potentially due to irritation of mucosal membrane. Some of the mice strains succumb easily to Salmonella infection and hence survival rate is the primary indicator of the dietary efficacy against infection. Hitchins et al. showed that feeding of freeze dried yoghurt to male weanling Sprague-Dawley rats increased overall survival rate and weight of the animals after intraperitoneal Salmonella challenge as compared to rats fed on milk diet for 1 week [45]. Similarly, dietary feeding of Herba Pogostemonis extract to Balb/c mice increased the overall survival rate as compared to control diet fed animals after intraperitoneal Salmonella challenge [46]. Feeding of Herba Pogostemonis (Pogostemon cablin Bantham extract) also reduced Salmonella liver damage as compared to control diet fed animals [46]. Recent studies similarly show association among various bioactive food compounds and prevention of Salmonellosis. Supplementing the diet of albino rats with olive oil, as an example, had been shown to have efficacy against Salmonella Typhi as a natural antimicrobial and non-toxic immune modulator [47]. These studies show that there are measurable markers for Salmonella infections in rodents and they can be used as a model to mimic Salmonella infections in human host.
Both foodborne pathogens and dietary components pass through the stomach acid, when ingested. Hence, gastric acidity is one of the important factors in determining stability of enteric pathogens. In a randomized controlled clinical trial, gastric hypochlorhydria (low hydrochloric acid) was found to be associated with increased Salmonella infections [48]. This hypothesis was also confirmed in the rodent model of Salmonella infection. Tennant et al. [49] showed that treatment of mice with antacids resulted in the decreased infectious dose of Salmonella as compared to normal mice.
Similar results were also observed in a constitutively hypochlorhydric mice (proton pump mutation) as compared to the normal mice [49]. Additionally, gastric pH not only affects the survival of pathogens but also affects digestion and absorption of foods. Lucas et al. showed that an increase in pH from 1.5 to 2.5 reduced digestion of the kiwifruit peptides [50]. Gastric pH also modulates absorption of micronutrients such as zinc. Henderson et al. observed higher plasma zinc levels in the young healthy volunteers at low pH as compared to plasma level in higher gastric pH volunteers [51]. The gastric pH is considerably different across species. For instance, mean gastric pH in mice is 3.1–4.5 and in rats ranges from 3.2 to 3.9, whereas in the humans it is 1.5–3.5. In addition to gastric pH, intestinal pH is also different in rodents as compared to humans. Mice and rats have a mean intestinal pH of 5.2 and 6.6, respectively, as compared to 7.2 in humans [52]. These studies show that gastric and intestinal pH could potentially affect bioactivity of dietary components and should be considered as one of the important factors in selecting a study model.
In rodents and humans, several disease symptoms can be confounding due to the differences in their anatomy and physiology. For example, in the non-typhoidal salmonellosis, vomiting and diarrhea are the main symptoms in humans. However, anatomically mice cannot vomit and due to this reason, the assessment of diarrhea could be very difficult in mice. In these cases, it becomes harder to translate the finding into clinical applications. Hence, these limitations of rodent models should be taken into consideration while interpreting the results from the dietary intervention studies for Salmonella infections in the rodent models for application in human clinical trials.
2.3. Summary of Effect of Dietary Components on Salmonella Infection in Pig Models
Pigs have been used in several studies involving dietary interventions [53]. Pigs have many more similarities to the human gastrointestinal tract as compared to rodents. Humans and pigs are similar in the body composition, cardiovascular, renal, nutritional, immunological, metabolic, and gastrointestinal aspects [53]. As such, several studies have been conducted in pig models of Salmonella infection interactions with dietary interventions. Michiels et al. demonstrated that supplementation of a mixture of formic, sorbic, and benzoic acid to the piglets for 35 days, significantly reduced the Salmonella fecal shedding as compared to the control group after oral challenge [54]. Dietary organic acids increase fecal cytotoxicity to Salmonella, but the effect can be dependent upon the environmental temperature. Rajtak et al. exhibited that supplementation of a pig diet with organic acid (Potassium- diformate) reduced the survival of Salmonella in pig feces when incubated at 22 °C but not at 4 °C [55]. Boyen et al. fed the supplemented diet with the coated butyric acid (2 g/kg of diet) to the pigs for 12 days and orally challenged the animals with Salmonella [56]. Fecal shedding of Salmonella was decreased in the coated butyric acids fed animals as compared to the un-coated group. It was hypothesized that coating prevents the degradation of fatty acids in the intestinal tract [56]. Dietary supplements also reduced inflammation after Salmonella infection in pigs. Chen et al. supplemented the pig diet with arginine (0.5%) for 1 week and infected the pigs intramuscularly with Salmonella [57]. Effects of various essential oils have been similarly reviewed by Omonijo et al. as effective antimicrobials in Swine production [58].
Fecal Salmonella shedding is one of the distinctive biomarkers of Salmonella infection in pig models. However, Salmonella colonization patterns are different in pigs as compared to humans. For instance, Salmonella Typhimurium has been observed to colonize in tonsils and respiratory tissues of infected pigs [59], whereas in humans, it does not colonize at those sites. The pig stomach is 2–3 times larger compared to humans [52], this anatomical difference may have impacts on Salmonella survival and digestibility of dietary components. Pig cecum is also several folds larger than the human cecum and may have implications in the Salmonella colonization [52]. In humans, stomach pH before eating is around 5, however, in pigs it is below 2. Consequently, pigs release a much greater extent of bile in the duodenum as compared to humans. Due to antimicrobial activities, bile could impact colonization of Salmonella in the proximal small intestine. Additionally, it can modulate digestion and absorption of the dietary components. Besides these differences, pigs are different in gastrointestinal thickness of mucus, and gastrointestinal motility and transit, as compared to humans. The distal small intestine of pigs contains a larger number of microbes as compared to humans and can degrade some carbohydrates with low digestibility compared to humans [60]. Hence, similar dietary interventions in pigs and humans may exhibit different potential. The pig immune system also differs from humans, however, implications of this difference have not been studied in regard to enteric infections. For instance, the gut of neonate piglets completely lacks leukocytes whereas human infants have a few leukocytes at birth [61]. Pig intestine contains a larger number of Peyer’s patches as compared to humans throughout the intestine [61].
2.4. Summary of Effect of Dietary Components on Salmonella Infection in Calf Models
Although there are appreciable differences between monogasters and ruminants, calves develop very similar clinical and pathological features such as diarrhea and enteritis to human, hence, calves are considered one of most reliable models to mimic the human non-typhoidal salmonellosis [36]. These similarities have been also discussed by Higginson et al. [62].
After Salmonella infection, the calves show similar clinical symptoms as humans such as fever, diarrhea, anorexia and dehydration and the intestinal pathological changes [63]. Hill et al. revealed that feeding of a commercially available blend of butyric acid, coconut oil, and flax oil to the male Holstein calves for 28 days altered the inflammatory response to intraperitoneal Salmonella toxoid as compared to the control group [64]. The dietary blend reduced hyperthermia, hypophagia, and serum TNF-α but increased the IL-4 as compared to the control group [64].
Despite above-mentioned similarities, calves also exhibit significant anatomical and physiological differences in the digestive system relative to humans. A ruminant’s stomach is four chambered and contains a large number of microflora that digests fibers, especially cellulose which remain undigested in humans. Sugars are fermented in ruminant stomach and as a result several volatile fatty acids are produced [65]. Most of the carbohydrates are converted into volatile fatty acids and a very small proportion of carbohydrates are absorbed as glucose. Additionally, the ruminant microflora differs from the human gut microflora to a great extent [66]. Hence, the same dietary components may produce different metabolites and physiological effects as compared to humans. Logistically, calves need a large amount of food and it is very expensive to conduct the dietary experimental studies in this model.
2.5. Summary of Dietary Interventions for Salmonella Infection in Humans
A variety of Salmonella serovars infect humans. Epidemiological studies have shown that typhoidal and non-typhoidal salmonellosis are the predominant types of infections [67]. Salmonellosis is clinically prognosed by headache, diarrhea, constipation, abdominal pain, chills, loss of appetite, and fever with an incubation time varying from hours to several days [68,69]. Typhoidal salmonellosis is less prevalent in the United States as compared to other developing countries [67]. In contrast, non-typhoidal salmonellosis presents a major and persisting public health challenge in North America. From 1998 to 2017, over 2600 single or multi-state non-typhoidal Salmonella outbreaks have occurred in the United States associated with animal and plant based foods [70]. Symptoms could be self-limiting, lasting for 1 week without treatment but could also lead to serious complications if left untreated, especially in immunocompromised subjects and those in at-risk populations [69]. Antimicrobial therapy is the first choice of treatment in persistent human salmonellosis. However, as discussed in the introduction section, the problem of drug resistance has become more prevalent due to extensive therapeutic use of antibiotics in healthcare facilities and subtherapeutic doses during animal food production [71]. Hence, dietary prophylactic interventions could be further utilized for prevention and alleviating symptoms of Salmonella infections. A few dietary prophylactic studies have been conducted in children for prevention of Salmonella infections. Stool frequency, vomiting, and Salmonella fecal shedding are the parameters measured in these clinical trials. Several other disease conditions also affect the incidence of Salmonella infections. Di Cagno et al. revealed that administration of gluten free diets in children with celiac disease did not reduce Salmonella shedding from stool as compared to healthy children [72]. Other dietary interventions are effective in reducing Salmonella infection. Lara et al. showed that feeding of dairy products containing probiotic mixtures of various strains of Lactobacillus to healthy children for 6 weeks decreased Salmonella serovars adhesion to the intestinal mucin [73]. Dietary interventions can also reduce frequency of stool and vomiting in Salmonella infected children. Rabbani et al. revealed that feeding of cooked banana for 1 week in children having persistent diarrhea, reduced frequency of the stool and vomiting as compared to children fed only with rice diet [74]. In another clinical trial in children, fermented food (lactic-acid fermented cereal gruel) was fed to healthy children three times a day for 2 weeks. After 2 weeks of feeding, stool swabs were taken from the treated and non-treated groups and analyzed for the presence of enteropathogenic bacteria including Salmonella. The fermented food reduced the presence of enteropathogenic bacteria as compared to the control diet [75]. These studies show that dietary interventions can be effective in the management of diarrheal diseases. However, there are several constraints in conducting dietary studies in Salmonella infections in humans that are prophylactic in nature. In addition to clinical equipoise, the major issues in conducting human clinical trials are time, cost, availability of appropriate stool and serum biomarkers and overall patient compliance and ethics. In presence of these curtailments, a dietary intervention could be pre-clinically evaluated in a relevant animal model to predict the safety and efficacy of the compound prior to administration in clinical trials [76].
It is noteworthy that bioactive compounds and probiotic diet might have a positive effect on colonization of Salmonella serovars in gastrointestinal area. As an example, a probiotic diet containing Enterococcus spp. could lead to increased fecal excreting and colonization of Salmonella in organs of piglets [77]. The current study is limited to discussing the literature that demonstrates antagonistic efficacy against colonization of Salmonella serovars, rather than those enhancing proliferation of the pathogen. Table 1 summarizes the pros and cons of Salmonella models discussed in the current study.
3. Potential Mechanisms of Protection against Salmonella Infections
3.1. Alteration in Bile Quality and Quantity
Bile is an important digestive fluid synthesized by the liver of many vertebrates. Bile plays a role in digestion of fats in small intestine by emulsification, micelle formation. As a result, absorption of fat-soluble vitamins such as vitamin A, D, E, and K is also increased in the presence of bile. Bile is stored in the gall bladder and released into the duodenum after receiving stimuli in the form of semi-digested fats and proteins from stomach. After digestion of fats, the majority of the bile is reabsorbed in terminal ileum. Cholycystokinin and secritin hormones in the gut control this process. Bile is alkaline and composed of phospholipids, bile acids, and surfactants. In the duodenum alkaline pH neutralizes stomach acid [78].
In addition to digestive role of bile, it exhibits an antimicrobial role against gastrointestinal pathogens [79]. Both bile quality and bile quantity may determine the multiplication of enteric pathogen [80]. Bile salts have been shown to act as antimicrobials especially on Salmonella and other enteric infections [81,82]. Different dietary fibers have been shown to affect bile composition to different extent and to improve colonization resistance against enteric pathogens [83]. Inagaki et al. showed that bile acids induces genes involved in enteroprotection by inhibiting pathogenic overgrowth and mucosal injury in the ileum in a mouse model of infection [84]. Diet consists of several compounds of plant and animal origin and hence considered as a multi-targeting intervention for prevention of enteric infection. Xu et al. [85] showed that consumption of dietary medium chain fatty acids increased fecal bile acids (cholic acid) significantly as compared to control group in C57BL/6J Mice. Kollanoor et al. demonstrated that feeding of Caprylic acid (a medium chain fatty acid) to poultry significantly reduced Salmonella infection in the intestine as well in organs therapeutically [83]. Further, in vitro study in hepatocytes showed that addition of medium chain fatty acids in culture media enhances cell surface expression and transport capacity of bile salt export pump (BSEP/ABCB11) [86]. Costarelli et al. compared diets containing different fatty acids in healthy premenopausal women and found that dietary linoleate increased postparandial plasma bile acid and cholycytokinin as compared to low fat diet [87]. Dietary fish oil increased fecal bile acids in a rodent model without increased gene expression for bile synthesis in the liver [88]. This study suggests although not all fatty acids increase bile acid synthesis in the liver, some could reduce bile absorption in the ileum. Studies have further exhibited that the change in bile acid release alters pH of the intestine and affects Salmonella adherence and survival. Several Salmonella genes are affected in the presence or absence of bile. Both bile quality and quantity have been shown repress Salmonella virulence in gut environment in in vivo models [89,90,91]. Antunes et al. [92] showed that Salmonella could multiply in the gall bladder of susceptible mice and causes typhoid. Bile acids exert antimicrobial actions on pathogens by virtue of their detergent properties. Cholic and deoxycholic acids in bile can damage bacterial DNA [79].
Dietary factors such as fiber may bind to bile acids and reduce reabsorption in colon [93]. Oat bran, pectin, and guar gum have been shown to increase bile acids in fecal matter [94,95,96]. Reduction in reabsorption of bile acids in the large intestine modulates the gut hormone feedback system and stimulates the liver to synthesize more bile acids [78]. This process could reduce alkalinity of the small intestine, and may increase gut motility, making the gut environment unfit for Salmonella infection [97].
3.2. Gut Mucosa
In order to reach epithelium, Salmonella needs to cross luminal barriers. Intestinal mucous is the first line of defense to Salmonella in the small intestine of rodents and humans [98]. Mucus in the small intestine is single layered and loosely attached to epithelium as compared to double-layered mucus of colon. Mucous is made up of secretory proteins called mucins and the predominant mucin in small intestine is Muc2 [99]. Abnormalities in mucous layers, underproduction of Muc2 by goblet cells and mutated Muc2 results in elevated risk for bacterial infection [100]. A study shows that during Salmonella infection, the mucin layer is disrupted and Salmonella obtain access to epithelium [101].
Various components of diet have been shown to upregulate expression of Muc2 in intestinal cells. Willemsen et al. showed that treatment of intestinal epithelial and fibroblast co-culture with short chain fatty acids significantly increased expression of Muc2 [102]. Ingestion of dietary fibers (soluble and insoluble) has been shown to increases proliferation of goblet cells and sialylated mucin in the small intestine of rats [103]. In another study, feeding of inulin/fructans in a rodent trial significantly increases mucous layer thickness in the colon and increases the number of goblet cells in crypts of distal jejunum as compared to control diet [104]. Morita et al. similarly exhibited that intake of dietary resistant starch in rodents reduces endotoxin influx from intestinal tissue and hypothesized that it could be partially due to alterations in mucosal barrier functions [105].
3.3. Antimicrobial Activities
After crossing the mucin layer in the gut, enteric pathogens need to penetrate epithelial layer in order to infect the organism. Human gut epithelia consist of a monolayer of epithelial cells. It separates the gut lumen from the lamina propria. Intestinal epithelial cellular junctions affect intestinal permeability as well as transcytosis capacity of individual cells. Strong cellular junctions are necessary to avoid the invasion of pathogens through epithelium. Salmonella can breach the epithelial barrier by employing para-cellular and trans-cellular mechanisms, including actin cytoskeleton of the epithelial cells and the secretion of the effector molecules [30].
Dietary components have been discussed in the past as factors to modulate the epithelial barrier [106]. Diet can have both positive and negative impacts on epithelial integrity. Liu et al. showed that when a high grain diet was fed to male goats, it resulted in the disruption of the ruminal epithelium as measured by the presence of systemic lipopolysacharide (LPS) [107]. However, diet can also impact epithelial integrity positively. In a study by Nofrarias et al., pigs were fed resistant starch for 97 days and consequently increased hypertrophy, reduced apoptosis in the crypts, lymphoid nodules in the colon, and increased mucin sulfuration were observed. These changes promoted epithelial protection compared to the control dietary group containing digestible starch [108]. Dietary components can also modulate the epithelial proteins such as occludins that secure junctions between the adjacent cells in the gut epithelium. Enteric pathogenic bacteria secrete LPS that causes inflammation and escalates loss of protein occludin that decreases the barrier function of epithelium. Park et al. showed in a rodent trial that dietary administration of gangliosides (a lipid) prevents LPS induced degradation of the occludin and reduces the total nitric oxide in the gut mucosa [109]. An in vitro study with Caco-2 cells demonstrated that addition of quercetin (a flavonoid) induces expression of zonula occludens-2, occludin, and claudin-1 and claudin-4 as compared to the control group [110]. All of these proteins play an important part in maintaining epithelial integrity. Salmonella entry into epithelial cells can result in epithelial necrosis and apoptosis. Int-407 cell line (human intestinal cell line) showed a significantly lesser extent of necrosis and apoptosis during Salmonella infection when treated with sterols and fatty acids found in the root extract of Hemidusmus indicus as compared to an untreated cell line [111]. Hence, protection of the epithelium can be considered an important target of dietary interventions in Salmonella infections.
3.4. Gut Microbiome
The gut contains more than a trillion symbiotic bacteria that play a major role in developing immunity as well as resistance against enteric infections. Initially it was hypothesized that the genetic factors were responsible for susceptible and resistant mouse strains against the enteric infections. However, currently literature delineates that the genetic factors are only one of the determinants of composition and structure of the gut microflora. As an example, Willing et al. successfully transferred the microbiota from resistant to susceptible mice and observed a delayed colonization of Citrobacter rodentium and mortality in susceptible strain [112]. In the same study, native gut microbiota of resistant mice was depleted by oral streptomycin (20 mg) 24 h prior to transplantation and replaced by the microbiota from susceptible mice. As a result, the oral antibiotic treatment reduced the innate defenses and a severe infection pathology was observed as compared to mice in control group. This experiment demonstrates that gut microbiota plays an important role in fighting the infection [112]. Similarly, mice were given a combination of antibiotics (Streptomycin, Vancomycin, Ampicillin, Neomycin, and Metronidazole) for 1 week in drinking water and later orally challenged with Salmonella Typhimurium 14028. The mice on the antibiotics showed a significantly higher number of Salmonella DNA in the cecum and large intestine as compared to control mice group [113]. The gut microbiota may affect enteric infections by modulating the intestinal immunity or by the direct competition. Symbiotic gut microbiota competes with pathogens for the nutrients such as iron and carbon sources [114]. Stelter et al. showed that Salmonella-induced mucosal lactins kills symbiotic gut microflora and then Salmonella takes advantage of this process for survival in gastrointestinal tract [115]. Salmonella induces acute inflammation in mice and neutrophils are recruited at the site of infection. Gill et al. showed that neutrophil elastases can shift mice gut microbiota and increase Salmonella colonization, while neutralization of neutrophil elastases decrease colonization of Salmonella [116]. These studies show that gut microbiota play an important role in protection from Salmonella infections and modulation of gut microflora for prevention of enteric infections warrants further studies.
Given the role of gut microbiota in protection against Salmonella, several studies have been conducted to test effects of dairy and native gut probiotics on Salmonella colonization. Probiotics are the microorganisms that induce health benefits when consumed in effective doses. Lactobacillus and Streptococcus are two widely studied categories of probiotics and their effectiveness against Salmonella is articulated by Castillo et al. [117]. Lactobacillus rhamnosus has been shown to reduce Salmonella adhesion to epithelial cells in in vitro model of Salmonella infection [118]. Probiotics not only compete with Salmonella for nutrients but also enhance protective immunity against the pathogen. Castillo et al. showed that oral administration of Lactobacillus in mice changes cytokine production and Toll Like Receptor (TLR) expression that is protective for mice against Salmonella infection [119]. Moreover, probiotics such as Bifidobacterium can directly affect virulence of Salmonella by releasing the molecules that down-regulate the expression of pathogenicity islands 1 and 2 [120]. Hence, Lactobacillus and Bifidobacterium have emerged as potential contributors for protection against enteric infections such as Salmonella serovars.
Diet is a major factor in the establishment of gut microbiome. As an example, previous studies exhibit that a change of diet from low-fat, high plant-based polysaccharide to the high-fat, and high simple sugar diet, could change structure of the gut microbiota very rapidly [121]. A shift of low-fat diet to the Western diet also changes metabolic pathways and modulates gene expression in gut microbiome [122]. Humanized mice (mice transplanted with human gut microflora) when fed a Western-type diet, showed an increased adiposity and this trait was transmissible through the transplantation of the gut microbiota in other mice [122]. Diet could also modulate gut microbiota directly by providing prebiotics—many studies have exhibited the efficacy of the prebiotics such as dietary fiber, fatty acids, and polyphenols for a shift in gut microflora [123,124].
3.5. Gut Immunity
The immune system of the gastrointestinal tract is the largest segment of the mammalian immune system. The gut encounters massive amounts of pathogens and dietary antigens that need to be neutralized. These functions emphasize the importance of gut immune system. The mucosal immune system is equipped with innate and adaptive immune defense mechanisms. Innate immunity provides the first line of defense against pathogens. The major players of the innate immune defense are macrophages, monocytes, neutrophils, epithelial cells, natural killer (NK) cells, and dendritic cells (DCs) [125]. Dendritic cells, macrophages, and epithelial cells are also termed as antigen presenting cells (APCs) because of their capacity of processing and presenting foreign antigens to other cells. APCs have a series of receptors called Pattern Recognition Receptors (PRRs) on their surfaces such as TLRs and Nod Like Receptors (NODs) to recognize the pathogens [126]. These receptors recognize motifs on pathogens known as the Pathogen Associated Molecular Patterns (PAMPs) [127]. The innate immune cells release inflammatory cytokines and mediators after sensing the PAMPs [128]. However, if innate immunity fails to resolve the inflammation and eliminate pathogen, adaptive immunity enters this process. In the gut adaptive immune system, the predominant response is antibody mediated and is represented by the Immunoglobulin A (IgA) [129]. The IgA is chiefly produced by the B cells in the intestinal mucosa triggered by anti-inflammatory cytokines such as TGF-β and IL-10 [130]. Hence, both innate and adaptive immune responses are required in the protection against infection and depends upon type of pathogen.
The role of the gut immune system in protection from enteric infections has been studied intensely [131,132,133]. Primary Salmonella infection increases interferon gamma (IFN-γ), tumor necrosis factor alpha (TNF-α), and interleukin 12 (IL-12) in circulation and in local tissues [133,134,135]. Major sources of IFN-γ and TNF-α are neutrophils and macrophages [136]. IL-12 is a cytokine induced in response to several bacteria and mediates onset of the Th1 protective response. Natural killer T (NKT) cells produce IFN-γ in response to IL-12 [137]. Infected macrophages also interact with NK cells in order to produce IFN-γ in humans [138]. Even though initial innate immune response restricts infection to a certain extent, it fails to inhibit multiplication of pathogens in deeper tissues. Hence, immune response is switched to adaptive response after some time and is achieved mainly by induction of CD4+ T cells, CD8+ T cells, and B cells [139]. In experimental models, depletion of CD4+ T cells had a more pronounced effect on protection from Salmonella as compared to CD8+ T cells. However, underlying mechanisms are not clear. The second major adaptive response to Salmonella is induction of the B cells to produce antibodies such as IgA. The antibodies bind Salmonella and prevent entry into deeper tissues. Administration of B cell hybridoma producing Salmonella specific IgA has been shown to prevent oral Salmonella infection in the mice [140]. These studies exhibited the potentially appreciable role of bioactive compounds for augmenting host immunity against Salmonella infections.
Dietary components such as dietary fiber and prebiotics manipulate both the innate and adaptive immunity [141]. Galdeano et al. demonstrated that feeding of probiotic fermented milk to the rats increases the number of macrophages and DCs with an increase in IFN-γ, TNF-α, and IL-12 after 5 days of nutrition [142]. Nutrients such as glutamine, arginine, vitamin A, and zinc have protective impacts against enteric infections [143]. Macrophages play an important role in clearance of Salmonella in primary infections. Modified arabinoxylan rice bran improves the phagocytic function of macrophages in the in vitro models of RAW264.7 cells [144]. Treatment of macrophages with the modified arabinoxylan rice bran increased the attachment and phagocytosis of yeast cells with an increase in TNF-α and IL-6 [144]. Wang et al. showed an enhanced Salmonella specific immune response in the orally vaccinated mice with attenuated Salmonella and fed with white button mushroom powder as compared to the only vaccinated mice [145]. The white button mushroom fed mice had higher number of Salmonella specific fecal IgA, IFN-γ, and TNF-α in splenocytes. These mice also showed an increased number of DCs and activation marker CD40 in splenocytes as compared to the control mice [145]. These studies show that dietary interventions could modulate pro-inflammatory responses and manipulate the innate and adaptive immunity [141].
4. Conclusions
Various dietary components could have considerable efficacy on prevention of Salmonella serovars infections. These effects may involve various mechanisms through impacting the gastrointestinal microbiota, immune system, and epithelium. The efficacy of various bioactive compounds for inhibiting the proliferation of Salmonella serovars in various in vitro, in vivo, animal models, and randomized studies reviewed creates the opportunity of mitigating the burden of Salmonellosis through dietary intervention. Despite striking similarities, animal models have major differences with human anatomy, as such delineated differences should be considered diligently for interpretation of these studies. Clinical equipoise, cost, time, and other ethical issues are also major curtailments for further conduct of randomized clinical trials with human subjects. The vast majority of the discussed literature demonstrate efficacy and mechanism of action of a sole bioactive compound. Pre-clinical studies investigating synergism and/or antagonistic activities of an array of bioactive compounds, additional randomized clinical trials, and well-planned epidemiological studies with comprehensive plans for control of confounders could augment the development of a validated and evidence-based guideline for mitigating the public health burden of human Salmonellosis through dietary compounds.
Author Contributions
A.K. co-wrote the first version of the manuscript. Authors A.A., M.H., and A.S. assisted in completion of the manuscript and reviewing the references for accuracy and formatting. A.F. co-wrote, revised, and edited the manuscript.
Acknowledgments
All authors read and approved the final manuscript. Author A.K. would like to express gratitude to former committee members of his doctoral degree for their support and encouragements. Financial support in part from the National Institute of Food and Agriculture of USDA (2017-07534; 2017-07975; 2017-06088) is acknowledged gratefully by the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Herrick, R.L.; Buchberger, S.G.; Clark, R.M.; Kupferle, M.; Murray, R.; Succop, P. A Markov model to estimate Salmonella morbidity, mortality, illness duration, and cost. Health Econ. 2012, 21, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Fouladkhah, A.; Geornaras, I.; Yang, H.; Belk, K.; Nightingale, K.K.; Woerner, D.; Smith, G.C.; Sofos, J.N. Sensitivity of Shiga Toxin-Producing Escherichia coli, Multidrug Resistant Salmonella, and Antibiotic Susceptible Salmonella to Lactic Acid on Inoculated Beef Trimmings. J. Food Prot. 2012, 75, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Fouladkhah, A.; Geornaras, I.; Sofos, J. Biofilm Formation of O157 and Non-O157 Shiga Toxin-Producing Escherichia coli and Multidrug-Resistant and Susceptible Salmonella Typhimurium and Newport and Their Inactivation by Sanitizers. J. Food Sci. 2013, 78, M880–M886. [Google Scholar] [CrossRef] [PubMed]
- Fouladkhah, A.; Geornaras, I.; Yang, H.; Sofos, J. Lactic Acid Resistance of Shiga Toxin-Producing Escherichia coli and Multidrug-Resistant and Susceptible Salmonella Typhimurium and Salmonella Newport in Meat Homogenate. Food Microbiol. 2013, 36, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States—major Pathogens. J. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Mahon, B.E.; Jones, T.F.; Griffin, P.M. An Assessment of the Human Health Impact of Seven Leading Foodborne Pathogens in the United States Using Disability Adjusted Life Years. Epidemiol. Infect. 2015, 143, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Bosilevac, J.M.; Arthur, T.M.; Bono, J.L.; Brichta-Harhay, D.M.; Kalchayanad, N.; King, D.A.; Shackelford, S.D.; Wheeler, M.L.; Koohmaraie, M. Prevalence and Enumeration of Escherichia coli O157: H7 and Salmonella in U.S. Abattoirs That Process Fewer than 1000 Head of Cattle per Day. J. Food Prot. 2009, 72, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Blickenstaff, K.; Glenn, A.; Ayers, S.L.; Friedman, S.L.; Abbott, J.W.; McDermott, P.F. Lactam Resistance in Salmonella Strains Isolated from Retail Meats in the United States by the National Antimicrobial Resistance Monitoring System between 2002 and 2006. Appl. Environ. Microbiol. 2009, 75, 7624–7630. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (US). Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention (US): Antlanta, GA, USA, 2013.
- Ao, T.T.; Feasey, N.A.; Gordon, M.A.; Keddy, K.H.; Angulo, F.J.; Crump, J.A. Global Burden of Invasive Nontyphoidal Salmonella Disease, 2010 (1). Emerg. Infect. Dis. 2015, 21, 941. [Google Scholar] [CrossRef]
- World Health Organization. Global Burden of Food Safety. Available online: Http://Www.Who.Int/Foodsafety/Areas_work/Foodborne-Diseases/Ferg/En/ (accessed on 1 January 2019).
- Grimont, P.A.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars. WHO Collab. Cent. Ref. Res. Salmonella 2007, 9, 1–166. [Google Scholar]
- Kearney, K. Food consumption trends and drivers. Philos. Trans. R. Soc. B 2010, 365, 2793–2807. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.L. Changing patterns of infectious disease. Nature 2000, 406, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Thiennimitr, P.; Winter, S.E.; Winter, M.G.; Xavier, M.N.; Tolstikov, V.; Huseby, D.L.; Sterzenbach, T.; Tsolis, R.M.; Roth, J.R.; Bäumler, A.J. Intestinal inflammation allows Salmonella to use ethanolamine to compete with the microbiota. Proc. Natl. Acad. Sci. USA 2011, 108, 17480–17485. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.E.; Thiennimitr, P.; Winter, M.G.; Butler, B.P.; Huseby, D.L.; Crawford, R.W.; Russell, J.M.; Bevins, C.L.; Adams, L.G.; Tsolis, R.M.; et al. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 2010, 467, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Stecher, B.; Robbiani, R.; Walker, A.W.; Westendorf, A.M.; Barthel, M.; Kremer, M.; Chaffron, S.; Macpherson, A.J.; Buer, J.; Parkhill, J.; et al. Salmonella enterica serovar typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol. 2007, 5, 2177–2189. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Kew, S. The immune system: A target for functional foods? Br. J. Nutr. 2002, 88, S165–S177. [Google Scholar] [CrossRef]
- Harrison, L.M.; Balan, K.V.; Babu, U.S. Dietary fatty acids and immune response to food-borne bacterial infections. Nutrients 2013, 5, 1801–1822. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.K.; Cao, W.; Vora, K.P.; Cruz, J.D.L.; Shieh, W.J.; Zaki, S.R.; Katz, J.M.; Sambhara, S.; Gangappa, S. Protein energy malnutrition decreases immunity and increases susceptibility to influenza infection in mice. J. Infect. Dis. 2012, 207, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Hekmatdoost, A.; Wu, X.; Morampudi, V.; Innis, S.M.; Jacobson, K. Dietary oils modify the host immune response and colonic tissue damage following Citrobacter rodentium infection in mice. Am. J. Physiol. 2013, 304, G917–G928. [Google Scholar] [CrossRef]
- Chandra, R.K. Nutrition, immunity and infection: From basic knowledge of dietary manipulation of immune responses to practical application of ameliorating suffering and improving survival. Proc. Natl. Acad. Sci. USA 1996, 93, 14304–14307. [Google Scholar] [CrossRef] [Green Version]
- He, R.R.; Wang, M.; Wang, C.Z.; Chen, B.T.; Lu, C.N.; Yao, X.S.; Chen, J.X.; Kurihara, H. Protective effect of apple polyphenols against stress-provoked influenza viral infection in restraint mice. J. Agric. Food Chem. 2011, 59, 3730–3737. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.L.; Keita, Å.V.; Parsons, B.N.; Prorok-Hamon, M.; Knight, P.; Winstanley, C.; Niamh, O.; Söderholm, J.D.; Rhodes, J.M.; Campbell, B.J. Soluble plantain fibre blocks adhesion and M-cell translocation of intestinal pathogens. J. Nutr. Biochem. 2013, 24, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, K.Z.; Santos, J.I.; Rosado, J.L.; Estrada-Garcia, T.; Haas, M.; Al Mamun, A.; DuPont, H.L.; Nanthakumar, N.N. Vitamin A supplementation modifies the association between mucosal innate and adaptive immune responses and resolution of enteric pathogen infections. Am. J. Clin. Nutr. 2011, 93, 578–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhaliwal, W.; Shawa, T.; Khanam, M.; Jagatiya, P.; Simuyandi, M.; Ndulo, N.; Bevins, C.L.; Sanderson, I.R.; Kelly, P. Intestinal antimicrobial gene expression: Impact of micronutrients in malnourished adults during a randomized trial. J. Infect. Dis. 2010, 202, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.C.; Garner, C.D.; Slauch, J.M.; Dwyer, Z.W.; Lawhon, S.D.; Frye, J.G.; McClelland, M.; Ahmer, B.M.; Altier, C. The intestinal fatty acid propionate inhibits Salmonella invasion through the post-translational control of HilD. Mol. Microbiol. 2013, 87, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Agerberth, B.; Bergman, P.; Gudmundsson, G.H. Helping the host: Induction of antimicrobial peptides as a novel therapeutic strategy against infections. In Antimicrobial Peptides and Innate Immunity; Springer: Basel, Switzerland, 2013; Volume 14, pp. 359–375. [Google Scholar]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, A.; Wang, X.; Gabello, M.; Valenzano, M.C.; Soler, A.P.; Ko, A.; Morin, P.J.; Mullin, J.M. Dietary methionine restriction improves colon tight junction barrier function and alters claudin expression pattern. Am. J. Physiol. Cell Physiol. 2010, 299, C1028–C1035. [Google Scholar] [CrossRef] [PubMed]
- Kau, A.L.; Ahern, P.P.; Griffin, N.W.; Goodman, A.L.; Gordon, J.I. Human nutrition, the gut microbiome and the immune system. Nature 2011, 474, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, S.J.; White, P.L.; Lathrop, S.L.; Solghan, S.M.; Medus, C.; McGlinchey, B.M.; Tobin-D’Angelo, M.; Marcus, R.; Mahon, B.E. Salmonella enterica Serotype Enteritidis: Increasing Incidence of Domestically Acquired Infections. Clin. Infect. Dis. 2012, 54, S488–S497. [Google Scholar] [CrossRef] [PubMed]
- Arguello, H.; Alvarez-Ordonez, A.; Carvajal, A.; Rubio, P.; Prieto, M. Role of Slaughtering in Salmonella Spreading and Control in Pork Production. J. Food Prot. 2013, 76, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, E.S.; Almond, G.W.; Routh, P.A.; Horton, J.R.; Dillman, R.C.; Orndorff, P.E. Experimental Salmonella typhi infection in the domestic pig, Sus scrofa domestica. Microb. Pathog. 2000, 29, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.L.; Zhang, S.; Tsolis, R.M.; Kingsley, R.A.; Adams, L.G.; Bäumler, A.J. Animal models of Salmonella infections: Enteritis versus typhoid fever. Microbes Infect. 2001, 3, 1335–1344. [Google Scholar] [CrossRef]
- Tsolis, R.M.; Kingsley, R.A.; Townsend, S.M.; Ficht, T.A.; Adams, L.G.; Bäumler, A.J. Of mice, calves, and men - Comparison of the mouse typhoid model with other Salmonella infections. Adv. Exp. Med. Biol. 1999, 473, 261–274. [Google Scholar] [PubMed]
- Oussalah, M.; Caillet, S.; Saucier, L.; Lacroix, M. Inhibitory effects of selected plant essential oils on the growth of four pathogenic bacteria: E. coli O157: H7, Salmonella Typhimurium, Staphylococcus aureus and Listeria monocytogenes. Food Control 2007, 18, 414–420. [Google Scholar] [CrossRef]
- Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.K.; Pillai, S.D.; Jayaraman, A.; Patil, B.S. Citrus flavonoid represses Salmonella pathogenicity island 1 and motility in S. typhimurium LT2. Int. J. Food Microbiol. 2011, 145, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ebani, V.V.; Nardoni, S.; Bertelloni, F.; Tosi, G.; Massi, P.; Pistelli, L.; Mancianti, F. In Vitro Antimicrobial Activity of Essential Oils against Salmonella enterica Serotypes Enteritidis and Typhimurium Strains Isolated from Poultry. Molecules 2019, 24, 900. [Google Scholar] [CrossRef]
- Chen, C.Y.; Tsen, H.Y.; Lin, C.L.; Lin, C.K.; Chuang, L.T.; Chen, C.S.; Chiang, Y.C. Enhancement of the immune response against Salmonella infection of mice by heat-killed multispecies combinations of lactic acid bacteria. J. Med. Microbiol. 2013, 62, 1657–1664. [Google Scholar] [CrossRef]
- Forbes, S.J.; Eschmann, M.; Mantis, N.J. Inhibition of Salmonella enterica serovar Typhimurium motility and entry into epithelial cells by a protective antilipopolysaccharide monoclonal immunoglobulin a antibody. Infect. Immun. 2008, 76, 4137–4144. [Google Scholar] [CrossRef]
- Hansen-Wester, I.; Hensel, M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001, 3, 549–559. [Google Scholar] [CrossRef]
- Bovee-Oudenhoven, I.M.J.; ten Bruggencate, S.J.M.; Lettink-Wissink, M.L.G.; van der Meer, R. Dietary fructo-oligosaccharides and lactulose inhibit intestinal colonisation but stimulate translocation of Salmonella in rats. Gut 2003, 52, 1572–1578. [Google Scholar] [CrossRef] [PubMed]
- Hitchins, A.D.; Wells, P.; McDonough, F.E.; Wong, N.P. Amelioration of the adverse effect of a gastrointestinal challenge with Salmonella enteritidis on weanling rats by a yogurt diet. Am. J. Clin. Nutr. 1985, 41, 92–100. [Google Scholar] [CrossRef]
- Kim, S.P.; Moon, E.; Nam, S.H.; Friedman, M. Composition of Herba Pogostemonis Water Extract and Protection of Infected Mice against Salmonella typhimurium-Induced Liver Damage and Mortality by Stimulation of Innate Immune Cells. J. Agric. Food Chem. 2012, 60, 12122–12130. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, P.O.; Aribisala, J.O.; Oladunmoye, M.K.; Arogunjo, A.O.; Ajayi-Moses, O.B. Therapeutic Effect of Goya Extra Virgin Olive Oil in Albino Rat Orogastricallly Dosed with Salmonella typhi. South Asian J. Res. Microbiol. 2019, 3, 1–9. [Google Scholar]
- Kelly, P.; Shawa, T.; Mwanamakondo, S.; Soko, R.; Smith, G.; Barclay, G.R.; Sanderson, I.R. Gastric and intestinal barrier impairment in tropical enteropathy and HIV: Limited impact of micronutrient supplementation during a randomised controlled trial. BMC Gastroenterol. 2010, 10, 72. [Google Scholar] [CrossRef]
- Tennant, S.M.; Hartland, E.L.; Phumoonna, T.; Lyras, D.; Rood, J.I.; Robins-Browne, R.M.; van Driel, I.R. Influence of gastric acid on susceptibility to infection with ingested bacterial pathogens. Infect. Immun. 2008, 76, 639–645. [Google Scholar] [CrossRef]
- Lucas, J.S.; Cochrane, S.A.; Warner, J.O.; Hourihane, J.O. The effect of digestion and pH on the allergenicity of kiwifruit proteins. Pediatr. Allergy Immunol.: Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2008, 19, 392–398. [Google Scholar] [CrossRef]
- Henderson, L.M.; Brewer, G.J.; Dressman, J.B.; Swidan, S.Z.; DuRoss, D.J.; Adair, C.H.; Barnett, J.L.; Berardi, R.R. Effect of intragastric pH on the absorption of oral zinc acetate and zinc oxide in young healthy volunteers. JPEN J. Parenter. Enter. Nutr. 1995, 19, 393–397. [Google Scholar] [CrossRef]
- Kararli, T.T. Comparison of the Gastrointestinal Anatomy, Physiology, and Biochemistry of Humans and Commonly Used Laboratory-Animals. Biopharm. Drug Dispos. 1995, 16, 351–380. [Google Scholar] [CrossRef]
- Guilloteau, P.; Zabielski, R.; Hammon, H.M.; Metges, C.C. Nutritional programming of gastrointestinal tract development. Is the pig a good model for man? Nutr. Res. Rev. 2010, 23, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Michiels, J.; Missotten, J.; Rasschaert, G.; Dierick, N.; Heyndrickx, M.; De Smet, S. Effect of organic acids on Salmonella colonization and shedding in weaned piglets in a seeder model. J. Food Prot. 2012, 75, 1974–1983. [Google Scholar] [CrossRef]
- Rajtak, U.; Boland, F.; Leonard, N.; Bolton, D.; Fanning, S. Roles of diet and the acid tolerance response in survival of common Salmonella serotypes in feces of finishing pigs. Appl. Environ. Microbiol. 2012, 78, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Boyen, F.; Haesebrouck, F.; Vanparys, A.; Volf, J.; Mahu, M.; Van Immerseel, F.; Rychlik, I.; Dewulf, J.; Ducatelle, R.; Pasmans, F. Coated fatty acids alter virulence properties of Salmonella typhimurium and decrease intestinal colonization of pigs. Vet. Microbiol. 2008, 132, 319–327. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, D.; Tian, G.; He, J.; Mao, X.; Mao, Q.; Yu, B. Dietary arginine supplementation alleviates immune challenge induced by Salmonella enterica serovar Choleraesuis bacterin potentially through the Toll-like receptor 4-myeloid differentiation factor 88 signalling pathway in weaned piglets. Br. J. Nutr. 2012, 108, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential oils as alternatives to antibiotics in swine production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Boyen, F.; Haesebrouck, F.; Maes, D.; Van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-typhoidal Salmonella infections in pigs: A closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 2008, 130, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.; Hennig, U.; Kuhla, S.; Brunner, R.M.; Kleessen, B.; Metges, C.C. Effect of inulin supplementation on selected gastric, duodenal, and caecal microbiota and short chain fatty acid pattern in growing piglets. Arch. Anim. Nutr. 2007, 61, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Scharek, L.; Tedin, K. The porcine immune system--differences compared to man and mouse and possible consequences for infections by Salmonella serovars. Berl. Und Munch. Tierarztl. Wochenschr. 2007, 120, 347–354. [Google Scholar]
- Higginson, E.E.; Simon, R.; Tennant, S.M. Animal models for salmonellosis: Applications in vaccine research. Clin. Vaccine Immunol. 2016, 23, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.F.; Paixao, T.A.; Tsolis, R.M.; Baumler, A.J.; Santos, R.L. Salmonellosis in cattle: Advantages of being an experimental model. Res. Vet. Sci. 2012, 93, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.M.; VandeHaar, M.J.; Sordillo, L.M.; Catherman, D.R.; Bateman Ii, H.G.; Schlotterbeck, R.L. Fatty acid intake alters growth and immunity in milk-fed calves. J. Dairy Sci. 2011, 94, 3936–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, R.R. Evolutionary Steps of Ecophysiological Adaptation and Diversification of Ruminants - a Comparative View of Their Digestive-System. Oecologia 1989, 78, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Li, R.W.; Connor, E.E.; Li, C.J.; Baldwin, R.L.; Sparks, M.E. Characterization of the rumen microbiota of pre-ruminant calves using metagenomic tools. Environ. Microbiol. 2012, 14, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, F.M.; Abu-El-Haija, M.A.; Gomez-Duarte, O.G. Salmonella infections: An update on epidemiology, management, and prevention. Travel Med. Infect. Dis. 2011, 9, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Connor, B.A.; Schwartz, E. Typhoid and paratyphoid fever in travellers. Lancet Infect. Dis 2005, 5, 623–628. [Google Scholar] [CrossRef]
- Hohmann, E.L. Nontyphoidal salmonellosis. Clin. Infect. Dis. 2001, 32, 263–269. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention. National Outbreak Reporting System (NORS), 2018. Available online: https://www.cdc.gov/nors/index.html (accessed on 1 January 2019).
- Glenn, L.M.; Lindsey, R.L.; Folster, J.P.; Pecic, G.; Boerlin, P.; Gilmour, M.W. Antimicrobial resistance genes in multidrug-resistant Salmonella enterica isolated from animals, retail meats, and humans in the United States and Canada. Microb. Drug Resist. 2013, 19, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; De Pasquale, I.; Ndagijimana, M.; Vernocchi, P.; Ricciuti, P.; Gagliardi, F.; Laghi, L.; Crecchio, C.; Guerzoni, M.E.; et al. Duodenal and faecal microbiota of celiac children: Molecular, phenotype and metabolome characterization. BMC Microbiol. 2011, 11, 219. [Google Scholar] [CrossRef]
- Lara-Villoslada, F.; Sierra, S.; Boza, J.; Xaus, J.; Olivares, M. Beneficial effects of consumption of a dairy product containing two probiotic strains, Lactobacillus coryniformis CECT5711 and Lactobacillus gasseri CECT5714 in healthy children. Nutr. Hosp. 2007, 22, 496–502. [Google Scholar]
- Rabbani, G.H.; Teka, T.; Zaman, B.; Majid, N.; Khatun, M.; Fuchs, G.J. Clinical studies in persistent diarrhea: Clinical studies in persistent diarrhea: Dietary management with green banana or pectin in Bangladeshi children. Gastroenterology 2001, 121, 554–560. [Google Scholar] [CrossRef]
- Kingamkono, R.; Sjogren, E.; Svanberg, U. Enteropathogenic bacteria in faecal swabs of young children fed on lactic acid-fermented cereal gruels. Epidemiol. Infect. 1999, 122, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Szabó, I.; Wieler, L.H.; Tedin, K.; Scharek-Tedin, L.; Taras, D.; Hensel, A.; Appel, B.; Nöckler, K. Influence of a probiotic strain of Enterococcus faecium on Salmonella enterica serovar Typhimurium DT104 infection in a porcine animal infection model. Appl. Environ. Microbiol. 2009, 75, 2621–2628. [Google Scholar]
- Pasetti, M.F.; Levine, M.M.; Sztein, M.B. Animal models paving the way for clinical trials of attenuated Salmonella enterica serovar Typhi live oral vaccines and live vectors. Vaccine 2003, 21, 401–418. [Google Scholar] [CrossRef]
- Hofmann, A.F.; Hagey, R.L. Bile acids: Chemistry, pathochemistry, biology, pathobiology, and therapeutics. Cell Mol. Life Sci. 2008, 65, 2461–2483. [Google Scholar] [CrossRef]
- Prieto, A.I.; Ramos-Morales, F.; Casadesus, J. Bile-induced DNA damage in Salmonella enterica. Genetics 2004, 168, 1787–1794. [Google Scholar] [CrossRef]
- Merritt, M.E.; Donaldson, J.R. Effect of bile salts on the DNA and membrane integrity of enteric bacteria. J. Med. Microbiol. 2009, 58, 1533–1541. [Google Scholar] [CrossRef] [Green Version]
- Begley, M.; Gahan, C.G.M.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Antunes, L.C.M.; Andersen, S.K.; Menendez, A.; Arena, E.T.; Han, J.; Ferreira, R.B.; Borchers, C.H.; Finlay, B.B. Metabolomics Reveals Phospholipids as Important Nutrient Sources during Salmonella Growth in Bile in Vitro and in Vivo. J. Bacteriol. 2011, 193, 4719–4725. [Google Scholar] [CrossRef]
- Kollanoor-Johny, A.; Mattson, T.; Baskaran, S.A.; Amalaradjou, M.A.R.; Hoagland, T.A.; Darre, M.J.; Khan, M.I.; Schreiber, D.T.; Donoghue, A.M.; Venkitanarayanan, K. Caprylic acid reduces Salmonella enteritidis populations in various segments of digestive tract and internal organs of 3- and 6-week-old broiler chickens, therapeutically. Poult. Sci. 2012, 91, 1686–1694. [Google Scholar] [CrossRef]
- Inagaki, T.; Moschetta, A.; Lee, Y.K.; Peng, L.; Zhao, G.; Downes, M.; Ruth, T.Y.; Shelton, J.M.; Richardson, J.A.; Repa, J.J.; et al. Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 3920–3925. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Xue, C.; Zhang, Y.; Liu, Y.; Wang, J.; Yu, X.; Zhang, X.; Zhang, R.; Yang, X.; Guo, C. Medium-Chain Fatty Acids Enhanced the Excretion of Fecal Cholesterol and Cholic Acid in C57BL/6J Mice Fed a Cholesterol-Rich Diet. Biosci. Biotechnol. Biochem. 2013, 77, 1390–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Hayashi, H.; Sugiyama, Y. Short- and medium-chain fatty acids enhance the cell surface expression and transport capacity of the bile salt export pump (BSEP/ABCB11). Biochim. Biophys. Acta 2010, 1801, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Costarelli, V.; Sanders, T.A. Acute effects of dietary fat composition on postprandial plasma bile acid and cholecystokinin concentrations in healthy premenopausal women. Br. J. Nutr. 2001, 86, 471–477. [Google Scholar] [CrossRef]
- Yang, Q.; Lan, T.; Chen, Y.; Dawson, P.A. Dietary fish oil increases fat absorption and fecal bile acid content without altering bile acid synthesis in 20-d-old weanling rats following massive ileocecal resection. Pediatr. Res. 2012, 72, 38–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prouty, A.M.; Brodsky, I.E.; Manos, J.; Belas, R.; Falkow, S.; Gunn, J.S. Transcriptional regulation of Salmonella enterica serovar typhimurium genes by bile. FEMS Immunol. Med. Microbiol. 2004, 41, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Prouty, A.M.; Gunn, J.S. Salmonella enterica serovar typhimurium invasion is repressed in the presence of bile. Infect. Immun. 2000, 68, 6763–6769. [Google Scholar] [CrossRef]
- Ye, W.; Li, Y.; Zhou, Z.; Wang, X.; Yao, J.; Liu, J.; Wang, C. Synthesis and antibacterial activity of new long-chain-alkyl bile acid-based amphiphiles. Bioorg. Chem. 2013, 51, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Wang, M.; Andersen, S.K.; Ferreira, R.B.; Kappelhoff, R.; Han, J.; Borchers, C.H.; Finlay, B.B. Repression of Salmonella enterica phoP expression by small molecules from physiological bile. J. Bacteriol. 2012, 194, 2286–2296. [Google Scholar] [CrossRef]
- Matsumoto, K.; Yokoyama, S.; Gato, N. Bile acid-binding activity of young persimmon (Diospyros kaki) fruit and its hypolipidemic effect in mice. Phytother. Res. 2010, 24, 205–210. [Google Scholar] [CrossRef]
- Andersson, K.E.; Immerstrand, T.; Swärd, K.; Bergenståhl, B.; Lindholm, M.W.; Öste, R.; Hellstrand, P. Effects of oats on plasma cholesterol and lipoproteins in C57BL/6 mice are substrain specific. Br. J. Nutr. 2010, 103, 513–521. [Google Scholar] [CrossRef]
- Chen, H.L.; Lin, Y.M.; Wang, Y.C. Comparative effects of cellulose and soluble fibers (pectin, konjac glucomannan, inulin) on fecal water toxicity toward Caco-2 cells, fecal bacteria enzymes, bile acid, and short-chain fatty acids. J. Agric. Food Chem. 2010, 58, 10277–10281. [Google Scholar] [CrossRef]
- Gunness, P.; Gidley, M.J. Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides. Food Funct. 2010, 1, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.F.; Eckmann, L. How bile acids confer gut mucosal protection against bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 4333–4334. [Google Scholar] [CrossRef] [Green Version]
- Zarepour, M.; Bhullar, K.; Montero, M.; Ma, C.; Huang, T.; Velcich, A.; Xia, L.; Vallance, B.A. The mucin Muc2 limits pathogen burdens and epithelial barrier dysfunction during Salmonella enterica serovar Typhimurium Colitis. Infect. Immun. 2013, 81, 3672–3683. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 2012, 15, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef]
- Kim, J.; Khan, W. Goblet Cells and Mucins: Role in Innate Defense in Enteric Infections. Pathogens 2013, 2, 55–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willemsen, L.E.; Koetsier, M.A.; van Deventer, S.J.; van Tol, E.A. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E (1) and E (2) production by intestinal myofibroblasts. Gut 2003, 52, 1442–1447. [Google Scholar] [CrossRef]
- Hino, S.; Takemura, N.; Sonoyama, K.; Morita, A.; Kawagishi, H.; Aoe, S.; Morita, T. Small intestinal goblet cell proliferation induced by ingestion of soluble and insoluble dietary fiber is characterized by an increase in sialylated mucins in rats. J. Nutr. 2012, 142, 1429–1436. [Google Scholar] [CrossRef]
- Kleessen, B.; Blaut, M. Modulation of gut mucosal biofilms. Br. J. Nutr. 2005, 93, S35–S40. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Tanabe, H.; Takahashi, K.; Sugiyama, K. Ingestion of resistant starch protects endotoxin influx from the intestinal tract and reduces D-galactosamine-induced liver injury in rats. J. Gastroenterol. Hepatol. 2004, 19, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Kosińska, A.; Andlauer, W. Modulation of Tight Junction Integrity by Food Components. Food Res. Int. 2013, 54, 951–960. [Google Scholar]
- Liu, J.H.; Xu, T.T.; Liu, Y.J.; Zhu, W.Y.; Mao, S.Y. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Psychol. Regul. Integr. Comp. Physiol. 2013, 305, R232–R241. [Google Scholar] [CrossRef] [PubMed]
- Nofrarias, M.; Martinez-Puig, D.; Pujols, J.; Majo, N.; Perez, J.F. Long-term intake of resistant starch improves colonic mucosal integrity and reduces gut apoptosis and blood immune cells. Nutrition 2007, 23, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Thomson, A.B.; Clandinin, M.T. Protection of intestinal occludin tight junction protein by dietary gangliosides in lipopolysaccharide-induced acute inflammation. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 321–328. [Google Scholar] [CrossRef]
- Suzuki, T.; Hara, H. Quercetin Enhances Intestinal Barrier Function through the Assembly of Zonnula Occludens-2, Occludin, and Claudin-1 and the Expression of Claudin-4 in Caco-2 Cells. J. Nutr. 2009, 139, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Devaraj, S.N. Protective role of Hemidesmus indicus R. Br. root extract against Salmonella typhimurium-induced cytotoxicity in Int 407 cell line. Phytother. Res. 2007, 21, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Willing, B.P.; Vacharaksa, A.; Croxen, M.; Thanachayanont, T.; Finlay, B.B. Altering Host Resistance to Infections through Microbial Transplantation. PLoS ONE 2011, 6, e26988. [Google Scholar] [CrossRef] [PubMed]
- Croswell, A.; Amir, E.; Teggatz, P.; Barman, M.; Salzman, N.H. Prolonged impact of antibiotics on intestinal microbial ecology and susceptibility to enteric Salmonella infection. Infect. Immun. 2009, 77, 2741–2753. [Google Scholar] [CrossRef]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Nunez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef]
- Stelter, C.; Käppeli, R.; König, C.; Krah, A.; Hardt, W.D.; Stecher, B.; Bumann, D. Salmonella-induced mucosal lectin RegIIIbeta kills competing gut microbiota. PLoS ONE 2011, 6, e20749. [Google Scholar] [CrossRef]
- Gill, N.; Ferreira, R.B.; Antunes, L.C.M.; Willing, B.P.; Sekirov, I.; Al-Zahrani, F.; Hartmann, M.; Finlay, B.B. Neutrophil elastase alters the murine gut microbiota resulting in enhanced Salmonella colonization. PLoS ONE 2012, 7, e49646. [Google Scholar] [CrossRef]
- Castillo, N.A.; de Moreno de LeBlanc, A.; Galdeano, C.M.; Perdigón, G. Probiotics: An alternative strategy for combating salmonellosis: Immune mechanisms involved. Food Res. Int. 2012, 45, 831–841. [Google Scholar] [CrossRef]
- Burkholder, K.M.; Bhunia, A.K. Salmonella enterica serovar typhimurium adhesion and cytotoxicity during epithelial cell stress is reduced by Lactobacillus rhamnosus GG. Gut Pathog. 2009, 1, 14. [Google Scholar] [CrossRef]
- Castillo, N.A.; Perdigon, G.; de Moreno de LeBlanc, A. Oral administration of a probiotic Lactobacillus modulates cytokine production and TLR expression improving the immune response against Salmonella enterica serovar typhimurium infection in mice. BMC Microbiol. 2011, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Bayoumi, M.A.; Griffiths, M.W. Probiotics down-regulate genes in Salmonella enterica serovar typhimurium pathogenicity islands 1 and 2. J. Food Prot. 2010, 73, 452–460. [Google Scholar] [CrossRef]
- McNulty, N.P.; Wu, M.; Erickson, A.R.; Pan, C.; Erickson, B.; Martens, E.C.; Pudlo, N.A.; Muegge, B.D.; Henrissat, B.; Hettich, R.L.; et al. Effects of Diet on Resource Utilization by a Model Human Gut Microbiota Containing Bacteroides cellulosilyticus WH2, a Symbiont with an Extensive Glycobiome. PLoS Biol. 2013, 11, e1001637. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci. Transl. Med. 2009, 1, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Toward, R.; Montandon, S.; Walton, G.; Gibson, G.R. Effect of prebiotics on the human gut microbiota of elderly persons. Gut Microbes 2012, 3, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res.: Off. J. Ital. Pharmacol. Soc. 2010, 61, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Walker, W.A. Innate immunity of the gut: Mucosal defense in health and disease. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Pattern recognition receptors: Doubling up for the innate immune response. Cell 2002, 111, 927–930. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Mccoy, K.D.; Johansen, F.E.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Fagarasan, S.; Honjo, T. Intestinal IgA synthesis: Regulation of front-line body defences. Nat. Rev. Immunol. 2003, 3, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Nanton, M.R.; Way, S.S.; Shlomchik, M.J.; McSorley, S.J. Cutting edge: B cells are essential for protective immunity against Salmonella independent of antibody secretion. J. Immunol. 2012, 189, 5503–5507. [Google Scholar] [CrossRef] [PubMed]
- Cross, M.L. Microbes versus microbes: Immune signals generated by probiotic lactobacilli and their role in protection against microbial pathogens. Fems Immunol. Med. Microbiol. 2002, 34, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Jouanguy, E.; DÖffinger, R.; Dupuis, S.; Pallier, A.; Altare, F.; Casanova, J.L. IL-12 and IFN-gamma in host defense against mycobacteria and salmonella in mice and men. Curr. Opin. Immunol. 1999, 11, 346–351. [Google Scholar] [CrossRef]
- Bao, S.; Beagley, K.W.; France, M.P.; Shen, J.; Husband, A.J. Interferon-gamma plays a critical role in intestinal immunity against Salmonella typhimurium infection. Immunology 2000, 99, 464–472. [Google Scholar] [CrossRef]
- Arnold, J.W.; Niesel, D.W.; Annable, C.R.; Hess, C.B.; Asuncion, M.; Cho, Y.J.; Peterson, J.W.; Klimpel, G.R. Tumor necrosis factor-alpha mediates the early pathology in Salmonella infection of the gastrointestinal tract. Microb. Pathog. 1993, 14, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Kirby, A.C.; Yrlid, U.; Wick, M.J. The innate immune response differs in primary and secondary Salmonella infection. J. Immunol. 2002, 169, 4450–4459. [Google Scholar] [CrossRef] [PubMed]
- Mastroeni, P.; Harrison, J.A.; Chabalgoity, J.A.; Hormaeche, C.E. Effect of interleukin 12 neutralization on host resistance and gamma interferon production in mouse typhoid. Infect. Immun. 1996, 64, 189–196. [Google Scholar] [PubMed]
- Lapaque, N.; Walzer, T.; Meresse, S.; Vivier, E.; Trowsdale, J. Interactions between Human NK Cells and Macrophages in Response to Salmonella Infection. J. Immunol. 2009, 182, 4339–4348. [Google Scholar] [CrossRef]
- Mittrucker, H.W.; Kaufmann, S.H.E. Immune response to infection with Salmonella typhimurium in mice. J. Leukoc. Biol. 2000, 67, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Michetti, P.; Mahan, M.J.; Slauch, J.M.; Mekalanos, J.J.; Neutra, M.R. Monoclonal Secretory Immunoglobulin-a Protects Mice against Oral Challenge with the Invasive Pathogen Salmonella-typhimurium. Infect. Immun. 1992, 60, 1786–1792. [Google Scholar] [PubMed]
- Schley, P.D.; Field, C.J. The immune-enhancing effects of dietary fibres and prebiotics. Br. J. Nutr. 2002, 87, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Núñez, I.N.; de Moreno de LeBlanc, A.; Carmuega, E.; Weill, R. Perdigón Impact of a probiotic fermented milk in the gut ecosystem and in the systemic immunity using a non-severe protein-energy-malnutrition model in mice. BMC Gastroenterol. 2011, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Guerrant, R.L.; Oria, R.B.; Moore, S.R.; Oria, M.O.B.; Lima, A.A.M. Malnutrition as an enteric infectious disease with long-term effects on child development. Nutr. Rev. 2008, 66, 487–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoneum, M.; Matsuura, M. Augmentation of macrophage phagocytosis by modified arabinoxylan rice bran (MGN-3/biobran). Int. J. Immunopathol. Pharm. 2004, 17, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Niu, X.; Du, X.; Smith, D.; Meydani, S.N.; Wu, D. Dietary supplementation with white button mushrooms augments the protective immune response to salmonella vaccine in mice. J. Nutr. 2014, 144, 98–105. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relationships between dietary bioactive components and Salmonella infection. Dietary bioactive components such as fiber, amino acids, vitamins and minerals, fatty acids, and polyphenols improve the gut epithelium, microbiota, and immunity that may eventually lead to increased resistance to Salmonella infection.
Figure 1.
Relationships between dietary bioactive components and Salmonella infection. Dietary bioactive components such as fiber, amino acids, vitamins and minerals, fatty acids, and polyphenols improve the gut epithelium, microbiota, and immunity that may eventually lead to increased resistance to Salmonella infection.


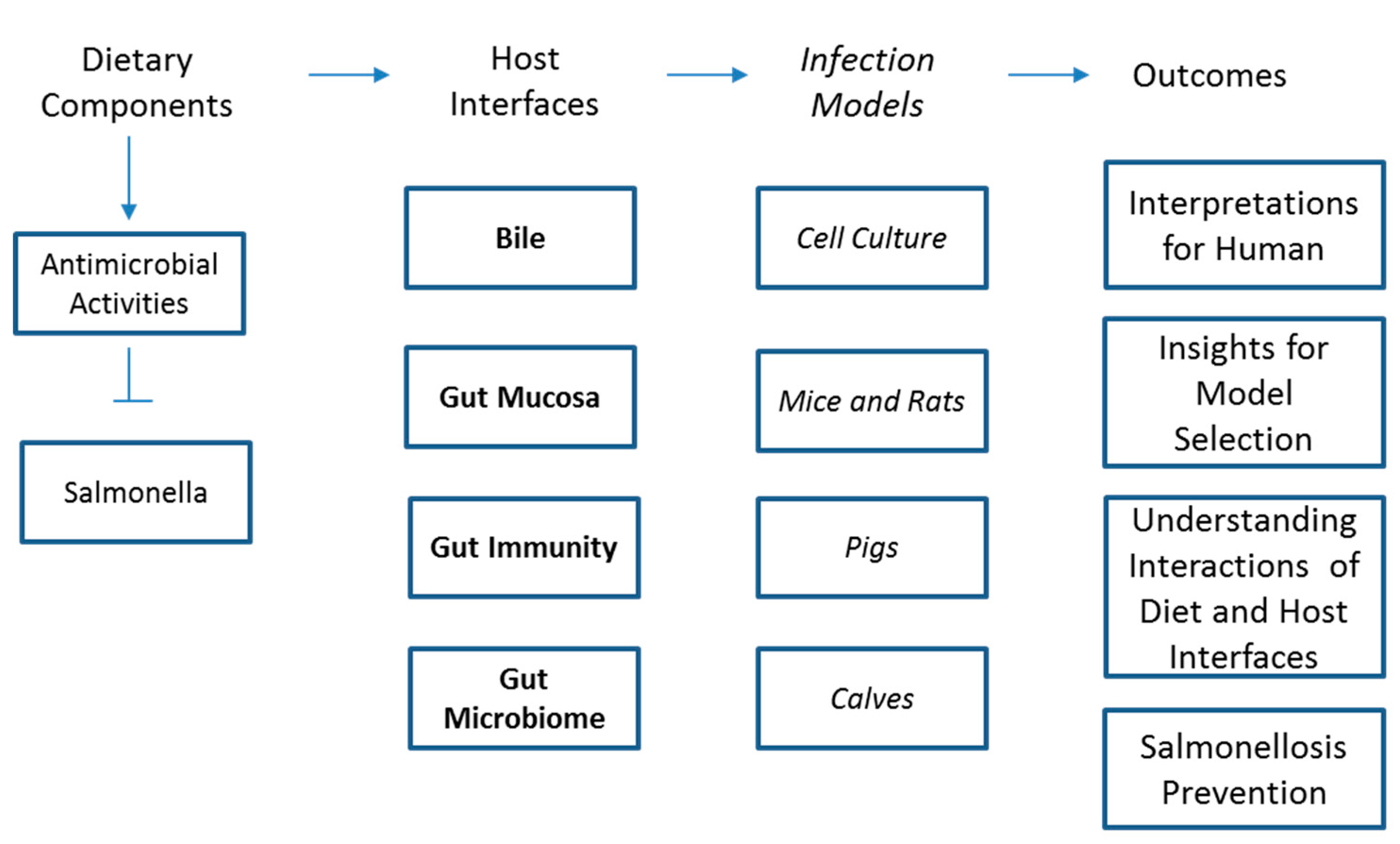{kind=link}
Table 1.
Strengths and weaknesses of Salmonella models discussed in the current study.
| Infection Model | Strength | Weaknesses |
|---|---|---|
| Salmonella Culture (Non-Host) | Direct interaction with the pathogen without confounders | Does not represent the interaction of dietary components with the host |
| Co-culture of Salmonella with host cell | Increased complexity of interaction compared to only pathogen culture, represents effects of intervention on the pathogen as well as the host | Does not represent involvement of all the host cell types that simultaneously happen together in human |
| Rodent Models | Represent a complex living system, very economical and convenient, ease in genetic manipulation to know mechanistic pathways | No diarrhea and vomiting, different intestinal immunity, different gastric environment, and anatomical structures |
| Pig Models | Similar to humans in body composition, cardiovascular, renal, nutritional, immunological, metabolic, and gastrointestinal aspects | Different than humans in Salmonella colonization pattern, gastric acidity, bile quantities, mucus thickness, immune system, not economical, not convenient |
| Calf Models | Develop similar clinical and pathological features such as diarrhea and enteritis | Stomach structure is different, not economical |
| Clinical Trials | The ideal model | Difficult to study the preventive effects of interventions due to ethical considerations |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kumar, A.; Allison, A.; Henry, M.; Scales, A.; Fouladkhah, A.C. Development of Salmonellosis as Affected by Bioactive Food Compounds. Microorganisms 2019, 7, 364. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090364
AMA Style
Kumar A, Allison A, Henry M, Scales A, Fouladkhah AC. Development of Salmonellosis as Affected by Bioactive Food Compounds. Microorganisms. 2019; 7(9):364. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090364
Chicago/Turabian StyleKumar, Ajay, Abimbola Allison, Monica Henry, Anita Scales, and Aliyar Cyrus Fouladkhah. 2019. "Development of Salmonellosis as Affected by Bioactive Food Compounds" Microorganisms 7, no. 9: 364. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090364
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.